
Small Molecules for the Treatment of Long-COVID-Related Vascular Damage and Abnormal Blood Clotting: A Patent-Based Appraisal

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Vascular Disease and Blood Clotting in Long COVID
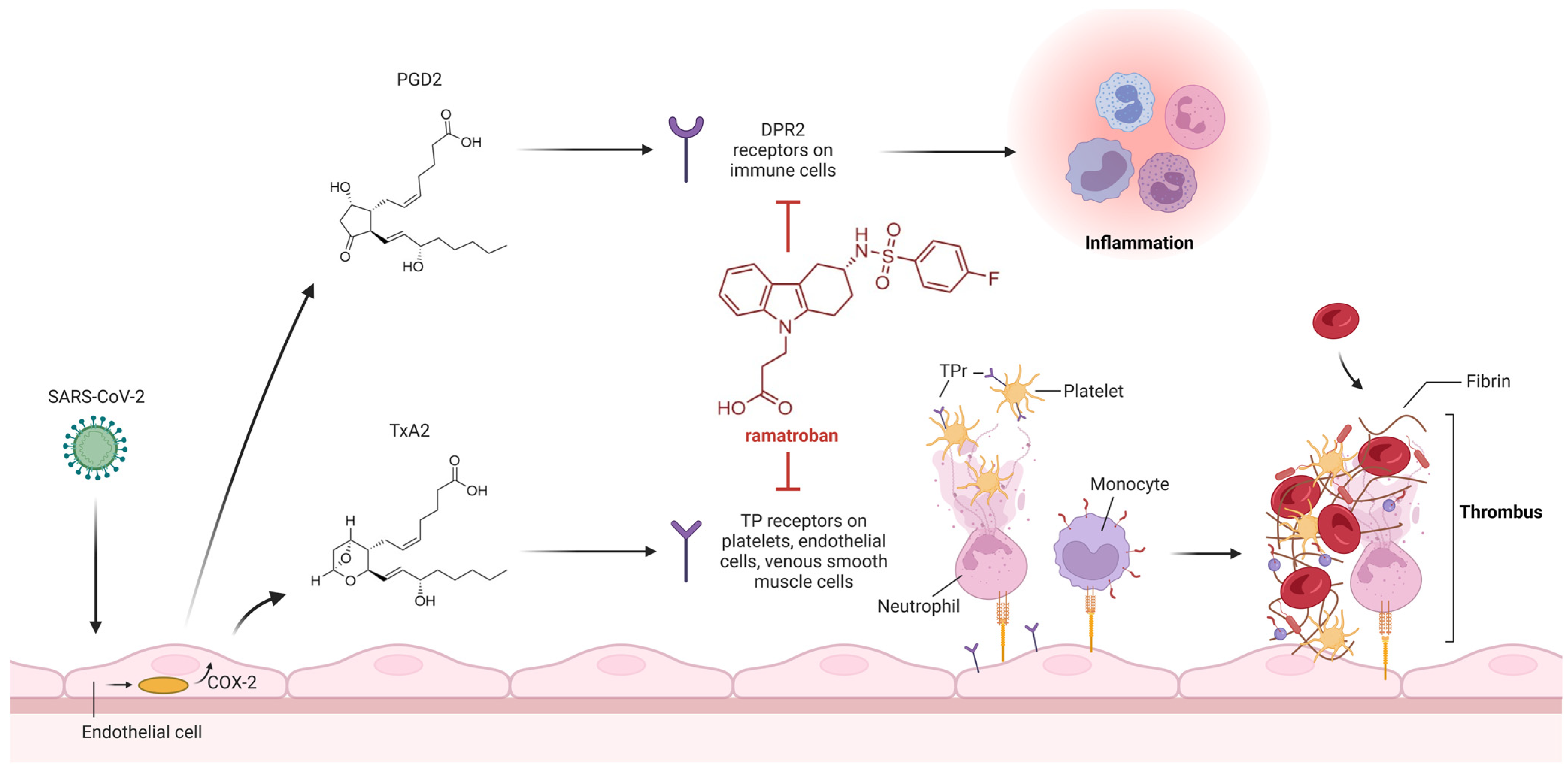
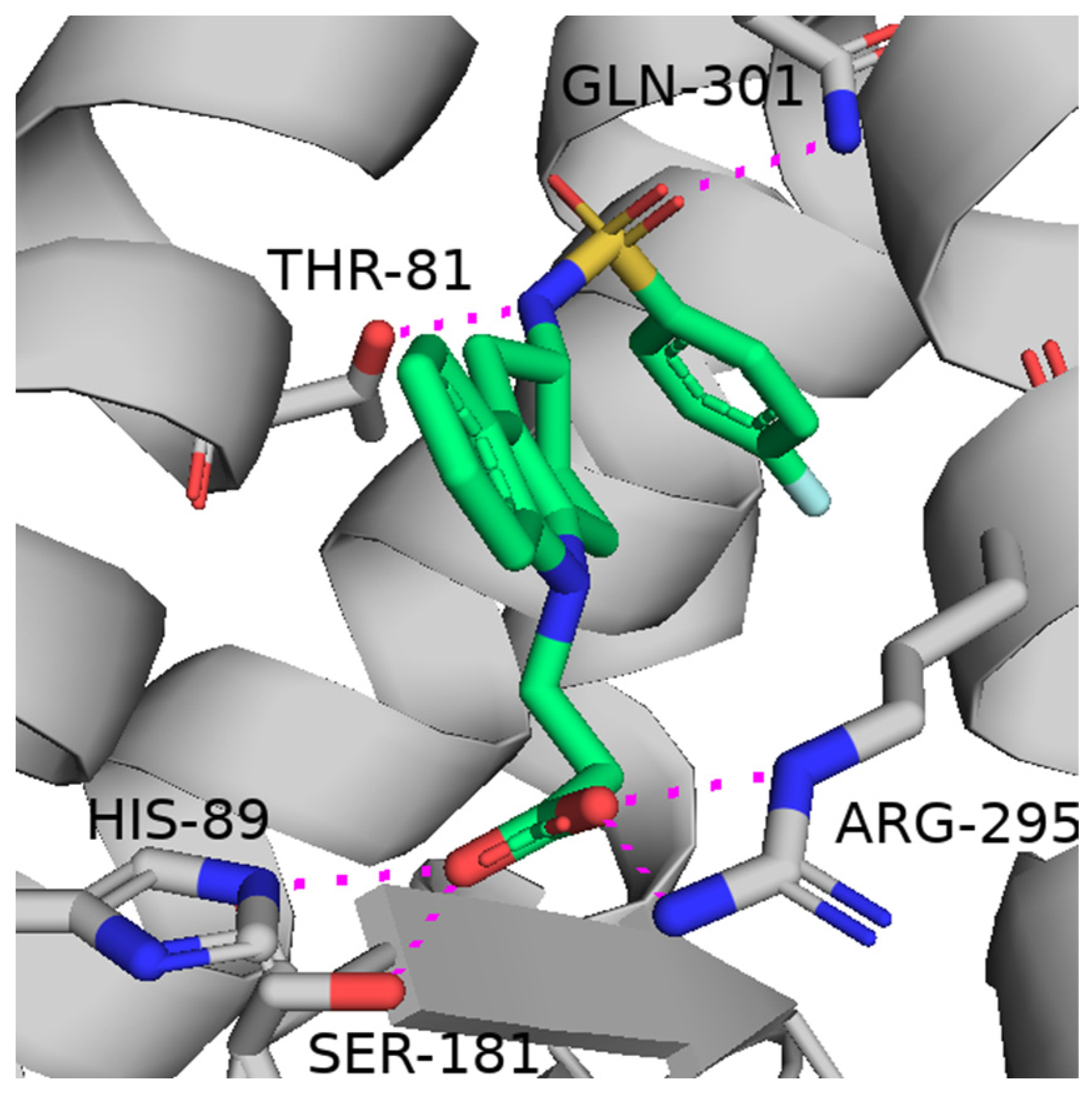
3. Thromboxane A2 Receptor Antagonists
4. Caspase Inhibitors
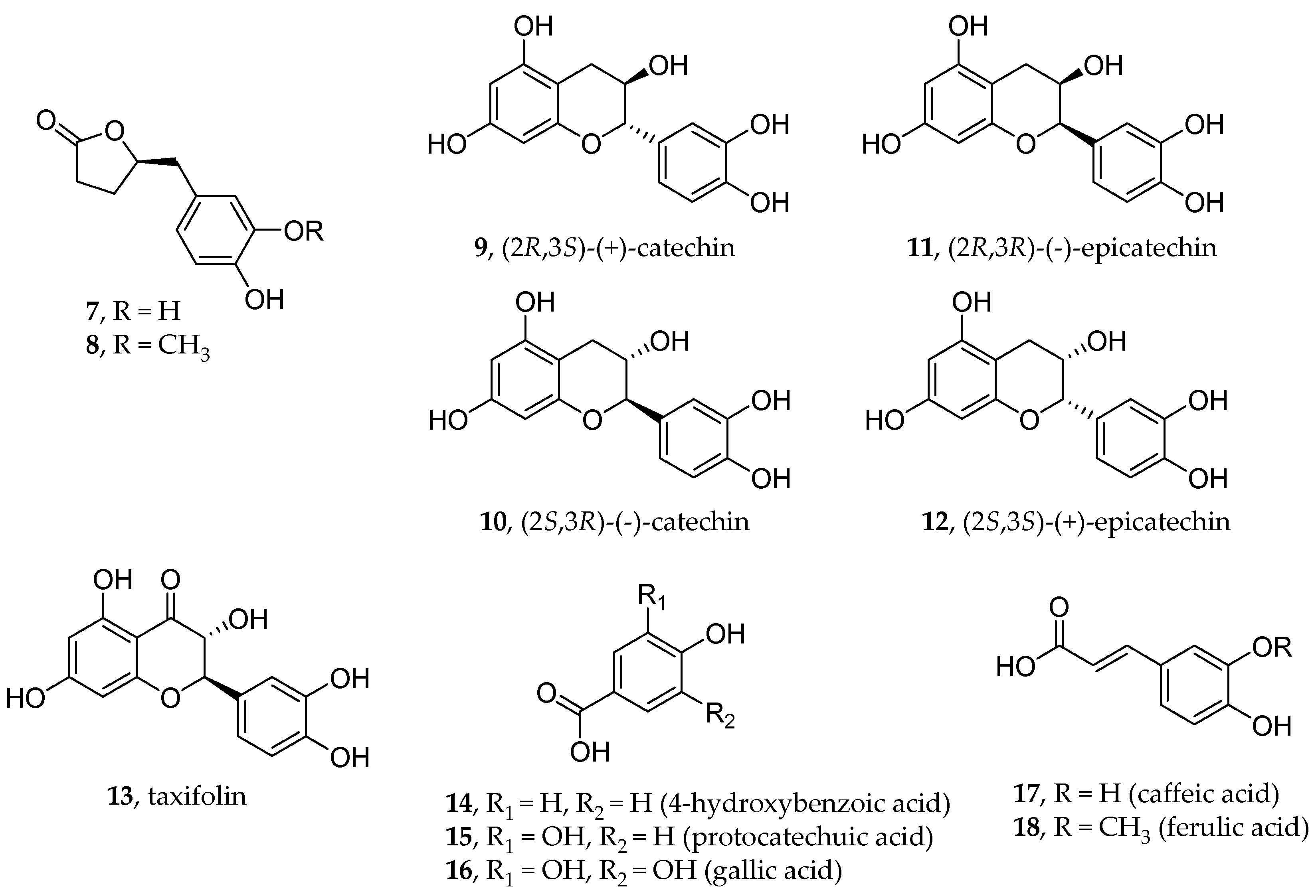
5. Polyphenols
6. NHE-1 Inhibitors
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.W.; et al. The Species Severe Acute Respiratory Syndrome-Related Coronavirus: Classifying 2019-nCoV and Naming It SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef]
- Hu, B.; Huang, S.; Yin, L. The Cytokine Storm and COVID-19. J. Med. Virol. 2021, 93, 250–256. [Google Scholar] [CrossRef]
- Nagoor Meeran, M.F.; Sharma, C.; Goyal, S.N.; Kumar, S.; Ojha, S. CB2 Receptor-Selective Agonists as Candidates for Targeting Infection, Inflammation, and Immunity in SARS-CoV-2 Infections. Drug Dev. Res. 2021, 82, 7–11. [Google Scholar] [CrossRef]
- Zhang, W.; Qin, C.; Fei, Y.; Shen, M.; Zhou, Y.; Zhang, Y.; Zeng, X.; Zhang, S. Anti-Inflammatory and Immune Therapy in Severe Coronavirus Disease 2019 (COVID-19) Patients: An Update. Clin. Immunol. 2022, 239, 109022. [Google Scholar] [CrossRef]
- Graziano, G.; Delre, P.; Carofiglio, F.; Brea, J.; Ligresti, A.; Kostrzewa, M.; Riganti, C.; Gioè-Gallo, C.; Majellaro, M.; Nicolotti, O.; et al. N-Adamantyl-Anthranil Amide Derivatives: New Selective Ligands for the Cannabinoid Receptor Subtype 2 (CB2R). Eur. J. Med. Chem. 2023, 248, 115109. [Google Scholar] [CrossRef] [PubMed]
- Gasperi, V.; Guzzo, T.; Topai, A.; Gambacorta, N.; Ciriaco, F.; Nicolotti, O.; Maccarrone, M. Recent Advances on Type-2 Cannabinoid (CB2) Receptor Agonists and Their Therapeutic Potential. Curr. Med. Chem. 2023, 30, 1420–1457. [Google Scholar] [CrossRef] [PubMed]
- Gambacorta, N.; Ciriaco, F.; Amoroso, N.; Altomare, C.D.; Bajorath, J.; Nicolotti, O. CIRCE: Web-Based Platform for the Prediction of Cannabinoid Receptor Ligands Using Explainable Machine Learning. J. Chem. Inf. Model. 2023, 63, 5916–5926. [Google Scholar] [CrossRef] [PubMed]
- Gambacorta, N.; Gasperi, V.; Guzzo, T.; Di Leva, F.S.; Ciriaco, F.; Sánchez, C.; Tullio, V.; Rozzi, D.; Marinelli, L.; Topai, A.; et al. Exploring the 1,3-Benzoxazine Chemotype for Cannabinoid Receptor 2 as a Promising Anti-Cancer Therapeutic. Eur. J. Med. Chem. 2023, 259, 115647. [Google Scholar] [CrossRef] [PubMed]
- George, P.M.; Reed, A.; Desai, S.R.; Devaraj, A.; Faiez, T.S.; Laverty, S.; Kanwal, A.; Esneau, C.; Liu, M.K.C.; Kamal, F.; et al. A Persistent Neutrophil-Associated Immune Signature Characterizes Post–COVID-19 Pulmonary Sequelae. Sci. Transl. Med. 2022, 14, eabo5795. [Google Scholar] [CrossRef] [PubMed]
