
Viral Threats to Fruit and Vegetable Crops in the Caribbean
 , ,
, ,
Abstract
:1. Introduction
2. Endemic and Potential Re-Emerging Threats
2.1. Papaya Ringspot Potyvirus
2.2. Citrus Tristeza Closterovirus
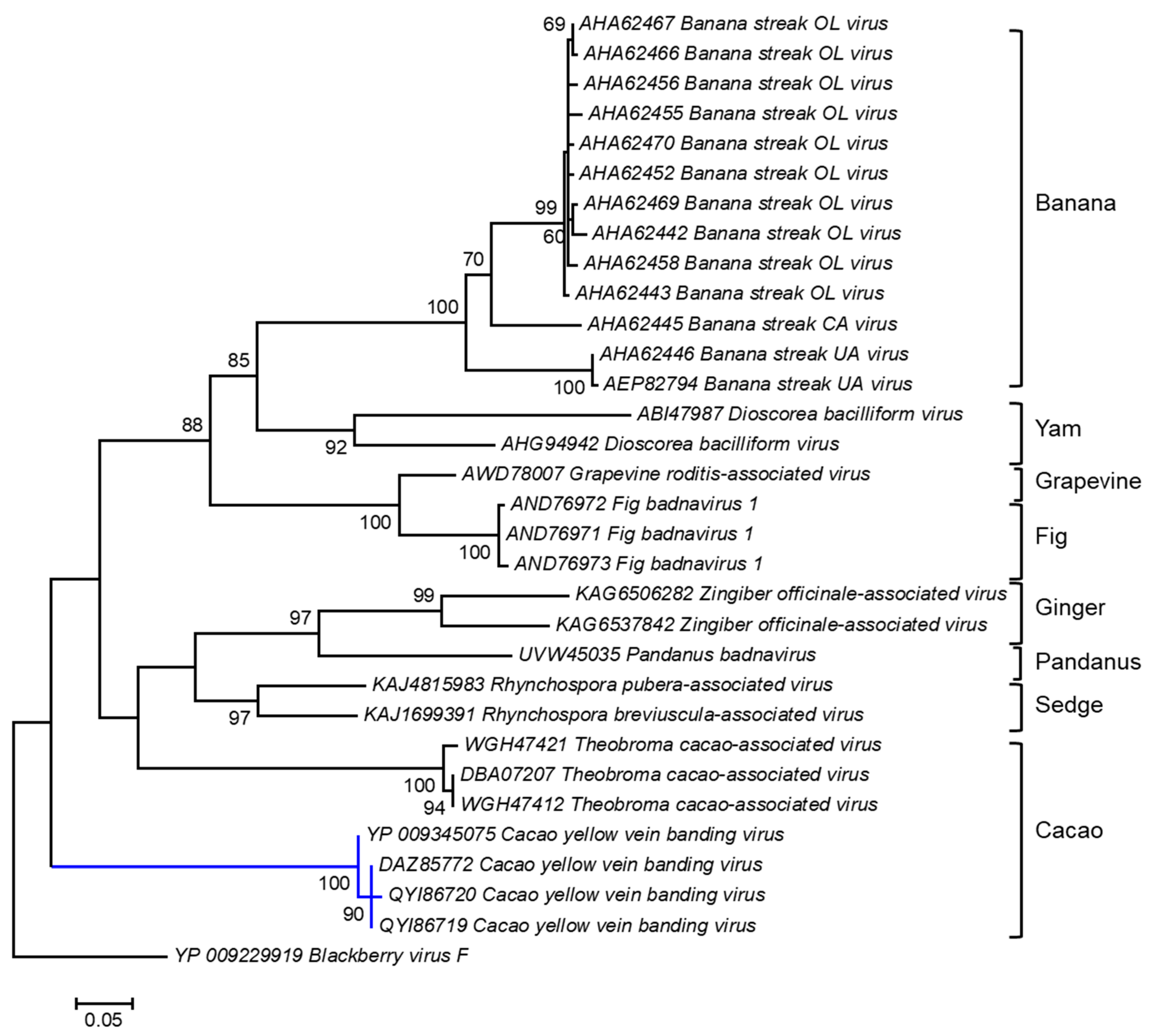
2.3. Cacao Mild Mosaic Badnavirus and Cacao Yellow Vein-Banding Badnavirus
3. Recent and Ongoing Threats
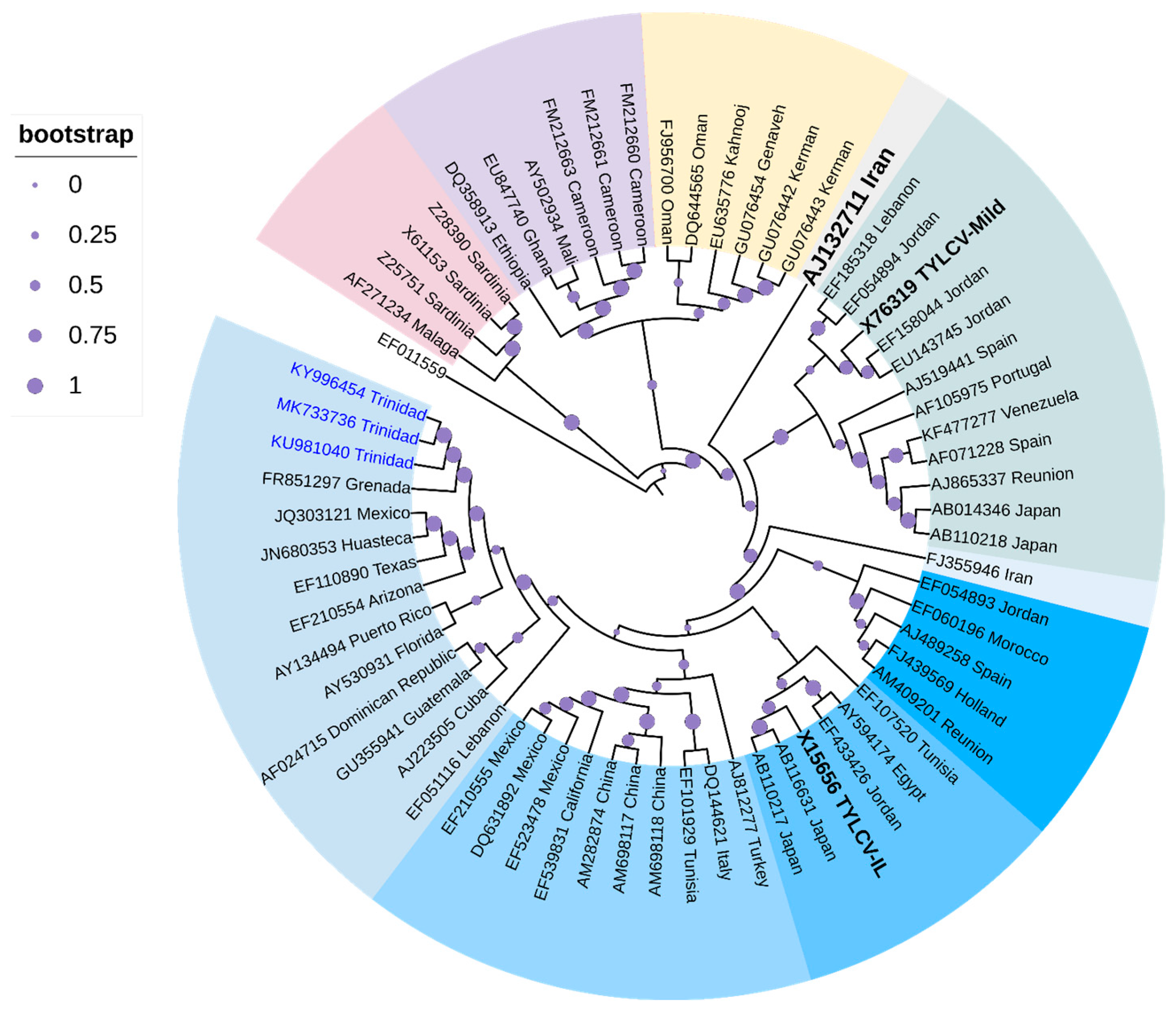
3.1. Tomato Yellow Leaf Curl Begomovirus
3.2. Sweet Potato Feathery Mottle Potyvirus and Sweet Potato Chlorotic Stunt Closterovirus
4. Newly Recognized Threats
4.1. Zucchini Yellow Mosaic Potyvirus
4.2. Cucurbit Yellow Stunting Disorder Crinivirus
5. Challenges and Perspectives
5.1. Quarantine and Biosecurity Measures
5.2. Early Detection and Diagnosis
5.3. Integrated Disease Management (IDM)
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fargette, D.; Konate, G.; Fauquet, C.; Muller, E.; Peterscmitt, M.; Thresh, J.M. Molecular Ecology and Emergence of Tropical Plant Viruses. Ann. Rev. Phytopathol. 2006, 44, 235–260. [Google Scholar] [CrossRef] [PubMed]
- Ristaino, J.B.; Anderson, P.K.; Bebber, D.P.; Brauman, K.A.; Cunniffe, N.J.; Fedoroff, N.V.; Finegold, C.; Garrett, K.A.; Gilligan, C.A.; Jones, C.M.; et al. The Persistent Threat of Emerging Plant Disease Pandemics to Global Food Security. Proc. Natl. Acad. Sci. USA 2021, 118, e2022239118. [Google Scholar] [CrossRef]
- Gilbertson, R.L.; Batuman, O.; Webster, C.G.; Adkins, S. Role of the Insect Supervectors Bemisia Tabaci and Frankliniella Occidentalis in the Emergence and Global Spread of Plant Viruses. Annu. Rev. Virol. 2015, 2, 67–93. [Google Scholar] [CrossRef]
- Tomlinson, K.R.; Bailey, A.M.; Alicai, T.; Seal, S.; Foster, G.D. Cassava Brown Streak Disease: Historical Timeline, Current Knowledge and Future Prospects. Mol. Plant Pathol. 2018, 19, 1282–1294. [Google Scholar] [CrossRef]
- Kumar, P.L.; Hanna, R.; Alabi, O.J.; Soko, M.M.; Oben, T.T.; Vangu, G.H.P.; Naidu, R.A. Banana Bunchy Top Virus in Sub-Saharan Africa: Investigations on Virus Distribution and Diversity. Virus Res. 2011, 159, 171–182. [Google Scholar] [CrossRef]
- Singh, K.; Wegulo, S.N.; Skoracka, A.; Kundu, J.K. Wheat Streak Mosaic Virus: A Century Old Virus with Rising Importance Worldwide. Mol. Plant Pathol. 2018, 19, 2193–2206. [Google Scholar] [CrossRef] [PubMed]
- Taliansky, M.E.; Robinson, D.J.; Murant, A.F. Groundnut Rosette Disease Virus Complex: Biology and Molecular Biology. Adv. Virus Res. 2000, 56, 357–400. [Google Scholar]
- Hanssen, I.M.; Lapidot, M.; Thomma, B.P. Emerging Viral Diseases of Tomato Crops. Mol. Plant-Microbe Interact. 2010, 23, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.A.; Davis, J.A.; Abad, J.A.; Cuellar, W.J.; Fuentes, S.; Kreuze, J.F.; Gibson, R.W.; Mukasa, S.B.; Tugume, A.K.; Tairo, F.D.; et al. Sweet potato Viruses: 15 Years of Progress on Understanding and Managing Complex Diseases. Plant Dis. 2012, 96, 168–185. [Google Scholar] [CrossRef]
- Karasev, A.V.; Gray, S.M. Continuous and Emerging Challenges of Potato Virus Y in potato. Annu. Rev. Phytopathol. 2013, 51, 571–586. [Google Scholar] [CrossRef]
- Legg, J.P.; Kumar, P.L.; Makeshkumar, T.; Tripathi, L.; Ferguson, M.; Kanju, E.; Ntawuruhunga, P.; Cuellar, W. Cassava Virus Diseases: Biology, Epidemiology, and Management. Adv. Virus Res. 2015, 91, 85–142. [Google Scholar] [PubMed]
- Tatineni, S.; Hein, G.L. Plant Viruses of Agricultural Importance: Current and Future Perspectives of Virus Disease Management Strategies. Phytopathology 2023, 113, 117–141. [Google Scholar] [CrossRef] [PubMed]
- Norris, R.F.; Kogan, M. Interactions between Weeds, Arthropod Pests, and their Natural Enemies in Managed Ecosystems. Weed Sci. 2000, 48, 94–158. [Google Scholar] [CrossRef]
- Wisler, G.C. Interactions between Weeds and Cultivated Plants as Related to Management of Plant Pathogens. Weed Sci. 2005, 53, 914–917. [Google Scholar] [CrossRef]
- Hasiów-Jaroszewska, B.; Boezen, D.; Zwart, M.P. Metagenomic Studies of Viruses in Weeds and Wild Plants: A Powerful Approach to Characterise Variable Virus Communities. Viruses 2021, 13, 1939. [Google Scholar] [CrossRef] [PubMed]
- Mangrauthia, S.K.; Singh Shakya, V.P.; Jain, R.K.; Praveen, S. Ambient Temperature Perception in Papaya for Papaya Ringspot Virus Interaction. Virus Genes 2009, 38, 429–434. [Google Scholar] [CrossRef]
- Ghini, R.; Bettiol, W.; Hamada, E. Diseases in Tropical and Plantation Crops as Affected by Climate Changes: Current Knowledge and Perspectives. Plant Pathol. 2011, 60, 122–132. [Google Scholar] [CrossRef]
- Singh, B.K.; Delgado-Baquerizo, M.; Egidi, E.; Guirado, E.; Leach, J.E.; Liu, H.; Trivedi, P. Climate Change Impacts on Plant Pathogens, Food Security and Paths Forward. Nat. Rev. Microbiol. 2023, 21, 640–656. [Google Scholar] [CrossRef] [PubMed]
- Rodoni, B. The Role of Plant Biosecurity in Preventing and Controlling Emerging Plant Virus Disease Epidemics. Virus Res. 2009, 141, 150–157. [Google Scholar] [CrossRef]
- United Nations Environment Programme (UNEP); Villasol, A.; Beltran, J. Caribbean Islands, GIWA Regional Assessment 4, Global International Waters Assessment; Fortnam, M., Blime, P., Eds.; University of Kalmar: Kalmar, Sweden, 2004. [Google Scholar]
