Activation of LTRs from Different Human Endogenous Retrovirus (HERV) Families by the HTLV-1 Tax Protein and T-Cell Activators

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
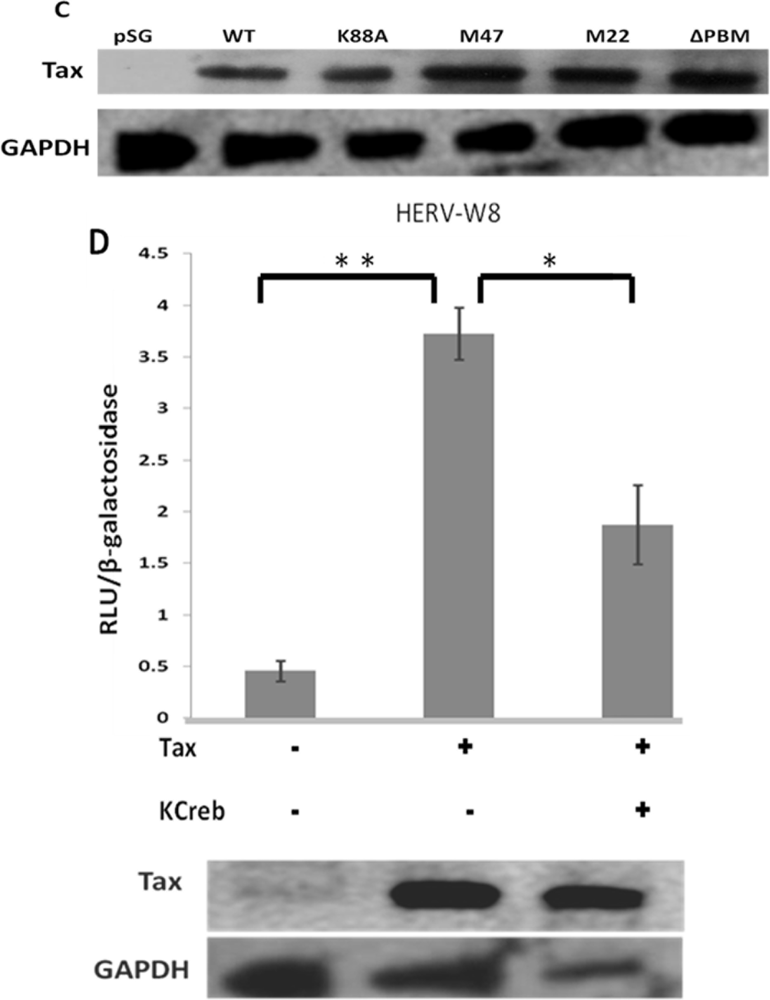
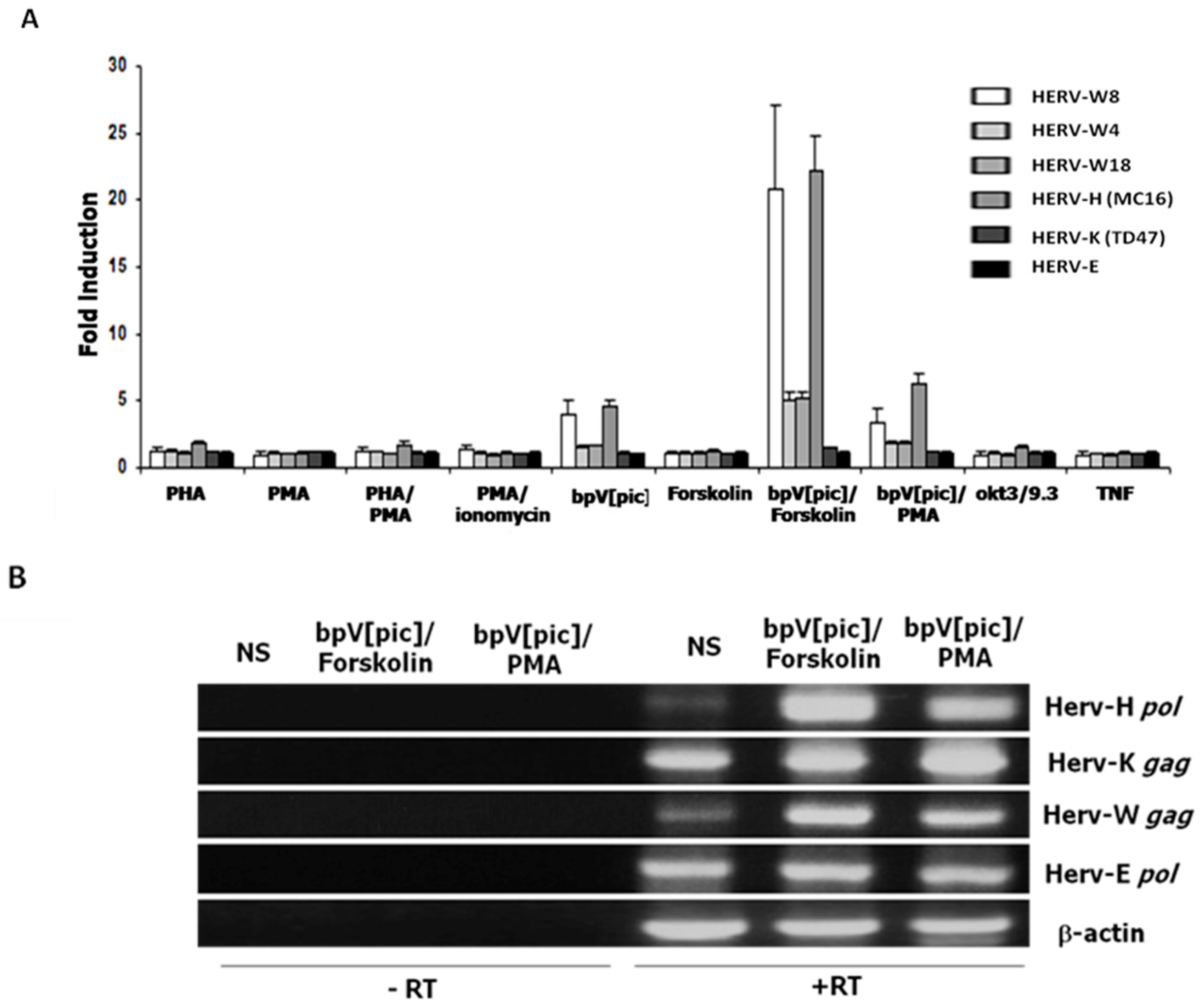
2.1. Different HERV LTRs Are Activated upon T Cell Activation
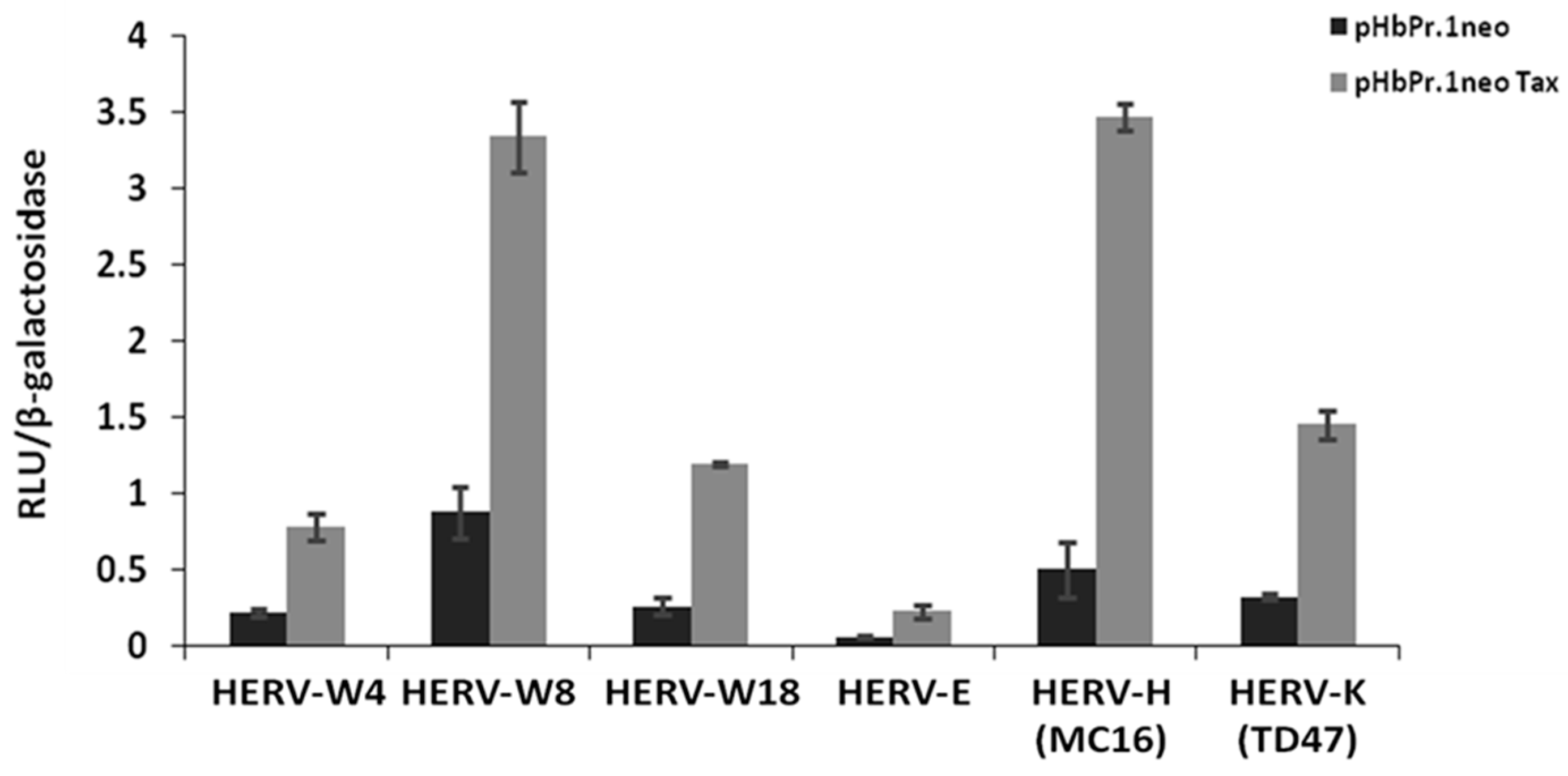
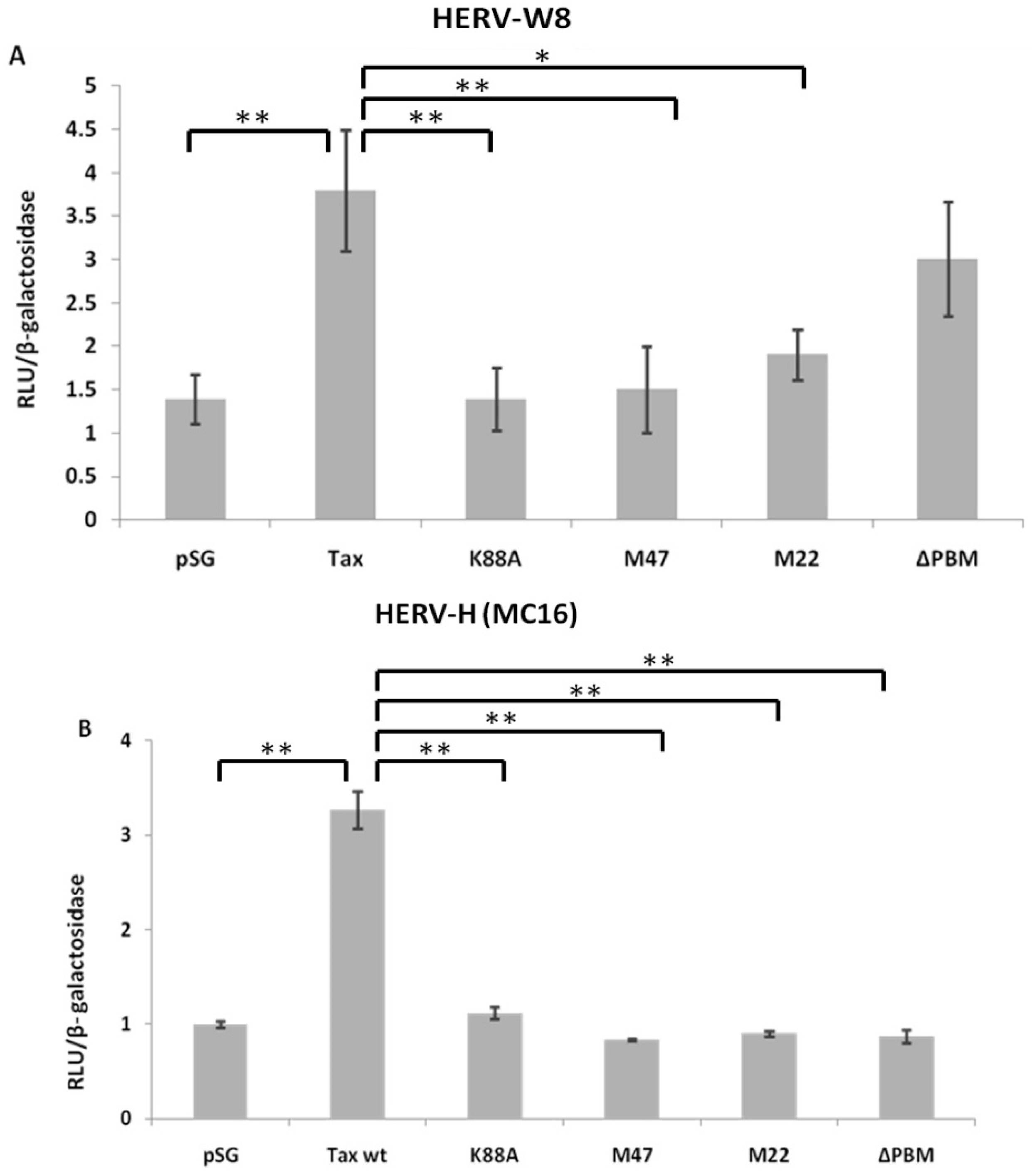
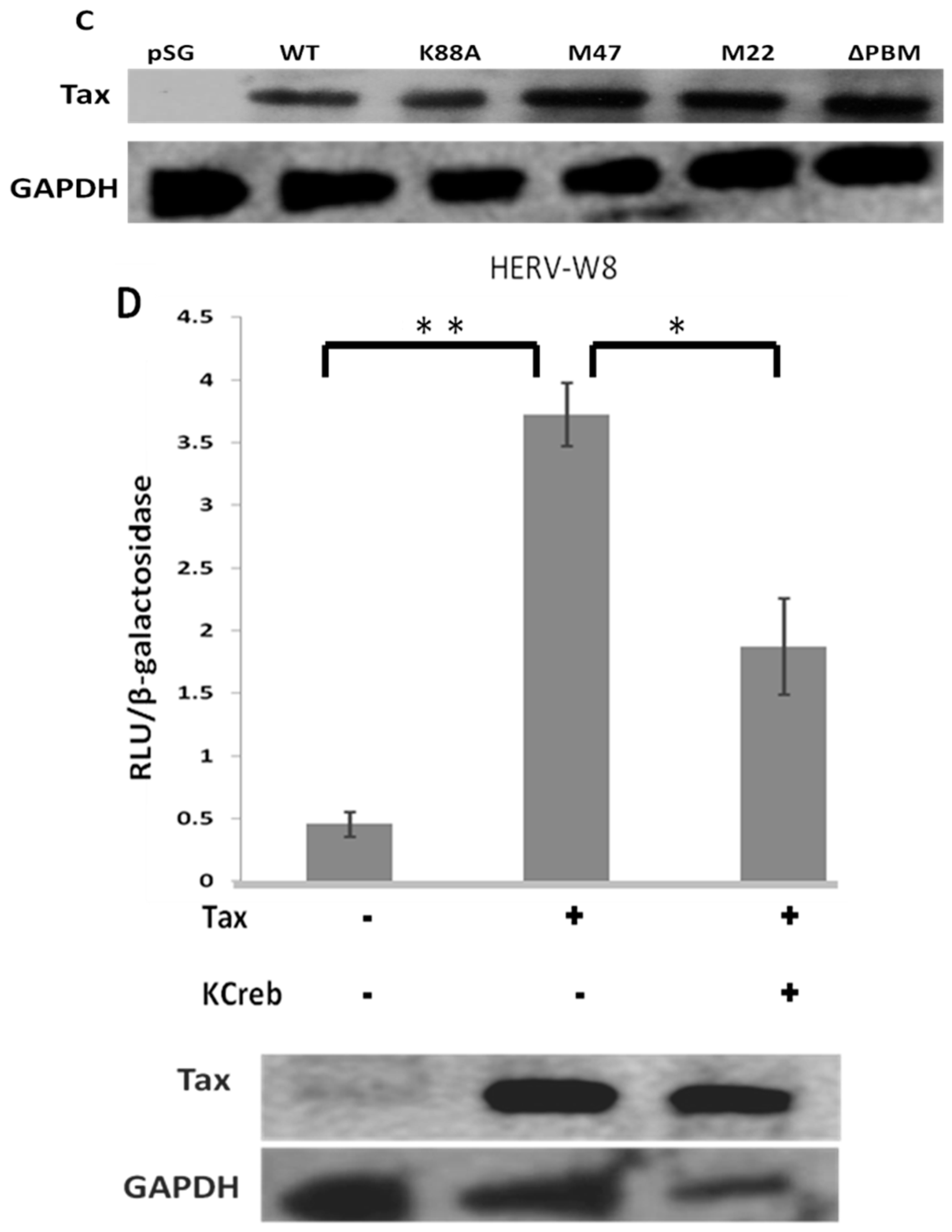
2.2. HTLV-1 Tax Activates Different HERV LTRs
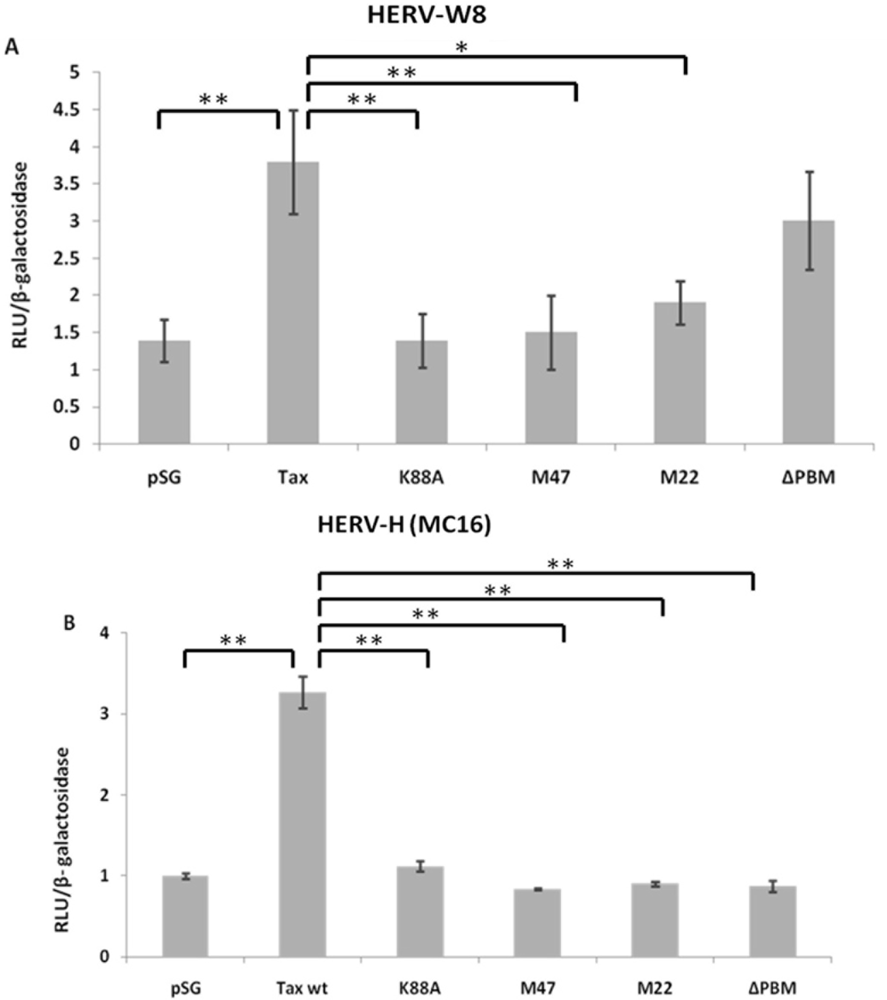
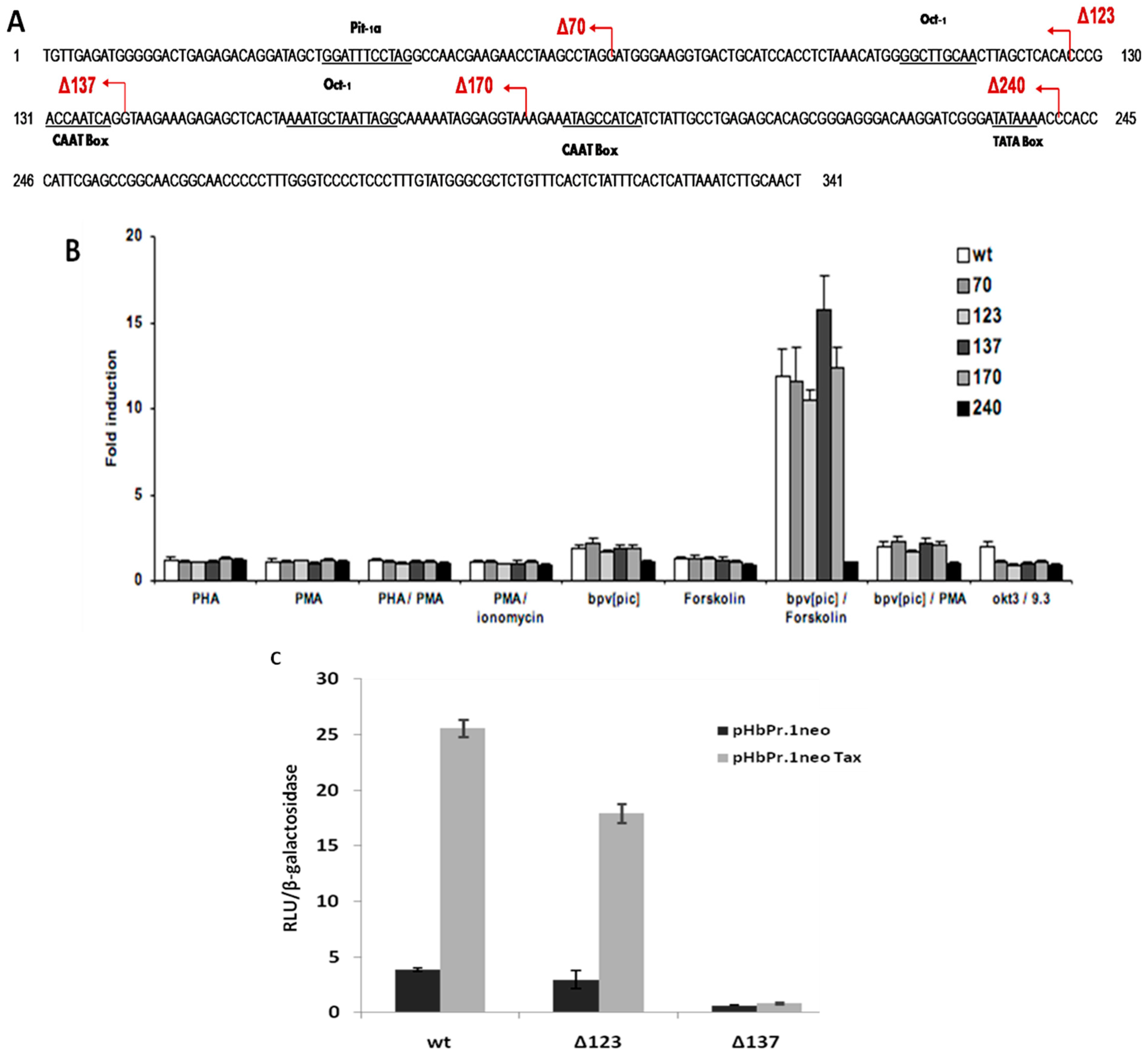
2.3. HERV-W8 LTR-Responsive Regions to Tax and T-Cell Activators Are Different
3. Experimental Section
3.1. Plasmids
3.2. Generation of Deletions Mutants by Exonuclease III
3.3. Transfection and Assay for Luciferase Activity
3.4. Total RNA Extraction and Semi-Quantitative RT-PCR
3.5. Western Blot Analyses
3.6. Statistical Analyses
4. Conclusions
Acknowledgments
References and Notes