- Fernández-de-las-Peñas, C.; Palacios-Ceña, D.; Gómez-Mayordomo, V.; Cuadrado, M.L.; Florencio, L.L. Defining Post-COVID Symptoms (Post-Acute COVID, Long COVID, Persistent Post-COVID): An Integrative Classification. Int. J. Environ. Res. Public Health 2021, 18, 2621. [Google Scholar] [CrossRef]
- Gambacorta, N.; Caputo, L.; Quintieri, L.; Monaci, L.; Ciriaco, F.; Nicolotti, O. Rational Discovery of Antiviral Whey Protein-Derived Small Peptides Targeting the SARS-CoV-2 Main Protease. Biomedicines 2022, 10, 1067. [Google Scholar] [CrossRef] [PubMed]
- Stefanelli, I.; Corona, A.; Cerchia, C.; Cassese, E.; Improta, S.; Costanzi, E.; Pelliccia, S.; Morasso, S.; Esposito, F.; Paulis, A.; et al. Broad-Spectrum Coronavirus 3C-like Protease Peptidomimetic Inhibitors Effectively Block SARS-CoV-2 Replication in Cells: Design, Synthesis, Biological Evaluation, and X-Ray Structure Determination. Eur. J. Med. Chem. 2023, 253, 115311. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.; Wood, J.; Jaycox, J.R.; Dhodapkar, R.M.; Lu, P.; Gehlhausen, J.R.; Tabachnikova, A.; Greene, K.; Tabacof, L.; Malik, A.A.; et al. Distinguishing Features of Long COVID Identified through Immune Profiling. Nature 2023, 623, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Ahamed, J.; Laurence, J. Long COVID Endotheliopathy: Hypothesized Mechanisms and Potential Therapeutic Approaches. J. Clin. Investig. 2022, 132, e161167. [Google Scholar] [CrossRef] [PubMed]
- Raveendran, A.V.; Jayadevan, R.; Sashidharan, S. Long COVID: An Overview. Diabetes Metab. Syndr. 2021, 15, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Proal, A.D.; VanElzakker, M.B. Long COVID or Post-Acute Sequelae of COVID-19 (PASC): An Overview of Biological Factors That May Contribute to Persistent Symptoms. Front. Microbiol. 2021, 12, 698169. [Google Scholar] [CrossRef]
- Riva, G.; Nasillo, V.; Tagliafico, E.; Trenti, T.; Comoli, P.; Luppi, M. COVID-19: More than a Cytokine Storm. Crit. Care 2020, 24, 549. [Google Scholar] [CrossRef]
- Iba, T.; Levy, J.H.; Levi, M.; Thachil, J. Coagulopathy in COVID-19. J. Thromb. Haemost. 2020, 18, 2103–2109. [Google Scholar] [CrossRef]
- Iba, T.; Wada, H.; Levy, J.H. Platelet Activation and Thrombosis in COVID-19. Semin. Thromb. Hemost. 2023, 49, 55–61. [Google Scholar] [CrossRef]
- Roncati, L.; Ligabue, G.; Nasillo, V.; Lusenti, B.; Gennari, W.; Fabbiani, L.; Malagoli, C.; Gallo, G.; Giovanella, S.; Lupi, M.; et al. A Proof of Evidence Supporting Abnormal Immunothrombosis in Severe COVID-19: Naked Megakaryocyte Nuclei Increase in the Bone Marrow and Lungs of Critically Ill Patients. Platelets 2020, 31, 1085–1089. [Google Scholar] [CrossRef]