- Taylor, M.A.; Enfield, D.B.; Chen, A.A. Influence of the Tropical Atlantic versus the Tropical Pacific on Caribbean Rainfall. J. Geophys. Res. Ocean. 2002, 107, 10–11. [Google Scholar] [CrossRef]
- Almazroui, M.; Islam, M.N.; Saeed, F.; Saeed, S.; Ismail, M.; Ehsan, M.A.; Diallo, I.; O’Brien, E.; Ashfaq, M.; Martínez-Castro, D.; et al. Projected Changes in Temperature and Precipitation over the United States, Central America, and the Caribbean in CMIP6 GCMs. Earth Syst. Environ. 2021, 5, 1–24. [Google Scholar] [CrossRef]
- Taylor, M.A.; Stephenson, T.S.; Chen, A.A.; Stephenson, K.A. Climate Change and the Caribbean: Review and Response. Caribb. Stud. 2012, 40, 169–200. [Google Scholar] [CrossRef]
- Stephenson, T.S.; Vincent, L.A.; Allen, T.; Van Meerbeeck, C.J.; McLean, N.; Peterson, T.C.; Taylor, M.A.; Aaron-Morrison, A.P.; Auguste, T.; Bernard, D. Changes in Extreme Temperature and Precipitation in the Caribbean Region, 1961–2010. Int. J. Climatol. 2014, 34, 2957–2971. [Google Scholar] [CrossRef]
- Teycheney, P.-Y.; Abadie, C.; Lotti, J. Development of Plant Epidemiological Surveillance Networks, Data Exchanges and Joint Response Strategies in the Caribbean: The French Experience; No. 1876-2017-321; Caribbean Food Crops Society: Frigate Bay, St. Kitts and Nevis, 2009. [Google Scholar]
- Morales, F.J. Distribution and Dissemination of Begomoviruses in Latin America and the Caribbean. In Bemisia: Bionomics and Management of a Global Pest; Stansly, P.A., Naranjo, S.E., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 283–318. [Google Scholar]
- World Development Indicators. Available online: http://data.worldbank.org/data-catalog/world-development-indicators (accessed on 13 December 2023).
- Tandon, H. Strengthening Sustainable Agriculture in the Caribbean. A Report for the FAO. 2014. Available online: https://www.competecaribbean.org/documents/strengthening-sustainable-agriculture-in-the-caribbean/ (accessed on 18 December 2023).
- Barker, D. Caribbean Agriculture in a Period of Global Change: Vulnerabilities and Opportunities. Caribb. Stud. 2012, 40, 41–61. [Google Scholar] [CrossRef]
- Beckford, C.L.; Campbell, D.R. The Role of Agriculture in Caribbean Economies: A Historical and Contemporary Analysis. In Domestic Food Production and Food Security in the Caribbean: Building Capacity and Strengthening Local Food Production Systems; Beckford, C.L., Campbell, D.R., Eds.; Palgrave Macmillan: New York, NY, USA, 2013; pp. 3–12. [Google Scholar]
- Pierre, R.E. Observations on the Golden Mosaic Symptoms of Beans (Phaseolus vulgaris L.) in Jamaica. In Tropical Diseases in Legumes; Bird, J., Maramorosch, K., Eds.; Academic Press: New York, NY, USA, 1975; pp. 55–59. [Google Scholar]
- McLaughlin, W.; Rojas, M.R.; Nakhla, M.K.; Hidayat, S.; Maxwell, D.P. Partial Molecular Characterization of Bean Golden Mosaic Virus Isolates from Jamaica and Central America. Plant Dis. 1994, 78, 1220. [Google Scholar] [CrossRef]
- Roye, M.E. Genetic Diversity and Phylogeny of Whitefly-transmitted Geminivirues from Jamaica. Ph.D. Thesis, The University of the West Indies, Mona, St. Andrew, Jamaica, 1996. [Google Scholar]
- Posnette, A.F. Viruses of Cacao in Trinidad. Trop. Agric. 1944, 21, 105–106. [Google Scholar]
- Baker, R.E.D.; Dale, W.T. Notes on a Virus Disease of Cacao. Ann. Appl. Biol. 1947, 34, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Chingandu, N.; Zia-Ur-Rehman, M.; Sreenivasan, T.N.; Surujdeo-Maharaj, S.; Umaharan, P.; Gutierrez, O.A.; Brown, J.K. Molecular Characterization of Previously Elusive Badnaviruses Associated with Symptomatic Cacao in the New World. Arch. Virol. 2017, 162, 1363–1371. [Google Scholar] [CrossRef]
- Stell, G. First Record of Tristeza. FAO Plant Prot. Bull. 1961, 9, 85. [Google Scholar]
- Hosein, I. Citrus Virus Disease of Trinidad, Jamaica and British Honduras. In Proceedings of the Second Conference of the International Organization of Citrus Virologists, Riverside, CA, USA, 16–25 September 1963; University of Florida Press: Gainesville, FL, USA, 1965; pp. 228–231. [Google Scholar]
- Lee, R.F.; McConnell, P.; Manjunath, K.L.; Cevik, B.; Nikolaeva, O.V.; Dekkers, M.G.H.; Niblett, C.L. The Citrus Tristeza Virus Epidemic in Bog Walk Valley, Jamaica. In Proceedings of the 15th Conference of the International Organization of Citrus Virologists, Riverside, CA, USA, 11–16 November 2001; University of Florida Press: Gainesville, FL, USA, 2002; pp. 95–101. [Google Scholar]
- Nolasco, G.; Santos, C.; Silva, G.; Fonesca, F. Development of an Asymmetric PCR-ELISA Typing Method for Citrus Tristeza Virus Based on the Coat Protein Gene. J. Virol. Methods 2009, 155, 97–108. [Google Scholar] [CrossRef]
- Fisher, L.C.; Tennant, P.F.; McLaughlin, W.A. Detection and Characterization of Citrus Tristeza Virus Stem Pitting Isolates in Jamaica. Eur. J. Plant Pathol. 2010, 127, 1–6. [Google Scholar] [CrossRef]
- Fisher, L.; Tennant, P.; Mc Laughlin, W. Diversity of Citrus Tristeza Virus in Jamaica. J. Plant Pathol. 2013, 95, 201–206. [Google Scholar]
- CABI Compendium. Available online: https://www.cabidigitallibrary.org/journal/cabicompendium (accessed on 22 November 2023).