- Griffiths, D.J. Endogenous retroviruses in the human genome sequence. Genome Biol. 2001, 2, 1017. [Google Scholar] [CrossRef] [PubMed]
- Lefranc, D.; Dubucquoi, S.; Almeras, L.; De Seze, J.; Tourvieille, B.; Dussart, P.; Aubert, J.P.; Vermersch, P.; Prin, L. Molecular analysis of endogenous retrovirus hres-1: Identification of frameshift mutations in region encoding putative 28-kda autoantigen. Biochem. Biophys. Res. Commun. 2001, 283, 437–444. [Google Scholar] [CrossRef]
- Bock, M.; Stoye, J.P. Endogenous retroviruses and the human germline. Curr. Opin. Genet. Dev. 2000, 10, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Vargas, A.; Moreau, J.; Landry, S.; LeBellego, F.; Toufaily, C.; Rassart, E.; Lafond, J.; Barbeau, B. Syncytin-2 plays an important role in the fusion of human trophoblast cells. J. Mol. Biol. 2009, 392, 301–318. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.H.; Handwerger, S. A placenta-specific enhancer of the human syncytin gene. Biol. Reprod. 2005, 73, 500–509. [Google Scholar] [CrossRef]
- Mi, S.; Lee, X.; Li, X.; Veldman, G.M.; Finnerty, H.; Racie, L.; LaVallie, E.; Tang, X.Y.; Edouard, P.; Howes, S.; et al. Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis. Nature 2000, 403, 785–789. [Google Scholar] [CrossRef]
- Bjerregaard, B.; Holck, S.; Christensen, I.J.; Larsson, L.I. Syncytin is involved in breast cancer-endothelial cell fusions. Cell. Mol. Life Sci. 2006, 63, 1906–1911. [Google Scholar] [CrossRef]
- Larsson, L.I.; Bjerregaard, B.; Wulf-Andersen, L.; Talts, J.F. Syncytin and cancer cell fusions. ScientificWorldJournal 2007, 7, 1193–1197. [Google Scholar] [CrossRef]
- Antony, J.M.; van Marle, G.; Opii, W.; Butterfield, D.A.; Mallet, F.; Yong, V.W.; Wallace, J.L.; Deacon, R.M.; Warren, K.; Power, C. Human endogenous retrovirus glycoprotein-mediated induction of redox reactants causes oligodendrocyte death and demyelination. Nat. Neurosci. 2004, 7, 1088–1095. [Google Scholar] [CrossRef]