- Elhadad, S.; Redmond, D.; Tan, A.; Huang, J.; Rodriguez, B.L.; Racine-Brzostek, S.E.; Subrahmanian, S.; Ahamed, J.; Laurence, J. Defibrotide Mitigates Endothelial Cell Injury Induced by Plasmas from Patients with COVID-19 and Related Vasculopathies. Thromb. Res. 2023, 225, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.C.; Devason, A.S.; Umana, I.C.; Cox, T.O.; Dohnalová, L.; Litichevskiy, L.; Perla, J.; Lundgren, P.; Etwebi, Z.; Izzo, L.T.; et al. Serotonin Reduction in Post-Acute Sequelae of Viral Infection. Cell 2023, 186, 4851–4867.e20. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, H.K.; Libby, P.; Ridker, P.M. COVID-19—A Vascular Disease. Trends Cardiovasc. Med. 2021, 31, 1–5. [Google Scholar] [CrossRef]
- Espacenet. Available online: https://worldwide.espacenet.com/ (accessed on 1 December 2023).
- Clinical Trials. Available online: https://clinicaltrials.gov/ (accessed on 1 December 2023).
- Xie, Y.; Xu, E.; Bowe, B.; Al-Aly, Z. Long-Term Cardiovascular Outcomes of COVID-19. Nat. Med. 2022, 28, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Mehra, M.R.; Desai, S.S.; Kuy, S.; Henry, T.D.; Patel, A.N. Cardiovascular Disease, Drug Therapy, and Mortality in COVID-19. N. Engl. J. Med. 2020, 382, e102. [Google Scholar] [CrossRef]
- Cervia-Hasler, C.; Brüningk, S.C.; Hoch, T.; Fan, B.; Muzio, G.; Thompson, R.C.; Ceglarek, L.; Meledin, R.; Westermann, P.; Emmenegger, M.; et al. Persistent Complement Dysregulation with Signs of Thromboinflammation in Active Long Covid. Science 2024, 383, eadg7942. [Google Scholar] [CrossRef] [PubMed]
- Roncati, L.; Ligabue, G.; Fabbiani, L.; Malagoli, C.; Gallo, G.; Lusenti, B.; Nasillo, V.; Manenti, A.; Maiorana, A. Type 3 Hypersensitivity in COVID-19 Vasculitis. Clin. Immunol. 2020, 217, 108487. [Google Scholar] [CrossRef]
- Becker, R.C. COVID-19-Associated Vasculitis and Vasculopathy. J. Thromb. Thrombolysis 2020, 50, 499–511. [Google Scholar] [CrossRef]
- Stark, K.; Massberg, S. Interplay between Inflammation and Thrombosis in Cardiovascular Pathology. Nat. Rev. Cardiol. 2021, 18, 666–682. [Google Scholar] [CrossRef]
- Theofilis, P.; Sagris, M.; Oikonomou, E.; Antonopoulos, A.S.; Siasos, G.; Tsioufis, C.; Tousoulis, D. Inflammatory Mechanisms Contributing to Endothelial Dysfunction. Biomedicines 2021, 9, 781. [Google Scholar] [CrossRef]
- Purgatorio, R.; Gambacorta, N.; de Candia, M.; Catto, M.; Rullo, M.; Pisani, L.; Nicolotti, O.; Altomare, C.D. First-in-Class Isonipecotamide-Based Thrombin and Cholinesterase Dual Inhibitors with Potential for Alzheimer Disease. Molecules 2021, 26, 5208. [Google Scholar] [CrossRef]
- ATTACC, ACTIV-4a, and REMAP-CAP Investigators. Therapeutic Anticoagulation with Heparin in Noncritically Ill Patients with COVID-19. N. Engl. J. Med. 2021, 385, 790–802. [Google Scholar] [CrossRef]