- Jensen, D. Papaya Virus Diseases with Special Reference to Papaya Ringspot. Phytopathology 1949, 39, 191–211. [Google Scholar]
- Chin, M.; Ahmad, M.H.; Tennant, P. Momordica Charantia is a Weed Host Reservoir for Papaya Ringspot Virus Type P in Jamaica. Plant Dis. 2007, 91, 1518. [Google Scholar] [CrossRef]
- Leather, R.I. A Catalogue of Some Plant Diseases and Fungi in Jamaica; Bulletin Number 61 (New Series); Ministry of Agriculture and Lands: Kingston, Jamaica, 1967; pp. 1–92. [Google Scholar]
- Ministry of Agriculture Investigations. Plant Pathology, Other Diseases Investigated, Tomato. Bulletin 1951, 49, 89. [Google Scholar]
- Turner, M.P.; Edman, F.; McKenzie, P. Plant Viruses Disease Survey of Jamaica. Jam. Minist. Agric. Investig. Bull. 1976, 66, 14–18. [Google Scholar]
- Turner, M.P.; Stace-Smith, R. Plant-Virus Disease Survey of Jamaica. FAO Plant Prot. Bull. 1979, 27, 9–11. [Google Scholar]
- Myers, L.R.S. The Etiology of Viruses Affecting Pepper (Capsicum spp.) in Jamaica. M.Phil. Thesis, University of the West Indies, Kingston, Jamaica, 1996. [Google Scholar]
- Evans, T.A.; Keil, C.A. Survey of Scotch Bonnet Peppers in Jamaica for Plant Viruses and Insect Pests. J. Environ. Monit. Restor. 2010, 6, 68–73. [Google Scholar]
- Roye, M.E.; Brown, S.; Spence, J.D.; Smith, K.; McLaughlin, W.A.; Brown, J.K.; Maxwell, D.P. Ten Years of Molecular Biology Research on Whitefly-Transmitted Geminiviruses from Jamaica: A Review. JAMST J. Sci Technol. 2003, 14, 98–118. [Google Scholar]
- Smith, K.N. Molecular Characterization, Recombination and Distribution of Cabbage Leaf Curl Virus from Jamaica. M.Phil. Thesis, University of the West Indies, Mona, St. Andrew, Jamaica, 2005. [Google Scholar]
- Umaharan, P.; Padidam, M.; Phelps, R.H.; Beachy, R.N.; Fauquet, C.M. Distribution and Diversity of Geminiviruses in Trinidad and Tobago. Phytopathology 1998, 88, 1262–1268. [Google Scholar] [CrossRef]
- McGlashan, D.; Polston, J.E.; Bois, D. Tomato Yellow Leaf Curl Geminivirus in Jamaica. Plant Dis. 1994, 78, 1219. [Google Scholar] [CrossRef]
- Roye, M.E.; Wernecke, M.E.; McLaughlin, W.A.; Nakhla, M.K.; Maxwell, D.P. Tomato Dwarf Leaf Curl Virus, a New Bipartite Geminivrius Associated with Tomatoes and Peppers in Jamaica and Mixed Infection with Tomato Yellow Leaf Curl Virus. Plant Pathol. 1999, 48, 370–378. [Google Scholar] [CrossRef]
- Roye, M.E.; Henry, N.M.; Burrell, P.D.; McLaughlin, W.A.; Nakhla, M.K.; Maxwell, D.P. A New Tomato-Infecting Begomovirus in Barbados. Plant Dis. 2000, 84, 1342. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, P.D.; McLaughlin, W.A.; Maxwell, D.P.; Amarakoon, I.I.; Roye, M.E. Detection of Begomoviruses in Caribbean Crops and Weeds in the Islands of Antigua, Barbados and St Kitts and Nevis. Trop. Agric. 2011, 41, 40211–40214. [Google Scholar]
- Chinnaraja, C.; Ramsubhag, A.; Jayaraj, J. Identification of Tomato Yellow Leaf Curl Virus Infecting Cowpea in Trinidad. Plant Dis. 2017, 101, 1830. [Google Scholar] [CrossRef]
- Johnson, L.; Tennant, P. Viruses infecting Sweet Potato in Jamaica. In Proceedings of the 16th Triennial Symposium, International Society for Tropical Root Crops, Abeokuta, Nigeria, 24–28 September 2012; Nigeria Federal University of Agriculture: Abeokuta, Nigeria; p. 10. [Google Scholar]
- Johnson, J. Detection and Diversity of Select Sweet Potato Viruses in Two Caribbean Countries, Jamaica and Trinidad and Tobago. Ph.D. Thesis, University of the West Indies, Mona, St. Andrew, Jamaica, 2022. [Google Scholar]
- Li, R.; Salih, S.; Hurtt, S. Detection of Geminiviruses in Sweetpotato by Polymerase Chain Reaction. Plant Dis. 2004, 88, 1347–1351. [Google Scholar] [CrossRef] [PubMed]
- Alleyne, A.T.; Cummins, C.; Rowe, K.; James, M.; Gutiérrez, D.L.; Fuentes, S. Sequencing and Assembly of Small RNAs Reveal the Presence of Several Begomoviruses, Potyviruses, Badnaviruses and Mastreviruses in the Sweet Potato Leaf Virome in Barbados. J. Plant Pathol. 2019, 101, 339–347. [Google Scholar] [CrossRef]
- Myers, L.; Murray, A. The Detection, Identification and Incidence of Sweet Potato Viruses in Jamaica; Inter-American Institute for Cooperation on Agriculture (IICA): Kingston, Jamaica, 2006; pp. 1–9. [Google Scholar]
- James, M.; Phillip, M.T.; Fuentes, S.; Salazar, L.F. A Study on the Cause of Sweet Potato Virus Disease in Barbados. Bridgetown. In Proceedings of the Barbados National Agricultural Conference 2003: Agricultural Research and Development, Meeting the Challenges of the 21st Century, Bridgetown, Barbados, 5–6 May 2003; p. 10. [Google Scholar]
- Gooding, G.V. Report to Agro 21 on Viruses of Cucurbits, Peppers and Tobacco; The AGRO 21 Programme: Kingston, Jamaica, 1985. [Google Scholar]
- Thomas, L.S. Distribution and Characterization of Pumpkin (Cucurbita pepo) Diseases in Jamaica. M.Phil. Thesis, University of the West Indies, Mona, St. Andrew, Jamaica, 2014. [Google Scholar]
- Chinnaraja, C.; Ramkissoon, A.; Rajendran, R.; Tony, S.T.; Ramsubhag, A.; Jayaraj, J. First Report of Zucchini Yellow Mosaic Virus and Squash Mosaic Virus Infecting Cucurbits in Trinidad. Plant Dis. 2016, 100, 866. [Google Scholar] [CrossRef]
- Pitter, P.; Mondal, S.; Chang, P.-G.; Myers Morgan, L.; Aikman, S.; Wintermantel, W.M.; Tennant, P. First Report of Cucurbit Yellow Stunting Disorder Virus Infecting Cucurbit Crops in Jamaica. Plant Dis. 2024; accepted. [Google Scholar] [CrossRef]
- Wang, C.H.; Yeh, S.-D. Divergence and Conservation of the Genomic RNAs of Taiwan and Hawaii Strains of Papaya Ringspot Potyvirus. Arch. Virol. 1997, 142, 271–285. [Google Scholar]
- Yeh, S.; Jan, F.; Chiang, C.; Doong, T.; Chen, M.; Chung, P.; Bau, H. Complete Nucleotide Sequence and Genetic Organization of Papaya Ringspot Virus RNA. J. Gen. Virol. 1992, 73, 2531–2541. [Google Scholar] [CrossRef] [PubMed]
- Purcifull, D.; Edwardson, J.; Hiebert, E.; Gonsalves, D. Papaya Ringspot Virus. In CMI/AAB Description of Plant Viruses; Coronel, R.E., Ed.; Wageningen University: Wageningen, The Netherlands, 1998; Volume 2, p. 8. [Google Scholar]
- Gonsalves, D. Papaya Diseases Caused by Viruses. In Compendium of Tropical Fruit Diseases; Ploet, R.C., Zentmyer, G.A., Nishijima, W.T., Rohrbach, K.G., Ohr, H.D., Eds.; APS Press: St. Paul, MN, USA, 1994; pp. 67–68. [Google Scholar]
- Bayot, R.G.; Villegas, V.N.; Magdalita, P.M.; Jovellana, M.D.; Espino, T.M.; Exconde, S.B. Seed transmissibility of Papaya Ringspot Virus. Philipp. J. Crop Sci. 1990, 15, 107–111. [Google Scholar]
- Ferwerda-Licha, M.; Pingel, R.L. Ongoing Survey on the Incidence of Papaya Ringspot Virus in Weeds Found in a Papaya Field in Northern Puerto Rico. Phytopathology 2003, 93, 525. [Google Scholar]
- Noa-Carrazana, J.C.; González-de-León, D.; Ruiz-Castro, B.S.; Piñero, D.; Silva-Rosales, L. Distribution of Papaya Ringspot Virus and Papaya Mosaic Virus in Papaya Plants (Carica papaya) in Mexico. Plant Dis. 2006, 90, 1004–1011. [Google Scholar] [CrossRef] [PubMed]
- Laney, A.G.; Avanzato, M.V.; Tzanetakis, I.E. High Incidence of Seed Transmission of Papaya Ringspot Virus and Watermelon Mosaic Virus, Two Viruses Newly Identified in Robinia Pseudoacacia. Eur. J. Plant Pathol. 2012, 134, 227–230. [Google Scholar] [CrossRef]
- Young, F. Papaya Ringspot Virus in Jamaica. In Proceedings of the Papaya Industry Seminar, Kingston, Jamaica, 24–26 October 1994; pp. 39–46. [Google Scholar]
- Bateson, M.F.; Lines, R.E.; Revill, P.; Chaleeprom, W.; Ha, C.V.; Gibbs, A.J.; Dale, J.L. On the Evolution and Molecular Epidemiology of the Potyvirus Papaya Ringspot Virus. J. Gen. Virol. 2002, 83, 2575–2585. [Google Scholar] [CrossRef] [PubMed]
- Olarte Castillo, X.A.; Fermin, G.; Tabima, J.; Rojas, Y.; Tennant, P.F.; Fuchs, M.; Sierra, R.; Bernal, A.J.; Restrepo, S. Phylogeography and Molecular Epidemiology of Papaya Ringspot Virus. Virus Res. 2011, 159, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Desbiez, C.; Wipf-Scheibel, C.; Millot, P.; Verdin, E.; Dafalla, G.; Lecoq, H. New species in the Papaya Ringspot Virus Cluster: Insights into the Evolution of the PRSV Lineage. Virus Res. 2017, 241, 88–94. [Google Scholar] [CrossRef]