- Weis, S.; Llenos, I.C.; Sabunciyan, S.; Dulay, J.R.; Isler, L.; Yolken, R.; Perron, H. Reduced expression of human endogenous retrovirus (HERV)-w gag protein in the cingulate gyrus and hippocampus in schizophrenia, bipolar disorder, and depression. J. Neural. Transm. 2007, 114, 645–655. [Google Scholar] [CrossRef]
- Yao, Y.; Schroder, J.; Nellaker, C.; Bottmer, C.; Bachmann, S.; Yolken, R.H.; Karlsson, H. Elevated levels of human endogenous retrovirus-w transcripts in blood cells from patients with first episode schizophrenia. Genes Brain Behav. 2008, 7, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.C.; Jacobson, S. Htlv-i associated myelopathy/tropical spastic paraparesis (HAM/TSP): A chronic progressive neurologic disease associated with immunologically mediated damage to the central nervous system. J. Neurovirol. 1997, 3, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Yamano, Y.; Nagai, M.; Brennan, M.; Mora, C.A.; Soldan, S.S.; Tomaru, U.; Takenouchi, N.; Izumo, S.; Osame, M.; Jacobson, S. Correlation of human T-cell lymphotropic virus type 1 (HTLV-1) mrna with proviral DNA load, virus-specific cd8(+) T cells, and disease severity in HTLV-1-associated myelopathy (HAM/TSP). Blood 2002, 99, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.P.; Franklin, A.A.; Uittenbogaard, M.N.; Giebler, H.A.; Nyborg, J.K. Pleiotropic effect of the human T-cell leukemia virus tax protein on the DNA binding activity of eukaryotic transcription factors. Proc. Natl. Acad. Sci. U. S. A. 1993, 90, 7303–7307. [Google Scholar] [CrossRef]
- Munoz, E.; Israel, A. Activation of nf-kappa b by the tax protein of HTLV-1. Immunobiology 1995, 193, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M. Mechanism of transcriptional activation of viral and cellular genes by oncogenic protein of HTLV-1. Leukemia 1994, 8, S51–S53. [Google Scholar]
- Higuchi, M.; Fujii, M. Distinct functions of HTLV-1 tax1 from HTLV-2 tax2 contribute key roles to viral pathogenesis. Retrovirology 2009, 6, 117. [Google Scholar] [CrossRef]