- Hempel, T.; Elez, K.; Krüger, N.; Raich, L.; Shrimp, J.H.; Danov, O.; Jonigk, D.; Braun, A.; Shen, M.; Hall, M.D.; et al. Synergistic Inhibition of SARS-CoV-2 Cell Entry by Otamixaban and Covalent Protease Inhibitors: Pre-Clinical Assessment of Pharmacological and Molecular Properties. Chem. Sci. 2021, 12, 12600–12609. [Google Scholar] [CrossRef] [PubMed]
- De Maio, F.; Rullo, M.; de Candia, M.; Purgatorio, R.; Lopopolo, G.; Santarelli, G.; Palmieri, V.; Papi, M.; Elia, G.; De Candia, E.; et al. Evaluation of Novel Guanidino-Containing Isonipecotamide Inhibitors of Blood Coagulation Factors against SARS-CoV-2 Virus Infection. Viruses 2022, 14, 1730. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- Trifirò, G.; Massari, M.; Da Cas, R.; Menniti Ippolito, F.; Sultana, J.; Crisafulli, S.; Giorgi Rossi, P.; Marino, M.; Zorzi, M.; Bovo, E.; et al. Renin-Angiotensin-Aldosterone System Inhibitors and Risk of Death in Patients Hospitalised with COVID-19: A Retrospective Italian Cohort Study of 43,000 Patients. Drug Saf. 2020, 43, 1297–1308. [Google Scholar] [CrossRef]
- Huggins, D.J. Structural Analysis of Experimental Drugs Binding to the SARS-CoV-2 Target TMPRSS2. J. Mol. Graph. Model. 2020, 100, 107710. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.A.; Goodman, Z.; Jabbar, A.; Vemulapalli, R.; Younes, Z.H.; Freilich, B.; Sheikh, M.Y.; Schattenberg, J.M.; Kayali, Z.; Zivony, A.; et al. A Randomized, Placebo-Controlled Trial of Emricasan in Patients with NASH and F1-F3 Fibrosis. J. Hepatol. 2020, 72, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Fraser, B.J.; Beldar, S.; Seitova, A.; Hutchinson, A.; Mannar, D.; Li, Y.; Kwon, D.; Tan, R.; Wilson, R.P.; Leopold, K.; et al. Structure and Activity of Human TMPRSS2 Protease Implicated in SARS-CoV-2 Activation. Nat. Chem. Biol. 2022, 18, 963–971. [Google Scholar] [CrossRef]
- Guertin, K.R.; Gardner, C.J.; Klein, S.I.; Zulli, A.L.; Czekaj, M.; Gong, Y.; Spada, A.P.; Cheney, D.L.; Maignan, S.; Guilloteau, J.-P.; et al. Optimization of the β-Aminoester Class of Factor Xa Inhibitors. Part 2: Identification of FXV673 as a Potent and Selective Inhibitor with Excellent In Vivo Anticoagulant Activity. Bioorganic Med. Chem. Lett. 2002, 12, 1671–1674. [Google Scholar] [CrossRef]
- Peiffer, A.L.; Garlick, J.M.; Wu, Y.; Wotring, J.W.; Arora, S.; Harmata, A.S.; Bochar, D.A.; Stephenson, C.J.; Soellner, M.B.; Sexton, J.Z.; et al. TMPRSS2 Inhibitor Discovery Facilitated through an In Silico and Biochemical Screening Platform. ACS Med. Chem. Lett. 2023, 14, 860–866. [Google Scholar] [CrossRef]
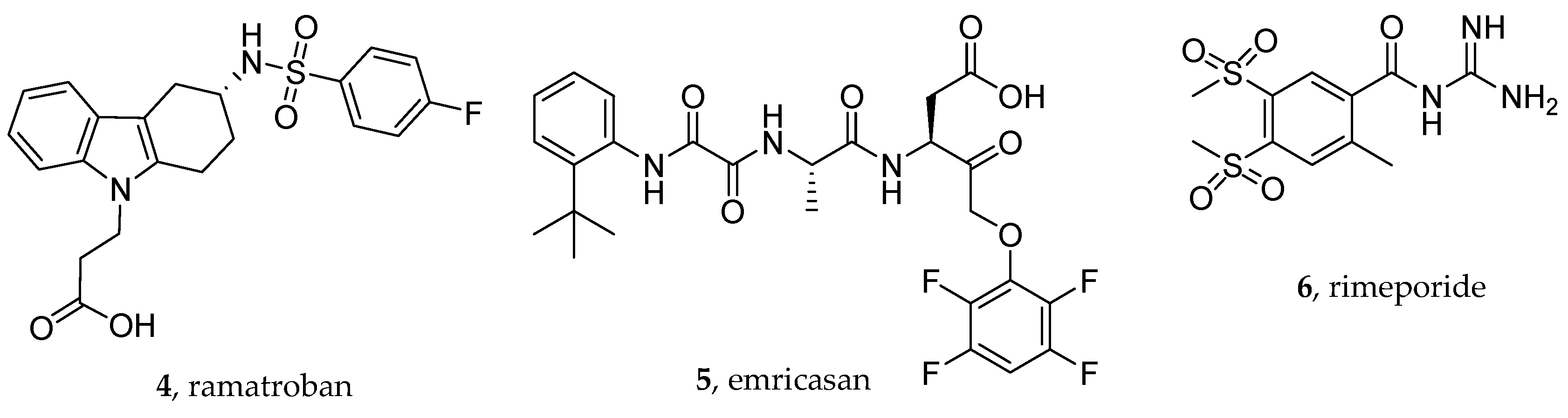
- Chiang, K.C.; Rizk, J.G.; Nelson, D.J.; Krishnamurti, L.; Subbian, S.; Imig, J.D.; Khan, I.; Reddy, S.T.; Gupta, A. Ramatroban for Chemoprophylaxis and Treatment of COVID-19: David Takes on Goliath. Expert Opin. Ther. Targets 2022, 26, 13–28. [Google Scholar] [CrossRef]