- Babu, K.S.; Banerjee, A. Biological and Molecular Evidence of Papaya Ring Spot Virus Pathotype P from Mid-Hills of Meghalaya, India. Indian Phytopathol. 2018, 71, 611–620. [Google Scholar] [CrossRef]
- Maina, S.; Barbetti, M.J.; Edwards, O.R.; Minemba, D.; Areke, M.W.; Jones, R.A.C. Genetic Connectivity between Papaya Ringspot Virus Genomes from Papua New Guinea and Northern Australia, and New Recombination Insights. Plant Dis. 2019, 103, 737–747. [Google Scholar] [CrossRef]
- Coates Beckford, P.; Tennant, P. Papaya. In Plant Disease: The Jamaican Experience; The University of the West Indies Press: St. Andrew, Jamaica, 2013; pp. 223–249. [Google Scholar]
- Tennant, P.F.; Gonsalves, C.; Ling, K.S.; Fitch, M.; Manshardt, R.; Slightom, J.L.; Gonsalves, D. Differential Protection against Papaya Ringspot Virus Isolates in Coat Protein Gene Transgenic Papaya and Classically Cross-Protected Papaya. Phytopathology 1994, 84, 1359–1365. [Google Scholar] [CrossRef]
- Tennant, P.F.; Ahmad, M.H.; Gonsalves, D. Transformation of Carica papaya L. with Virus Coat Protein Genes for Studies on Resistance to Papaya Ringspot Virus from Jamaica. Trop. Agric. 2002, 79, 1–9. [Google Scholar]
- Tennant, P.; Ahmad, M.H.; Gonsalves, D. Field Resistance of Coat Protein Transgenic Papaya to Papaya Ringspot Virus in Jamaica. Plant Dis. 2005, 89, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Fermin, G.; Tennant, P. Opportunities and Constraints to Biotechnological Applications in the Caribbean: Transgenic Papayas in Jamaica and Venezuela. Plant Cell Rep. 2011, 30, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yeh, S.; Chiu, R.; Gonsalves, D. Effectiveness of Cross-Protection by Mild Mutants of Papaya Ringspot for Control of Ringspot Disease of Papaya in Taiwan. Plant Dis. 1987, 71, 491–497. [Google Scholar] [CrossRef]
- Magdalita, P.; Opina, O.; Espino, R.; Villegas, V. Epidemiology of Papaya Ringspot Virus in the Philippines. Philipp. Phytopathol. Soc. 1988, 25, 1–11. [Google Scholar]
- Teliz, D.; Nieto, D.; Flores, C.; Garcia, E.; Villanueva, J. Integrated Management of Papaya in Mexico. In Proceedings of the International Symposium on Tropical Fruits, Vitória, Brazil, 7–12 November 1993; p. 42. [Google Scholar]
- Folimonova, S.Y.; Sun, Y.D. Citrus Tristeza Virus: From Pathogen to Panacea. Ann. Rev. Virol. 2022, 9, 417–435. [Google Scholar] [CrossRef] [PubMed]
- Moreno, P.; Ambrós, S.; Albiach-Martí, M.R.; Guerri, J.; Peña, L. Citrus Tristeza Virus: A Pathogen that Changed the Course of the Citrus Industry. Mol. Plant Pathol. 2008, 9, 251–268. [Google Scholar] [CrossRef]
- Lee, R.F.; Bar-Joseph, M. Tristeza. In Compendium of Citrus Diseases; Timmer, L.W., Garnsey, S.M., Graham, J.H., Eds.; The American Phytopathological Society: St. Paul, MN, USA, 2000; pp. 61–63. [Google Scholar]
- Liu, Q.; Zhang, S.; Mei, S.; Zhou, Y.; Wang, J.; Han, G.Z.; Chen, L.; Zhou, C.; Cao, M. Viromics Unveils Extraordinary Genetic Diversity of the Family Closteroviridae in Wild Citrus. PLOS Pathog. 2021, 17, e1009751. [Google Scholar] [CrossRef]
- Costa, A.S. Present Status of the Tristeza Disease of Citrus in South America. FAO Plant Prot. Bull. 1956, 4, 97–105. [Google Scholar]
- Rocha-Peña, M.A.; Lee, R.F.; Lastra, R.; Niblett, C.L.; Ochoa-Corona, F.M.; Garnsey, S.M.; Yokomi, R.K. Citrus Tristeza Virus and its Aphid Vector Toxoptera Citricida: Threats to Citrus Production in the Caribbean and Central and North America. Plant Dis. 1995, 79, 437–445. [Google Scholar] [CrossRef]
- Coates Beckford, P.; Tennant, P. Citrus. In Plant Disease: The Jamaican Experience; The University of the West Indies Press: St. Andrew, Jamaica, 2013; pp. 85–141. [Google Scholar]
- Persad, A.; Hoy, M.; Nguyen, R. Establishment of Lipolexis oregmae (Hymenoptera: Aphidiidae) in a Classical Biological Control Program Directed against the Brown Citrus Aphid (Homoptera: Aphididae) in Florida. Fla. Entomol. 2007, 90, 204–213. [Google Scholar] [CrossRef]
- Hoy, M.A.; Jeyaprakash, A.; Clarke-Harris, D.; Rhodes, L. Molecular and Field Analyses of the Fortuitous Establishment of Lipolexis oregmae (Hymenoptera: Aphidiidae) in Jamaica as a Natural Enemy of the Brown Citrus Aphid. Biocontrol. Sci. Technol. 2007, 17, 473–482. [Google Scholar] [CrossRef]
- Motamayor, J.C.; Risterucci, A.M.; Lopez, P.A.; Ortiz, C.F.; Moreno, A.; Lanaud, C. Cacao Domestication I: The Origin of the Cacao Cultivated by the Mayas. Heredity 2002, 89, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Sobrinho, R.; Chingandu, N.; Gutierrez, O.A.; Marelli, J.-P.; Brown, J.K. A Complex of Badnavirus Species Infecting Cacao Reveals Mixed Infections, Extensive Genomic Variability, and Interspecific Recombination. Viruses 2020, 12, 443. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.; Hohn, T.; Selvarajan, R. Badnaviruses: The Current Global Scenario. Viruses 2016, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.; Selvarajan, R.; Balasubramanian, V. Emerging and Re-Emerging Diseases Caused by Badnaviruses. Pathogens 2023, 12, 245. [Google Scholar] [CrossRef]
- Harper, G.; Hull, R. Cloning and Sequence Analysis of Banana Streak Virus DNA. Virus Genes 1998, 17, 271278. [Google Scholar] [CrossRef]
- Muller, E.; Ravel, S.; Agret, C.; Abrokwah, F.; Dzahini-Obiatey, H.; Galyuon, I.; Kouakou, K.; Jeyaseelan, E.C.; Allainguillaume, J.; Wetten, A. Next Generation Sequencing Elucidates Cacao Badnavirus Diversity and Reveals the Existence of More Than Ten Viral Species. Virus Res. 2018, 244, 235–251. [Google Scholar] [CrossRef]
- Ullah, I.; Daymond, A.J.; Hadley, P.; End, M.J.; Umaharan, P.; Dunwell, J.M. Identification of Cacao Mild Mosaic Virus (CaMMV) and Cacao Yellow Vein-Banding Virus (CYVBV) in Cocoa (Theobroma cacao) Germplasm. Viruses 2021, 13, 2152. [Google Scholar] [CrossRef]
- Kirkpatrick, T.W. Insect Transmission of Cacao Virus Disease in Trinidad. Bull. Entomol. Res. 1950, 41, 99. [Google Scholar] [CrossRef]
- Sreenivasan, T.N. The Enigma of the ICS 76 Plants at Reading; UK Baker RED: Reading, UK, 2009. [Google Scholar]
- Ciferri, R. Una Virosis del Cacao en Colombia y en la República Dominicana. Rev. Fac. Nac. Agron. Medellín 1948, 8, 79–84. [Google Scholar]
- Posnette, A.F.; Palma, M. Observations on Cacao in the Paria Peninsula, Venezuela. Trop. Agric. 1944, 21, 104. [Google Scholar]
- Katzourakis, A.; Gifford, R.J. Endogenous Viral Elements in Animal Genomes. PLoS Genet. 2010, 6, e1001191. [Google Scholar] [CrossRef]
- Ullah, I.; Dunwell, J.M. Bioinformatic, Genetic and Molecular Analysis of Several Badnavirus Sequences Integrated in the Genomes of Diverse Cacoa (Theobroma cacao L.) Germplasm. Saudi J. Biol. Sci. 2023, 30, 103648. [Google Scholar] [CrossRef] [PubMed]
- Le, S.Q.; Gascuel, O. An Improved General Amino Acid Replacement Matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Abrokwah, F.; Dzahini-Obiatey, H.; Galyuon, I.; Osae-Awuku, F.; Muller, E. Geographical Distribution of Cacao Swollen Shoot Virus Molecular Variability in Ghana. Plant Dis. 2016, 100, 2011–2017. [Google Scholar] [CrossRef] [PubMed]
- Muller, E.; Ullah, I.; Dunwell, J.M.; Daymond, A.J.; Richardson, M.; Allainguillaume, J.; Wetten, A. Identification and Distribution of Novel Badnaviral Sequences Integrated in the Genome of Cacao (Theobroma cacao). Sci. Rep. 2021, 11, 8270. [Google Scholar] [CrossRef]
- End, M.J.; Daymond, A.J.; Hadley, P. Technical Guidelines for the Safe Movement of Cacao Germplasm; Global Cacao Genetic Resources Network (CacaoNet): Rome, Italy, 2017; Revised from the FAO/IPGRI Technical Guidelines No. 20 (Third Update, October 2017). [Google Scholar]
- Mauck, K.E. Variation in Virus Effects on Host Plant Phenotypes and Insect Vector Behavior: What Can It Teach Us about Virus Evolution? Curr. Opin. Virol. 2016, 21, 114–123. [Google Scholar] [CrossRef]
- Mahatma, L.; Mahatma, M.K.; Pandya, J.R.; Solanki, R.K.; Solanki, V.A. Epidemiology of Begomoviruses: A Global Perspective. In Plant Viruses: Evolution and Management; Gaur, R., Petrov, N., Patil, B., Stoyanova, M., Eds.; Springer: Singapore, 2016; pp. 171–188. [Google Scholar]
- Yan, Z.; Wolters, A.A.; Navas-Castillo, J.; Bai, Y. The Global Dimension of Tomato Yellow Leaf Curl Disease: Current Status and Breeding Perspectives. Microorganisms 2021, 9, 740. [Google Scholar] [CrossRef] [PubMed]
- Padidam, M.; Sawyer, S.; Fauquet, C.M. Possible Emergence of New Geminiviruses by Frequent Recombination. Virology 1999, 265, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Briddon, R.W.; Patil, B.L.; Bagewadi, B.; Nawaz-ul-Rehman, M.S.; Fauquet, C.M. Distinct Evolutionary Histories of the DNA-A and DNA-B Components of Bipartite Begomoviruses. BMC Evol. Biol. 2010, 10, 97. [Google Scholar] [CrossRef] [PubMed]
- Briddon, R.W.; Stanley, J. Subviral Agents Associated with Plant Single-Stranded DNA Viruses. Virology 2006, 344, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Patil, B.L.; Dasgupta, I. Defective Interfering DNAs of Plant Viruses. Crit. Rev. Plant Sci. 2006, 25, 47–64. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Martínez-Zubiaur, Y.; Moriones, E.; Navas-Castillo, J. A Novel Class of DNA Satellites Associated with New World Begomoviruses. Virology 2012, 426, 1–6. [Google Scholar] [CrossRef]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging Virus Diseases Transmitted by Whiteflies. Ann. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef] [PubMed]
- Wisler, G.C.; Duffus, J.E.; Liu, H.Y.; Li, R.H. Ecology and Epidemiology of Whitefly-Transmitted Closteroviruses. Plant Dis. 1998, 82, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, Y.; Zchori-Fein, E.; Mozes-Daube, N.; Kontsedalov, S.; Skaljac, M.; Brumin, M.; Sobol, I.; Czosnek, H.; Vavre, F.; Fleury, F.; et al. The Transmission Efficiency of Tomato Yellow Leaf Curl Virus by the whitefly Bemisia tabaci is Correlated with the Presence of a Specific Symbiotic Bacterium Species. J. Virol. 2010, 84, 9310–9317. [Google Scholar] [CrossRef]
- Czosnek, H.; Hariton-Shalev, A.; Sobol, I.; Gorovits, R.; Ghanim, M. The Incredible Journey of Begomoviruses in Their Whitefly Vector. Viruses 2017, 9, 273. [Google Scholar] [CrossRef]
- Mabvakure, B.; Martin, D.P.; Kraberger, S.; Cloete, L.; van Brunschot, S.; Geering, A.D.; Thomas, J.E.; Bananej, K.; Lett, J.M.; Lefeuvre, P.; et al. Ongoing Geographical Spread of Tomato Yellow Leaf Curl Virus. Virology 2016, 498, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Muniz, Y.; Granier, M.; Caruth, C.; Umaharan, P.; Marchal, C.; Pavis, C.; Wicker, E.; Martinez, Y.; Peterschmitt, M. Extensive Settlement of the Invasive MEAM1 Population of Bemisia tabaci (Hemiptera: Aleyrodidae) in the Caribbean and Rare Detection of Indigenous Populations. Environ. Entomol. 2011, 40, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Fiallo-Olivé, E.; Lett, J.M.; Martin, D.P.; Roumagnac, P.; Varsani, A.; Zerbini, F.M.; Navas-Castillo, J. ICTV Report Consortium, 2021. ICTV virus taxonomy profile: Geminiviridae. J. Gen. Virol. 2021, 102, 001696. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Shu, Y.-N.; Liu, C.; Chi, Y.; Liu, Y.-Q.; Wang, X.-W. Transovarial Transmission of Tomato Yellow Leaf Curl Virus by Seven Species of the Bemisia tabaci Complex Indigenous to China: Not All Whiteflies Are the Same. Virology 2019, 531, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Kil, E.-J.; Kim, S.; Lee, Y.-J.; Byun, H.-S.; Park, J.; Seo, H.; Kim, C.-S.; Shim, J.-K.; Lee, J.-H.; Kim, J.-K.; et al. Tomato Yellow Leaf Curl Virus (TYLCV-IL): A Seed-Transmissible Geminivirus in Tomatoes. Sci. Rep. 2016, 6, 19013. [Google Scholar] [CrossRef] [PubMed]
- Papayiannis, L.C.; Katis, N.I.; Idris, A.M.; Brown, J.K. Identification of Weed Hosts of Tomato Yellow Leaf Curl Virus in Cyprus. Plant Dis. 2011, 95, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Wernecke, M.E.; Roye, M.E.; McLaughlin, W.A.; Nakhla, M.K.; Maxwell, D.P. Identification of Tomato Yellow Leaf Curl Geminivirus (TYLCV-Is) in Tomatoes and Pepper in Jamaica. In Proceedings of the Biology and Molecular Epidemiology of Geminiviruses, Tucson, Arizona, 3–8 June 1995; p. 17. [Google Scholar]
- Chinnaraja, C.; Ramkissoon, A.; Ramsubhag, A.; Jayaraman, J. Molecular Characterization of Tomato Yellow Leaf Curl Virus Infecting Tomato (Lycopersicum esculentum Mill.) in Trinidad and Tobago, West Indies. Trop. Agric. 2018, 95, 19–28. [Google Scholar]
- Polston, J.E.; McGovern, R.J.; Brown, L.G. Introduction of Tomato Yellow Leaf Curl Virus in Florida and Implications for the Spread of this and Other Geminiviruses of Tomato. Plant Dis. 1999, 83, 984–988. [Google Scholar] [CrossRef]
- Lefeuvre, P.; Martin, D.P.; Harkins, G.; Lemey, P.; Gray, A.J.A.; Meredith, S.; Lakay, F.; Monjane, A.; Lett, J.-M.; Varsani, A.; et al. The Spread of Tomato Yellow Leaf Curl Virus from the Middle East to the World. PLoS Pathog. 2010, 6, e1001164. [Google Scholar] [CrossRef]
- Delatte, H.; Dalmon, A.; Rist, D.; Soustrade, I.; Wuster, G.; Lett, J.M.; Goldbach, R.W.; Peterschmitt, M.; Reynaud, B. Tomato Yellow Leaf Curl Virus can be Acquired and Transmitted by Bemisia tabaci (Gennadius) from Tomato Fruit. Plant Dis. 2003, 87, 1297–1300. [Google Scholar] [CrossRef]
- Lee, H.; Song, W.; Kwak, H.R.; Kim, J.D.; Park, J.; Auh, C.K.; Kim, D.H.; Lee, K.Y.; Lee, S.; Choi, H.S. Phylogenetic Analysis and Inflow Route of Tomato Yellow Leaf Curl Virus (TYLCV) and Bemisia tabaci in Korea. Mol. Cells 2010, 30, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Polston, J.E.; De Barro, P.; Boykin, L.M. Transmission Specificities of Plant Viruses with the Newly Identified Species of the Bemisia tabaci Species Complex. Pest Manag. Sci. 2014, 70, 1547–1552. [Google Scholar] [CrossRef] [PubMed]
- Marchant, W.G.; Mugerwa, H.; Gautam, S.; Al-Aqeel, H.; Polston, J.E.; Rennberger, G.; Srinivasan, R. Phylogenomic and Population Genetics Analyses of Extant Tomato Yellow Leaf Curl Virus Strains on a Global Scale. Front. Virol. 2023, 3, 1221156. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Riley, D.G.; Srinivasan, R. Integrated Management of Tomato Yellow Leaf Curl Virus and its Whitefly Vector in Tomato. J. Econ. Entomol. 2019, 112, 1526–1540. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, M.; Legg, J.P.; Wintermantel, W.M.; Polston, J.E. Management of Whitefly-Transmitted Viruses in Open-Field Production Systems. In Advances in Virus Research; Loebenstein, G., Katis, N., Eds.; Academic Press: New York, NY, USA, 2014; Volume 90, pp. 147–206. [Google Scholar]
- Lapidot, M.; Karniel, U.; Gelbart, D.; Fogel, D.; Evenor, D.; Kutsher, Y.; Makhbash, Z.; Nahon, S.; Shlomo, H.; Chen, L.; et al. A Novel Route Controlling Begomovirus Resistance by the Messenger RNA Surveillance Factor Pelota. PLoS Genet. 2015, 11, e1005538. [Google Scholar] [CrossRef] [PubMed]
- Caro, M.; Verlaan, M.G.; Julián, O.; Finkers, R.; Wolters, A.A.; Hutton, S.F.; Scott, J.W.; Kormelink, R.; Visser, R.G.F.; Díez, M.J.; et al. Assessing the Genetic Variation of Ty-1 and Ty-3 Alleles Conferring Resistance to Tomato Yellow Leaf Curl Virus in a Broad Tomato Germplasm. Mol. Breed. 2015, 35, 132. [Google Scholar] [CrossRef]
- Marchant, W.G.; Gautam, S.; Hutton, S.F.; Srinivasan, R. Tomato Yellow Leaf Curl Virus-Resistant and -Susceptible Tomato Genotypes Similarly Impact the Virus Population Genetics. Front. Plant Sci. 2020, 11, 599697. [Google Scholar] [CrossRef]
- Voorburg, C.M.; Yan, Z.; Bergua-Vidal, M.; Wolters, A.M.A.; Bai, Y.; Kormelink, R. Ty-1, a Universal Resistance Gene against Geminiviruses that is Compromised by Co-Replication of a Betasatellite. Mol. Plant Pathol. 2020, 21, 160–172. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database. 2021. Available online: https://www.fao.org/food-agriculture-statistics/en/Rome (accessed on 20 December 2023).