- Gupta, A.; Kalantar-Zadeh, K.; Reddy, S.T. Ramatroban as a Novel Immunotherapy for COVID-19. J. Mol. Genet. Med. 2020, 14. [Google Scholar] [CrossRef]
- Maroli, N.; Bhasuran, B.; Natarajan, J.; Kolandaivel, P. The Potential Role of Procyanidin as a Therapeutic Agent against SARS-CoV-2: A Text Mining, Molecular Docking and Molecular Dynamics Simulation Approach. J. Biomol. Struct. Dyn. 2022, 40, 1230–1245. [Google Scholar] [CrossRef]
- Bardelčíková, A.; Miroššay, A.; Šoltýs, J.; Mojžiš, J. Therapeutic and Prophylactic Effect of Flavonoids in Post-COVID-19 Therapy. Phytother. Res. 2022, 36, 2042–2060. [Google Scholar] [CrossRef] [PubMed]
- Conrad, K.P. Might Proton Pump or Sodium-Hydrogen Exchanger Inhibitors Be of Value to Ameliorate SARS-CoV-2 Pathophysiology? Physiol. Rep. 2021, 8, e14649. [Google Scholar] [CrossRef] [PubMed]
- Plassmeyer, M.; Alpan, O.; Corley, M.J.; Premeaux, T.A.; Lillard, K.; Coatney, P.; Vaziri, T.; Michalsky, S.; Pang, A.P.S.; Bukhari, Z.; et al. Caspases and Therapeutic Potential of Caspase Inhibitors in Moderate–Severe SARS-CoV-2 Infection and Long COVID. Allergy 2022, 77, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Ogletree, M.L.; Chander Chiang, K.; Kulshrestha, R.; Agarwal, A.; Agarwal, A.; Gupta, A. Treatment of COVID-19 Pneumonia and Acute Respiratory Distress With Ramatroban, a Thromboxane A2 and Prostaglandin D2 Receptor Antagonist: A Four-Patient Case Series Report. Front. Pharmacol. 2022, 13, 904020. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Archambault, A.; Zaid, Y.; Rakotoarivelo, V.; Turcotte, C.; Doré, É.; Dubuc, I.; Martin, C.; Flamand, O.; Amar, Y.; Cheikh, A.; et al. High Levels of Eicosanoids and Docosanoids in the Lungs of Intubated COVID-19 Patients. FASEB J. 2021, 35, e21666. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, T.; Matsui, T.; Okamoto, Y.; Ohta, A.; Shichijo, M. Ramatroban (BAY u 3405): A Novel Dual Antagonist of TXA2 Receptor and CRTh2, a Newly Identified Prostaglandin D2 Receptor. Cardiovasc. Drug Rev. 2004, 22, 71–90. [Google Scholar] [CrossRef]
- Trisciuzzi, D.; Siragusa, L.; Baroni, M.; Cruciani, G.; Nicolotti, O. An Integrated Machine Learning Model To Spot Peptide Binding Pockets in 3D Protein Screening. J. Chem. Inf. Model. 2022, 62, 6812–6824. [Google Scholar] [CrossRef] [PubMed]
- Rizk, J.G.; Kalantar-Zadeh, K.; Mehra, M.R.; Lavie, C.J.; Rizk, Y.; Forthal, D.N. Pharmaco-Immunomodulatory Therapy in COVID-19. Drugs 2020, 80, 1267–1292. [Google Scholar] [CrossRef] [PubMed]
- Alnemri, E.S.; Livingston, D.J.; Nicholson, D.W.; Salvesen, G.; Thornberry, N.A.; Wong, W.W.; Yuan, J. Human ICE/CED-3 Protease Nomenclature. Cell 1996, 87, 171. [Google Scholar] [CrossRef] [PubMed]