- Gibson, R.W.; Mpembe, I.; Alicai, T.; Carey, E.E.; Mwanga, R.O.M.; Seal, S.E.; Vetten, H.J. Symptoms, Aetiology and Serological Analysis of Sweet Potato Virus Disease in Uganda. Plant Pathol. 1998, 47, 95–102. [Google Scholar] [CrossRef]
- Karyeija, R.F.; Kreuze, J.F.; Gibson, R.W.; Valkonen, J.P.T. Synergistic Interactions of a Potyvirus and a Phloem-Limited Crinivirus in Sweet Potato Plants. Virology 2000, 269, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.A.C. Global Plant Virus Disease Pandemics and Epidemics. Plants 2021, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Gibson, R.W.; Aritua, V. The Perspective of Sweet Potato Chlorotic Stunt Virus in Sweet Potato Production in Africa: A Review. Afr. Crop Sci. J. 2002, 10, 281–310. [Google Scholar] [CrossRef]
- Tugume, A.K.; Mukasa, S.B.; Kalkkinen, N.; Valkonen, J.P. Recombination and Selection Pressure in the Ipomovirus Sweet Potato Mild Mottle Virus (Potyviridae) in Wild Species and Cultivated Sweet Potato in the Centre of Evolution in East Africa. J. Gen. Virol. 2010, 91, 1092–1108. [Google Scholar] [CrossRef] [PubMed]
- Tugume, A.K.; Amayo, R.; Weinheimer, I.; Mukasa, S.B.; Rubaihayo, P.R.; Valkonen, J.P. Genetic Variability and Evolutionary Implications of RNA Silencing Suppressor Genes in RNA1 of Sweet Potato Chlorotic Stunt Virus Isolates Infecting Sweet Potato and Related Wild Species. PLoS ONE 2013, 8, e81479. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Lain, S.; Garcia, J.A. Highlights and Prospects of Potyvirus Molecular Biology. J. Gen. Virol. 1992, 73, 1–16. [Google Scholar] [CrossRef]
- Dougherty, W.G.; Carrington, J.C. Expression and Function of Potyviral Gene Products. Annu. Rev. Phytopathol. 1988, 26, 123. [Google Scholar] [CrossRef]
- Kreuze, J.; Fuentes, S. Sweet Potato Viruses. In Encyclopedia of Virology; Mahy, B.W.J., Van Regenmortel, M.H.V., Eds.; Academic Press: Amsterdam, The Netherlands, 2009; pp. 659–669. [Google Scholar]
- Kreuze, J.F.; Savenkov, E.I.; Valkonen, J.P.T. Complete Genome Sequence and Analyses of the Subgenomic RNAs of Sweet Potato Chlorotic Stunt Virus Reveal Several New Features for the Genus Crinivirus. J. Virol. 2002, 76, 9260–9270. [Google Scholar] [CrossRef] [PubMed]
- Dolja, V.V.; Kreuze, J.F.; Valkonen, J.P. Comparative and Functional Genomics of Closteroviruses. Virus Res. 2006, 117, 38–51. [Google Scholar] [CrossRef]
- Tairo, F.; Mukasa, S.B.; Jones, R.A.C.; Kullaya, A.; Rubaihayo, P.R.; Valkonen, J.P.T. Unravelling the Genetic Diversity of the Three Main Viruses Involved in Sweet Potato Virus Disease (SPVD), and its Practical Implications. Mol. Plant Pathol. 2005, 6, 199–211. [Google Scholar] [CrossRef]
- Ishak, J.A.; Kreuze, J.F.; Johansson, A.; Mukasa, S.B.; Tairo, F.; Abo El-Abbas, F.M.; Valkonen, J.P.T. Some Molecular Characteristics of Three Viruses from Sweet Potato virus Disease (SPVD)-Affected Sweet Potato Plants in Egypt. Arch. Virol. 2003, 148, 2449–2460. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.; Loebenstein, G. Role of a Whitefly-Transmitted Agent in Infection of Sweet Potato by Cucumber Mosaic Virus. Plant Dis. 1991, 75, 291–292. [Google Scholar] [CrossRef]
- Cuellar, W.J.; Galvez, M.; Fuentes, S.; Tugume, J.; Kreuze, J. Synergistic Interactions of Begomoviruses with Sweet Potato Chlorotic Stunt Virus (Genus Crinivirus) in Sweet Potato (Ipomoea batatas L.). Mol. Plant Pathol. 2015, 16, 459–471. [Google Scholar] [CrossRef]
- Lopez, L.; Alleyne, A.T. Key Operational and Technical Gaps for the Provision of Clean Sweet Potato Planting Material in Barbados; FAO Project TCP/BAR/3701: Activity #4 Report; Food and Agriculture Organization: Rome, Italy, 2020; p. 26. [Google Scholar]
- Alleyne, A.T. Structural and Functional Analysis of a Resistance Gene and Resistance Gene Analogs in Local Cultivars of Ipomoea batatas Lam. from Barbados. Physiol. Mol. Plant Pathol. 2022, 120, 101855. [Google Scholar] [CrossRef]
- Gadhave, K.R.; Gautam, S.; Rasmussen, D.A.; Srinivasan, R. Aphid Transmission of Potyvirus: The Largest Plant-Infecting RNA Virus Genus. Viruses 2020, 12, 773. [Google Scholar] [CrossRef]
- Holmes, E.C. The Evolution and Emergence of RNA Viruses; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Nigam, D.; LaTourrette, K.; Souza, P.F.; Garcia-Ruiz, H. Genome-Wide Variation in Potyviruses. Front. Plant Sci. 2019, 10, 1439. [Google Scholar] [CrossRef]
- Chinnadurai, C.; Kollam, M.; Ramsubhag, A.; Jayaraman, J. Genome Characterization of Zucchini Yellow Mosaic Virus Infecting Cucurbits Reveals the Presence of a New Genotype in Trinidad and Tobago in the Caribbean Region. Arch. Virol. 2021, 166, 1661–1669. [Google Scholar] [CrossRef]
- Gal-on, A. Zucchini Yellow Mosaic Virus: Insect Transmission and Pathogenicity—The Tails of Two Proteins. Mol. Plant Pathol. 2007, 8, 139–150. [Google Scholar] [CrossRef]
- Provvidenti, R.; Gonsalves, D.; Humaydan, H.S. Occurrence of Zucchini Yellow Mosaic virus in Cucurbits from Connecticut, New York, Florida and California. Plant Dis. 1984, 68, 443–446. [Google Scholar] [CrossRef]
- Coutts, B.A.; Kehoe, M.A.; Jones, R.A.C. Minimising Losses caused by Zucchini Yellow Mosaic Virus in Vegetable Cucurbit Crops in Tropical, Sub-Tropical and Mediterranean Environments Through Cultural Methods and Host Resistance. Virus Res. 2011, 159, 141–160. [Google Scholar] [CrossRef]
- Cambra, M.; Vidal, E. A Vector-borne Disease Caused by Plum Pox Virus: Vector Species, Transmission Mechanism, Epidemiology and Mitigation Strategies to Reduce its Natural Spread. Acta Hortic. 2017, 1163, 57–68. [Google Scholar] [CrossRef]
- Stapleton, J.J.; Summers, C.G. Reflective Mulches for Management of Aphids and Aphid-borne Virus Diseases in Late-Season Cantaloupe (Cucumis melo L. var. cantalupensis). Crop Prot. 2002, 21, 891–898. [Google Scholar] [CrossRef]
- Hooks, C.R.; Fereres, A. Protecting Crops from Non-persistently Aphid-transmitted Viruses: A Review on the Use of Barrier Plants as a Management Tool. Virus Res. 2006, 120, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Boquel, S.; Giguère, M.A.; Pelletier, Y. Effect of Mineral Oils on Host Plant Selection and Probing Behavior of Rhopalosiphum Padi. Entomol. Exp. Appl. 2016, 160, 241–250. [Google Scholar] [CrossRef]
- Dupuis, B. The Movement of Potato Virus Y (PVY) in the Vascular System of Potato Plants. Eur. J. Plant. Pathol. 2017, 147, 365–373. [Google Scholar] [CrossRef]
- Desbiez, C.; Wipf-Scheibel, C.; Lecoq, H. Biological and Serological Variability, Evolution and Molecular Epidemiology of Zucchini Yellow Mosaic Virus (ZYMV, Potyvirus) with Special Reference to Caribbean Islands. Vir. Res. 2002, 85, 5–16. [Google Scholar] [CrossRef]
- Lecoq, H.; Wipf-Scheibel, C.; Chandeysson, C.; Lê Van, A.; Fabre, F.; Desbiez, C. Molecular Epidemiology of Zucchini Yellow Mosaic Virus in France: An Historical Overview. Virus Res. 2009, 141, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, M.; Bar-Joseph, M.; Candresse, T.; Maree, H.J.; Martelli, G.P.; Melzer, M.J.; Menzel, W.; Minafra, A.; Sabanadzovic, S.; Report Consortium, I. ICTV Virus Taxonomy Profile: Closteroviridae. J. Gen. Virol. 2020, 101, 364–365. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.; Duffus, J. A Review of a Yellowing and Stunting Disorder of Cucurbits in the United Arab Emirates. Emir. J. Food Agric. 1990, 2, 1–16. [Google Scholar] [CrossRef]
- Sinclair, J.W.; Crosby, K.M. A Review of Cucurbit Yellow Stunting Disorder Virus (CYSDV)—A “New” Virus Affecting Melons in the Lower Rio Grande Valley. Subtrop. Plant Sci. 2002, 54, 54–58. [Google Scholar]
- Wintermantel, W.M.; Hladky, L.L.; Cortez, A.A.; Natwick, E.T. A New Expanded Host Range of Cucurbit Yellow Stunting Disorder Virus Includes Three Agricultural Crops. Plant Dis. 2009, 93, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Janssen, D. Cucurbit Yellow Stunting Disorder. Available online: https://www.cabidigitallibrary.org/doi/10.1079/cabicompendium.17070 (accessed on 9 January 2024).
- Tzanetakis, I.E.; Martin, R.R.; Wintermantel, W.M. Epidemiology of Criniviruses: An Emerging Problem in World Agriculture. Front. Microbiol. 2013, 4, 119. [Google Scholar] [CrossRef] [PubMed]
- Celix, A.; Lopez-Sese, A.; Almarza, N.; Gomez-Guillamon, M.L.; Rodriguez-Cerezo, E. Characterization of Cucurbit Yellow Stunting Disorder Virus, a Bemisia tabaci-Transmitted Closterovirus. Phytopathology 1996, 86, 1370–1376. [Google Scholar]
- Jones, R.A. Plant Virus Emergence and Evolution: Origins, New Encounter Scenarios, Factors Driving Emergence, Effects of Changing World Conditions, and Prospects for Control. Virus Res. 2009, 141, 113–130. [Google Scholar] [CrossRef] [PubMed]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global Biodiversity Conservation: The Critical Role of Hotspots. In Biodiversity Hotspots: Distribution and Protection of Conservation Priority Areas; Zachos, F., Habel, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 3–22. [Google Scholar]
- Turner, M.P. Rugose Mosaic on Red Pontiac Potato. Food Agric. Organ. United Nations Plant Prot. Bull. 1978, 26, 131–132. [Google Scholar]
- Hallman, G. Phytosanitary Measures to Prevent the Introduction of Invasive Species. In Biological Invasions, Ecological Studies; Nentwig, W., Ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 2007; Volume 193, pp. 367–384. [Google Scholar]
- Khoury, W.E.; Makkouk, K. Integrated Plant Disease Management in Developing Countries. J. Plant Pathol. 2010, 92, S35–S42. [Google Scholar]
- Saravanakumar, D.; Bartholomew, E.S.; Seepersad, G.; Gore-Francis, J.; Goldsmith, J.; Ramnanan, N.; Chang, P.G.; Bridgemohan, P.; Sewsaran, R.; Medrano-Cabral, S.; et al. Prioritisation of quarantine pest list for the Caribbean using a multi-criteria decision approach. NeoBiota 2023, 88, 1–16. [Google Scholar] [CrossRef]
- Van der Graaff, N.A.; Khoury, W. Biosecurity in the Movement of Commodities as a Component of Global Food Security. In The Role of Plant Pathology in Food Safety and Food Security; Strange, R.N., Gullido, M.L., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 25–39. [Google Scholar]
- Nelson, R. International Plant Pathology: Past and Future Contributions to Global Food Security. Phytopathology 2020, 110, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Elena, S.F.; Agudelo-Romero, P.; Lalić, J. The Evolution of Viruses in Multi-host Fitness Landscapes. Open Virol. J. 2009, 3, 1. [Google Scholar] [CrossRef]
- Alexander, H.M.; Mauck, K.E.; Whitfield, A.E.; Garrett, K.A.; Malmstrom, C.M. Plant-Virus Interactions and the Agro-Ecological Interface. Eur. J. Plant Pathol. 2014, 138, 529–547. [Google Scholar] [CrossRef]
- Syller, J. Biological and Molecular Events Associated with Simultaneous Transmission of Plant Viruses by Invertebrate and Fungal Vectors. Mol. Plant Pathol. 2014, 15, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Fraile, A.; García-Arenal, F. Environment and Evolution Modulate Plant Virus Pathogenesis. Curr Opin Virol. 2016, 17, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.R.; James, D.; Lévesque, C.A. Impacts of Molecular Diagnostic Technologies on Plant Disease Management. Ann. Rev. Phytopathol. 2000, 38, 207–239. [Google Scholar] [CrossRef]
- Hema, M.; Konakalla, N.C. Recent Developments in Detection and Diagnosis of Plant Viruses. In Recent Developments in Applied Microbiology and Biochemistry; Buddolla, V., Ed.; Elsevier Science: Amsterdam, The Netherlands, 2021; Volume 2, pp. 163–180. [Google Scholar]
- Lecoq, H.; Desbiez, C. Viruses of Cucurbit Crops in the Mediterranean Region: An Ever-changing Picture. Adv. Virus Res. 2012, 84, 67–126. [Google Scholar] [PubMed]
- Bhat, A.I.; Aman, R.; Mahfouz, M.M. Onsite Detection of Plant Viruses using Isothermal Amplification Assays. Plant Biotechnol. J. 2022, 20, 1859–1873. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.; Lockhart, C.; Pongolini, S.; Karesh, W.B.; Baylis, M.; Goldberg, T.; Slingenbergh, J.; Gale, P.; Venturini, T.; Catchpole, M.; et al. Drivers for Emerging Issues in Animal and Plant Health. EFSA J. 2016, 14, e00512. [Google Scholar] [CrossRef] [PubMed]
- Cooper, I.; Jones, R.A.C. Wild Plants and Viruses: Under Investigated Ecosystems. Adv. Virus Res. 2006, 67, 1–47. [Google Scholar]
- Roossinck, M.J.; García-Arenal, F. Ecosystem Simplification, Biodiversity Loss and Plant Virus Emergence. Curr. Opin. Virol. 2015, 10, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Prajapat, R.; Marwal, A.; Gaur, R.K. Begomovirus Associated with Alternative Host Weeds: A Critical Appraisal. Arch. Phytopathol. Plant Prot. 2014, 47, 157–170. [Google Scholar] [CrossRef]
- García-Arenal, F.; Zerbini, F.M. Life on the Edge: Geminiviruses at the Interface between Crops and Wild Plant Hosts. Ann. Rev.Virol. 2019, 6, 411–433. [Google Scholar] [CrossRef]
- Hill, J.E.; Strandberg, J.O.; Hiebert, E.; Lazarowitz, S.G. Asymmetric Infectivity of Pseudorecombinants of Cabbage Leaf Curl Virus and Squash Leaf Curl Virus: Implications for Bipartite Geminivirus Evolution and Movement. Virology 1998, 250, 283–292. [Google Scholar] [CrossRef]
- Abouzid, A.M.; Hiebert, E.; Strandbery, J.O. Cloning Identification, and Partial Sequencing of the Genomic Components of a Geminivirus Infecting the Brassicaceae. Phytophathology 1992, 82, 1070. [Google Scholar]
- Rampersad, S.N.; Umaharan, P. Detection of Two Bipartite Geminiviruses Infecting Dicotyledonous Weeds in Trinidad. Plant Dis. 2003, 87, 602. [Google Scholar] [CrossRef] [PubMed]
- McLeish, M.J.; Fraile, A.; García-Arenal, F. Population Genomics of Plant Viruses: The Ecology and Evolution of Virus Emergence. Phytopathology 2021, 111, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.A.; Naidu, R.A. Global Dimensions of Plant Virus Diseases: Current Status and Future Perspectives. Ann. Rev. Virol. 2019, 6, 387–409. [Google Scholar] [CrossRef] [PubMed]
- Stansly, P.; Natwick, E. Integrated Systems for Managing Bemisia tabaci in Protected and Open Field Agriculture. In Bemisia: Bionomics and Management of a Global Pest; Stansly, P.A., Naranjo, S.E., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 467–497. [Google Scholar]
- Gilbertson, R.L.; Rojas, M.R.; Kon, T.; Jaquez, J. Introduction of Tomato Yellow Leaf Curl Virus into the Dominican Republic: The Development of a Successful Integrated Pest Management Strategy. In Tomato Yellow Leaf Curl Virus Disease: Management, Molecular Biology, Breeding for Resistance; Czosnek, H., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 279–303. [Google Scholar]
- Ganpat, W.; Harder, A.; Moore, A. Envisioning the Future of Extension and Advisory Services in the Caribbean. J. Int. Agric. Ext. Educ. 2014, 21, 19–31. [Google Scholar] [CrossRef]
- Gibson, R.W.; Kreuze, J.F. Degeneration in Sweet Potato Due to Viruses, Virus-cleaned Planting Material and Reversion: A Review. Plant Pathol. 2015, 64, 1–15. [Google Scholar] [CrossRef]
- Perring, T.M.; Cooper, A.D.; Rodriguez, R.J.; Farrar, C.A.; Bellows, T.S., Jr. Identification of a Whitefly Species by Genomic and Behavioral Studies. Science 1993, 259, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Naveen, N.C.; Chaubey, R.; Kumar, D.; Rebijith, K.B.; Rajagopal, R.; Subrahmanyam, B.; Subramanian, S. Insecticide Resistance Status in the Whitefly, Bemisia tabaci Genetic Groups Asia-I, Asia-II-1 and Asia-II-7 on the Indian Subcontinent. Sci. Rep. 2017, 7, 40634. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.Z.; Ahmed, S.; Al-Ashkar, I.; El Sabagh, A.; Liu, L.; Zhong, G. Evaluation of Resistance Development in Bemisia tabaci Genn. (Homoptera: Aleyrodidae) in Cotton against Different Insecticides. Insects 2021, 12, 996. [Google Scholar] [CrossRef]
- Rajna, S.; Praveen, K.; Laneesha, M.; Kelageri, S.S. Recent Trends in Insecticide Resistance Research on Whiteflies (Hemiptera: Aleyrodidae): A Bibliometric Profile. Curr. Sci. 2021, 120, 1433–1440. [Google Scholar] [CrossRef]
- Rampersad, S.N. Proposed Strategies for Begomovirus Disease Management in Tomato in Trinidad. Plant Health Prog. 2003, 4, 14. [Google Scholar] [CrossRef]
- Wang, A.; Krishnaswamy, S. Eukaryotic Translation Initiation Factor 4E-Mediated Recessive Resistance to Plant Viruses and its Utility in Crop Improvement. Mol. Plant Pathol. 2012, 13, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Kis, A.; Hamar, É.; Tholt, G.; Bán, R.; Havelda, Z. Creating Highly Efficient Resistance against Wheat Dwarf Virus in Barley by Employing CRISPR/Cas9 System. Plant Biotechnol. J. 2019, 17, 1004. [Google Scholar] [CrossRef] [PubMed]
- Schmitt-Keichinger, C. Manipulating Cellular Factors to Combat Viruses: A Case Study from the Plant Eukaryotic Translation Initiation Factors eIF4. Front. Microbiol. 2019, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Collinge, D.B.; Sarrocco, S. Transgenic Approaches for Plant Disease Control: Status and Prospects 2021. Plant Pathol. 2022, 71, 207–225. [Google Scholar] [CrossRef]
- Kumar, K.; Gambhir, G.; Dass, A.; Tripathi, A.K.; Singh, A.; Jha, A.K.; Yadava, P.; Choudhary, M.; Rakshit, S. Genetically Modified Crops: Current Status and Future Prospects. Planta 2020, 251, 91. [Google Scholar] [CrossRef] [PubMed]
- Rosado, A.; Eriksson, D. Biosafety Legislation and the Regulatory Status of the Products of Precision Breeding in the Latin America and the Caribbean Region. Plants People Planet 2022, 4, 214–231. [Google Scholar] [CrossRef]