- Man, S.M.; Kanneganti, T.-D. Converging Roles of Caspases in Inflammasome Activation, Cell Death and Innate Immunity. Nat. Rev. Immunol. 2016, 16, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Imre, G. Cell Death Signalling in Virus Infection. Cell. Signal. 2020, 76, 109772. [Google Scholar] [CrossRef] [PubMed]
- Yapasert, R.; Khaw-on, P.; Banjerdpongchai, R. Coronavirus Infection-Associated Cell Death Signaling and Potential Therapeutic Targets. Molecules 2021, 26, 7459. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Han, M.; Li, T.; Sun, W.; Wang, D.; Fu, B.; Zhou, Y.; Zheng, X.; Yang, Y.; Li, X.; et al. Effective Treatment of Severe COVID-19 Patients with Tocilizumab. Proc. Natl. Acad. Sci. USA 2020, 117, 10970. [Google Scholar] [CrossRef]
- Spinner, C.D.; Gottlieb, R.L.; Criner, G.J.; Arribas López, J.R.; Cattelan, A.M.; Soriano Viladomiu, A.; Ogbuagu, O.; Malhotra, P.; Mullane, K.M.; Castagna, A.; et al. Effect of Remdesivir vs Standard Care on Clinical Status at 11 Days in Patients With Moderate COVID-19: A Randomized Clinical Trial. JAMA 2020, 324, 1048–1057. [Google Scholar] [CrossRef]
- Alijotas-Reig, J.; Esteve-Valverde, E.; Belizna, C.; Selva-O’Callaghan, A.; Pardos-Gea, J.; Quintana, A.; Mekinian, A.; Anunciacion-Llunell, A.; Miró-Mur, F. Immunomodulatory Therapy for the Management of Severe COVID-19. Beyond the Anti-Viral Therapy: A Comprehensive Review. Autoimmun. Rev. 2020, 19, 102569. [Google Scholar] [CrossRef]
- Grobler, C.; Maphumulo, S.C.; Grobbelaar, L.M.; Bredenkamp, J.C.; Laubscher, G.J.; Lourens, P.J.; Steenkamp, J.; Kell, D.B.; Pretorius, E. COVID-19: The Rollercoaster of Fibrin(Ogen), D-Dimer, Von Willebrand Factor, P-Selectin and Their Interactions with Endothelial Cells, Platelets and Erythrocytes. Int. J. Mol. Sci. 2020, 21, 5168. [Google Scholar] [CrossRef]
- Wilk, A.J.; Rustagi, A.; Zhao, N.Q.; Roque, J.; Martínez-Colón, G.J.; McKechnie, J.L.; Ivison, G.T.; Ranganath, T.; Vergara, R.; Hollis, T.; et al. A Single-Cell Atlas of the Peripheral Immune Response in Patients with Severe COVID-19. Nat. Med. 2020, 26, 1070–1076. [Google Scholar] [CrossRef]
- Linton, S.D.; Aja, T.; Armstrong, R.A.; Bai, X.; Chen, L.-S.; Chen, N.; Ching, B.; Contreras, P.; Diaz, J.-L.; Fisher, C.D.; et al. First-in-Class Pan Caspase Inhibitor Developed for the Treatment of Liver Disease. J. Med. Chem. 2005, 48, 6779–6782. [Google Scholar] [CrossRef]
- Stack, J.H.; Beaumont, K.; Larsen, P.D.; Straley, K.S.; Henkel, G.W.; Randle, J.C.R.; Hoffman, H.M. IL-Converting Enzyme/Caspase-1 Inhibitor VX-765 Blocks the Hypersensitive Response to an Inflammatory Stimulus in Monocytes from Familial Cold Autoinflammatory Syndrome Patients. J. Immunol. 2005, 175, 2630–2634. [Google Scholar] [CrossRef]
- Wei, Z.-Y.D.; Liang, K.; Shetty, A.K. Complications of COVID-19 on the Central Nervous System: Mechanisms and Potential Treatment for Easing Long COVID. Aging Dis. 2023, 14, 1492–1510. [Google Scholar] [CrossRef]
- Kim, J.; Zhang, J.; Cha, Y.; Kolitz, S.; Funt, J.; Escalante Chong, R.; Barrett, S.; Kusko, R.; Zeskind, B.; Kaufman, H. Advanced Bioinformatics Rapidly Identifies Existing Therapeutics for Patients with Coronavirus Disease-2019 (COVID-19). J. Transl. Med. 2020, 18, 257. [Google Scholar] [CrossRef]
- Alpan, O.; Gupta, R.; Latterich, M.; Hubka, M.; Bukhari, Z.; Ndhlovu, L. D020 role of cellular caspases and therapeutic potential of a pan-caspase inhibitor, emricasan, in COVID-19. Ann. Allergy Asthma Immunol. 2021, 127, S15–S16. [Google Scholar] [CrossRef]
- Garcia-Tsao, G.; Bosch, J.; Kayali, Z.; Harrison, S.A.; Abdelmalek, M.F.; Lawitz, E.; Satapathy, S.K.; Ghabril, M.; Shiffman, M.L.; Younes, Z.H.; et al. Randomized Placebo-Controlled Trial of Emricasan for Non-Alcoholic Steatohepatitis-Related Cirrhosis with Severe Portal Hypertension. J. Hepatol. 2020, 72, 885–895. [Google Scholar] [CrossRef]
- Shiffman, M.; Freilich, B.; Vuppalanchi, R.; Watt, K.; Chan, J.L.; Spada, A.; Hagerty, D.T.; Schiff, E. Randomised Clinical Trial: Emricasan versus Placebo Significantly Decreases ALT and Caspase 3/7 Activation in Subjects with Non-Alcoholic Fatty Liver Disease. Aliment. Pharmacol. Ther. 2019, 49, 64–73. [Google Scholar] [CrossRef]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.A.; Ruschitzka, F.; Moch, H. Endothelial Cell Infection and Endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef]
- Xu, S.; Ilyas, I.; Weng, J. Endothelial Dysfunction in COVID-19: An Overview of Evidence, Biomarkers, Mechanisms and Potential Therapies. Acta Pharmacol. Sin. 2023, 44, 695–709. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhao, T.; Zhou, X.; Xiang, Y.; Gutierrez-Castrellon, P.; Ma, X. Inflammatory Pathways in COVID-19: Mechanism and Therapeutic Interventions. MedComm 2022, 3, e154. [Google Scholar] [CrossRef]
- Uhlenhut, K.; Högger, P. Facilitated Cellular Uptake and Suppression of Inducible Nitric Oxide Synthase by a Metabolite of Maritime Pine Bark Extract (Pycnogenol). Free Radic. Biol. Med. 2012, 53, 305–313. [Google Scholar] [CrossRef]
- Zibadi, S.; Rohdewald, P.J.; Park, D.; Watson, R.R. Reduction of Cardiovascular Risk Factors in Subjects with Type 2 Diabetes by Pycnogenol Supplementation. Nutr. Res. 2008, 28, 315–320. [Google Scholar] [CrossRef]
- Weichmann, F.; Rohdewald, P. Projected Supportive Effects of PycnogenolⓇ in Patients Suffering from Multi-Dimensional Health Impairments after a SARS-CoV2 Infection. Int. J. Antimicrob. Agents 2020, 56, 106191. [Google Scholar] [CrossRef]