- Aregbesola, O.Z.; Legg, J.P.; Sigsgaard, L.; Lund, O.S.; Rapisarda, C. Potential Impact of Climate Change on Whiteflies and Implications for the Spread of Vectored Viruses. J. Pest Sci. 2019, 92, 381–392. [Google Scholar] [CrossRef]
- Douglass, K.; Cooper, J. Archaeology, Environmental Justice, and Climate Change on Islands of the Caribbean and Southwestern Indian Ocean. Proc. Natl. Acad. Sci. USA 2020, 117, 8254–8262. [Google Scholar] [CrossRef]
- Lincoln Lenderking, H.; Robinson, S.A.; Carlson, G. Climate Change and Food Security in Caribbean Small Island Developing States: Challenges and Strategies. Int. J. Sustain. Dev. World Ecol. 2021, 28, 238–245. [Google Scholar] [CrossRef]
- Dash, S.P.; Dipankar, P.; Burange, P.S.; Rouse, B.T.; Sarangi, P.P. Climate Change: How it Impacts the Emergence, Transmission, Resistance and Consequences of Viral Infections in Animals and Plants. Crit. Rev. Microbiol. 2021, 47, 307–322. [Google Scholar] [CrossRef] [PubMed]
- Altieri, M.A.; Nicholls, C.I.; Henao, A.; Lana, M.A. Agroecology and the Design of Climate Change-Resilient Farming Systems. Agron. Sustain. Dev. 2015, 35, 869–890. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Virus | Common Host Name | Host Species | Reported Location | Year of Detection | Method of Detection a | References |
|---|---|---|---|---|---|---|---|
| Endemic and potential re-emerging threats | |||||||
| Begomovirus | Bean golden mosaic virus | Red kidney bean | Phaseolus vulgaris | Jamaica | 1975, 1994, 1996 | Symptomatology, PCR, Sanger sequencing | [31,32,33] |
| Badnavirus | Cacao mild mosaic virus | Cacao | Theobroma cacao | Trinidad | 1944, 1947, 2005 | Symptomatology, HTS | [34,35,36] |
| Cacao yellow vein-banding virus | Cacao | Theobroma cacao | Trinidad | 1944, 1947, 2005 | Symptomatology, HTS | [34,35,36] | |
| Closterovirus | Citrus tristeza virus | Citrus | Citrus spp. | Jamaica, Trinidad | 1960, 1965, 2002, 2009, 2010, 2013 | Bio-indexing, ELISA, RT-PCR, Sanger sequencing | [37,38,39,40,41,42] |
| Cucumovirus | Cucumber mosaic virus | Tomato, hot pepper, sweet potato | Solanum lycopersicum, Capsicum chinense, Ipomoea batatas | Jamaica, Trinidad | 1974, 1992, 2004, 2011 | Symptomology, NCM-ELISA | [43] |
| Potyvirus | Papaya ringspot virus | Papaya | Carica papaya | Jamaica, Trinidad | 1929, 2014 | Symptomatology, ELISA, RT-PCR, Sanger sequencing | [44,45] |
| Potato virus Y | Sweet pepper, scotch bonnet pepper, bird pepper, tobacco, tomato | Capsicum annuum, Capsicum chinense, Capsicum baccatum, Nicotiana tabacum, Solanum lycopersicum | Jamaica | 1950–1951, 1961, 1976, 1979, 1996, 2010 | Symptomatology; Biological indicators, transmission tests, ELISA | [46,47,48,49,50,51] | |
| Tobamovirus | Tobacco mosaic virus | Tomato, hot pepper | Solanum lycopersicum, Capsicum chinense | Trinidad | 1974, 1992, 2006 | Symptomology, ELISA | [43] |
| Recent and ongoing threats | |||||||
| Begomovirus | Cabbage leaf curl virus | Cabbage | Brassica oleracea | Jamaica | 1990s | PCR & Sanger sequencing | [52,53] |
| Pepper huasteco yellow vein virus | Sweet pepper | Capsicum annuum | Trinidad | 1995 | Hybridization, PCR, Sanger sequencing | [54] | |
| Potato yellow mosaic virus | Tomato, sweet pepper, wild chili pepper, ochro, red kidney bean | Solanum lycopersicum, Capsicum annuum, Capsicum frutescens, Abelmoschus esculentus, Phaseolus vulgaris | Trinidad | 1995 | Hybridization, PCR, Sanger sequencing | [54] | |
| Tomato mottle virus | Tomato | Solanum lycopersicum | Trinidad | 1998 | Dot blot hybridization, PCR | [43] | |
| Tomato mosaic Havana virus | Tomato, scotch bonnet pepper | Solanum lycopersicum, Capsicum chinense | Jamaica | 1994, 1996 | PCR, RFLP, Sanger sequencing | [55,56] | |
| Tomato yellow leaf curl virus | Tomato, sweet pepper, wild chili pepper, ochro, red kidney bean, cowpea | Solanum lycopersicum, Capsicum annuum, Capsicum frutescens, Abelmoschus esculentus, Phaseolus vulgaris, Vigna unguiculata | Barbados, Jamaica, Trinidad | 1993, 1994, 1995, 1998, 2014–2016 | Hybridization, PCR, RFLP, Sanger sequencing | [54,55,56,57,58,59] | |
| Tomato yellow mosaic virus | Sweet pepper | Capsicum annuum | Trinidad | 1995 | Hybridization, PCR, Sanger sequencing | [54] | |
| Sweet potato leaf curl virus | Sweet potato | Ipomoea batatas | Barbados, Jamaica | 2004, 2013, 2017, 2018 | RT-PCR, Sanger sequencing, RCA, HTS | [60,61,62,63] | |
| Crinivirus | Sweet potato chlorotic stunt virus | Sweet potato | Ipomoea batatas | Barbados, Jamaica | 2001, 2013–2014 | NCM-ELISA, RT-PCR, HTS | [63,64,65] |
| Ipomovirus | Sweet potato mild mottle virus | Sweet potato | Ipomoea batatas | Jamaica | 2011 | NCM-ELISA | [60,61] |
| Potyvirus | Ipomoea vein mosaic virus (Sweet potato virus 2) | Sweet potato | Ipomoea batatas | Barbados | 2001 | NCM-ELISA | [65] |
| Sweet potato feathery mottle virus | Sweet potato | Ipomoea batatas | Barbados, Jamaica, | 2001, 2013–2014 | NCM-ELISA, RT-PCR, HTS | [63,64,65] | |
| Sweet potato latent virus | Sweet potato | Ipomoea batatas | Jamaica | 2011 | NCM-ELISA | [60,61] | |
| Sweet potato mild speckling virus | Sweet potato | Ipomoea batatas | Jamaica | 2006, 2011 | NCM-ELISA | [60,64] | |
| Sweet potato virus G | Sweet potato | Ipomoea batatas | Barbados, Jamaica | 2001, 2006, 2013 | NCM-ELISA, HTS | [61,64,65] | |
| Tobacco etch virus | Sweet pepper, scotch bonnet pepper, tomato | Capsicum annuum, Capsicum chinense, Solanum lycopersicum | Barbados, Jamaica, Trinidad | Early to mid-1990s, 2006 | Symptomatology, biological indicators, transmission tests, ELISA, RT-PCR, Sanger sequencing | [43,50,51] | |
| Watermelon mosaic virus/papaya ringspot virus—watermelon type | Cantaloupe, cucumber, pumpkin, watermelon | Cucumis melo, Cucumis sativa, Cucurbita pepo, Citrullus lanatus | Jamaica, Trinidad | 1986, 2020 | Symptomatology, biological indicators transmission tests | [43,66] | |
| Newly recognized threats | |||||||
| Begomovirus | Ipomoea yellow vein virus | Sweet potato | Ipomoea batatas | Barbados | 2018 | RT-PCR, HTS | [65] |
| Comovirus | Squash mosaic virus (Comovirus curcubitae) | Pumpkin | Cucurbita spp. | Jamaica, Trinidad | 2014, 2015 | ELISA, RT-PCR | [67,68] |
| Crinivirus | Cucurbit yellow stunting disorder virus | Cantaloupe, cucumber, watermelon | Cucumis melo, Cucumis sativa, Citrullus lanatus | Jamaica | 2019, 2020 | RT-PCR | [69] |
| Potyvirus | Yam mild mosaic virus | Yam | Dioscorea spp. | Barbados | 2018 | RT-PCR | [43] |
| Zucchini yellow mosaic virus | Pumpkin | Cucurbita spp. | Jamaica, Trinidad | 2014, 2016 | Symptomology, RT-PCR, Sanger sequencing | [67,68] | |
| Genus | Virus | Common Host Name | Host Species | Reported Location | Year of Detection | References |
|---|---|---|---|---|---|---|
| Alphanucleorhabdovirus | Maize mosaic virus (Alphanucleorhabdovirus maydis) | Corn | Zea mays | Trinidad | 1933 | [43] |
| Caulimovirus | Cauliflower mosaic virus | Crucifers | Brassica spp. | Trinidad | 1948 | [43] |
| Comovirus | Cowpea severe mosaic virus | Cowpea | Vigna unguiculata | Trinidad | 1962, 2007 | [43] |
| Polerovirus | Potato leafroll virus | Tomato | Solanum lycopersicum | Trinidad | 1974 | [43] |
| Potexvirus | Cassava common mosaic virus | Cassava | Manihot esculenta | Trinidad | 1989 | [46] |
| Potyvirus | Bean common mosaic virus | Red kidney bean | Phaseolus vulgaris | Barbados, Jamaica | 1990s | [46] |
| Dasheen mosaic virus | Dasheen | Colocasia esculenta | Trinidad | 2014 | [43] | |
| Johnsongrass mosaic virus | Sugarcane | Saccharum officinarum | Jamaica | 1919 | [46] | |
| Maize dwarf mosaic virus | Sugarcane | Saccharum officinarum | Jamaica | 1919 | [46] | |
| Pepper vein banding virus | Sweet pepper | Capsicum annuum | Jamaica | 1960 | [46] | |
| Potato virus Y in rugose mosaic | Irish potato | Solanum tuberosum | Jamaica | 1974 | [193] | |
| Sorghum mosaic virus | Sugarcane | Saccharum officinarum | Jamaica | 1919 | [46] | |
| Soybean mosaic virus | Soybean | Glycine max | Jamaica | 1940 | [46] | |
| Sugarcane mosaic virus | Sugarcane, corn | Saccharum officinarum, Zea mays | Jamaica, Trinidad | 1919, 1942, 1989, 2022 | [43,46] | |
| Yam mosaic virus | Yam | Dioscorea spp. | Jamaica, Trinidad | 1963, 1979 | [43,46] | |
| Tobamovirus | Pepper mild mottle virus | Scotch bonnet pepper | Capsicum chinense | Trinidad | 2014 | [40] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tennant, P.; Rampersad, S.; Alleyne, A.; Johnson, L.; Tai, D.; Amarakoon, I.; Roye, M.; Pitter, P.; Chang, P.-G.; Myers Morgan, L. Viral Threats to Fruit and Vegetable Crops in the Caribbean. Viruses 2024, 16, 603. https://doi.org/10.3390/v16040603
Tennant P, Rampersad S, Alleyne A, Johnson L, Tai D, Amarakoon I, Roye M, Pitter P, Chang P-G, Myers Morgan L. Viral Threats to Fruit and Vegetable Crops in the Caribbean. Viruses. 2024; 16(4):603. https://doi.org/10.3390/v16040603
Chicago/Turabian StyleTennant, Paula, Sephra Rampersad, Angela Alleyne, Lloyd Johnson, Deiondra Tai, Icolyn Amarakoon, Marcia Roye, Patrice Pitter, Peta-Gaye Chang, and Lisa Myers Morgan. 2024. "Viral Threats to Fruit and Vegetable Crops in the Caribbean" Viruses 16, no. 4: 603. https://doi.org/10.3390/v16040603
APA StyleTennant, P., Rampersad, S., Alleyne, A., Johnson, L., Tai, D., Amarakoon, I., Roye, M., Pitter, P., Chang, P.-G., & Myers Morgan, L. (2024). Viral Threats to Fruit and Vegetable Crops in the Caribbean. Viruses, 16(4), 603. https://doi.org/10.3390/v16040603