- Enseleit, F.; Sudano, I.; Périat, D.; Winnik, S.; Wolfrum, M.; Flammer, A.J.; Fröhlich, G.M.; Kaiser, P.; Hirt, A.; Haile, S.R.; et al. Effects of Pycnogenol on Endothelial Function in Patients with Stable Coronary Artery Disease: A Double-Blind, Randomized, Placebo-Controlled, Cross-over Study. Eur. Heart J. 2012, 33, 1589–1597. [Google Scholar] [CrossRef]
- Mihaila, R.G. A Minireview on NHE1 Inhibitors. A Rediscovered Hope in Oncohematology. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech Repub. 2015, 159, 519–526. [Google Scholar] [CrossRef]
- Gazmuri, R.J.; Radhakrishnan, J.; Ayoub, I.M. Sodium-Hydrogen Exchanger Isoform-1 Inhibition: A Promising Pharmacological Intervention for Resuscitation from Cardiac Arrest. Molecules 2019, 24, 1765. [Google Scholar] [CrossRef]
- Yeves, A.M.; Ennis, I.L. Na+/H+ Exchanger and Cardiac Hypertrophy. Hipertens. Y Riesgo Vasc. 2020, 37, 22–32. [Google Scholar] [CrossRef]
- Previtali, S.C.; Gidaro, T.; Díaz-Manera, J.; Zambon, A.; Carnesecchi, S.; Roux-Lombard, P.; Spitali, P.; Signorelli, M.; Szigyarto, C.A.-K.; Johansson, C.; et al. Rimeporide as a FIrst- in-Class NHE-1 Inhibitor: Results of a Phase Ib Trial in Young Patients with Duchenne Muscular Dystrophy. Pharmacol. Res. 2020, 159, 104999. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecules | Target(s)/Mechanism(s) of Action a | Phase of Clinical or Preclinical Investigation |
|---|---|---|
| Ramatroban | DPr2 and TPr receptor antagonist | Phase 2/3 |
| Emricasan | Pan-caspase inhibitor | Phase 1 (terminated) |
| Pycnogenol® | Antioxidant, eNOS | Phase 3 |
| Rimeporide | NHE-1 inhibitors | Preclinical investigation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samarelli, F.; Graziano, G.; Gambacorta, N.; Graps, E.A.; Leonetti, F.; Nicolotti, O.; Altomare, C.D. Small Molecules for the Treatment of Long-COVID-Related Vascular Damage and Abnormal Blood Clotting: A Patent-Based Appraisal. Viruses 2024, 16, 450. https://doi.org/10.3390/v16030450
Samarelli F, Graziano G, Gambacorta N, Graps EA, Leonetti F, Nicolotti O, Altomare CD. Small Molecules for the Treatment of Long-COVID-Related Vascular Damage and Abnormal Blood Clotting: A Patent-Based Appraisal. Viruses. 2024; 16(3):450. https://doi.org/10.3390/v16030450
Chicago/Turabian StyleSamarelli, Francesco, Giovanni Graziano, Nicola Gambacorta, Elisabetta Anna Graps, Francesco Leonetti, Orazio Nicolotti, and Cosimo Damiano Altomare. 2024. "Small Molecules for the Treatment of Long-COVID-Related Vascular Damage and Abnormal Blood Clotting: A Patent-Based Appraisal" Viruses 16, no. 3: 450. https://doi.org/10.3390/v16030450
APA StyleSamarelli, F., Graziano, G., Gambacorta, N., Graps, E. A., Leonetti, F., Nicolotti, O., & Altomare, C. D. (2024). Small Molecules for the Treatment of Long-COVID-Related Vascular Damage and Abnormal Blood Clotting: A Patent-Based Appraisal. Viruses, 16(3), 450. https://doi.org/10.3390/v16030450