- Matsuoka, M.; Jeang, K.T. Human T-cell leukemia virus type 1 (HTLV-1) and leukemic transformation: Viral infectivity, TAX, HBZ and therapy. Oncogene 2011, 30, 1379–1389. [Google Scholar] [CrossRef]
- Kwun, H.J.; Han, H.J.; Lee, W.J.; Kim, H.S.; Jang, K.L. Transactivation of the human endogenous retrovirus K long terminal repeat by herpes simplex virus type 1 immediate early protein 0. Virus Res. 2002, 86, 93–100. [Google Scholar] [CrossRef]
- Lee, W.J.; Kwun, H.J.; Kim, H.S.; Jang, K.L. Activation of the human endogenous retrovirus W long terminal repeat by herpes simplex virus type 1 immediate early protein 1. Mol. Cells 2003, 15, 75–80. [Google Scholar]
- Nellaker, C.; Yao, Y.; Jones-Brando, L.; Mallet, F.; Yolken, R.H.; Karlsson, H. Transactivation of elements in the human endogenous retrovirus W family by viral infection. Retrovirology 2006, 3, 44. [Google Scholar] [CrossRef] [PubMed]
- Vargas, A.; Moreau, J.; Le Bellego, F.; Lafond, J.; Barbeau, B. Induction of trophoblast cell fusion by a protein tyrosine phosphatase inhibitor. Placenta 2008, 29, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Fortin, J.F.; Barbeau, B.; Robichaud, G.A.; Pare, M.E.; Lemieux, A.M.; Tremblay, M.J. Regulation of nuclear factor of activated T cells by phosphotyrosyl-specific phosphatase activity: A positive effect on HIV-1 long terminal repeat-driven transcription and a possible implication of shp-1. Blood 2001, 97, 2390–2400. [Google Scholar] [CrossRef] [PubMed]
- Barat, C.; Tremblay, M.J. Treatment of human t cells with bisperoxovanadium phosphotyrosyl phosphatase inhibitors leads to activation of cyclooxygenase-2 gene. J. Biol. Chem. 2003, 278, 6992–7000. [Google Scholar] [CrossRef] [PubMed]
- Blanchette, J.; Pouliot, P.; Olivier, M. Role of protein tyrosine phosphatases in the regulation of interferon-{gamma}-induced macrophage nitric oxide generation: Implication of erk pathway and ap-1 activation. J. Leukoc. Biol. 2007, 81, 835–844. [Google Scholar] [CrossRef]
- Kelleher, C.A.; Wilkinson, D.A.; Freeman, J.D.; Mager, D.L.; Gelfand, E.W. Expression of novel-transposon-containing mrnas in human t cells. J. Gen. Virol. 1996, 77, 1101–1110. [Google Scholar] [CrossRef]
- Johnston, J.B.; Silva, C.; Holden, J.; Warren, K.G.; Clark, A.W.; Power, C. Monocyte activation and differentiation augment human endogenous retrovirus expression: Implications for inflammatory brain diseases. Ann. Neurol. 2001, 50, 434–442. [Google Scholar] [CrossRef]
- Lawoko, A.; Johansson, B.; Rabinayaran, D.; Pipkorn, R.; Blomberg, J. Increased immunoglobulin G, but not m, binding to endogenous retroviral antigens in HIV-1 infected persons. J. Med. Virol. 2000, 62, 435–444. [Google Scholar] [CrossRef]
- Garrison, K.E.; Jones, R.B.; Meiklejohn, D.A.; Anwar, N.; Ndhlovu, L.C.; Chapman, J.M.; Erickson, A.L.; Agrawal, A.; Spotts, G.; Hecht, F.M.; et al. T cell responses to human endogenous retroviruses in HIV-1 infection. PLoS Pathog. 2007, 3, e165. [Google Scholar] [CrossRef]
- Contreras, X.; Lenasi, T.; Peterlin, B.M. HIV latency: Present knowledge, future directions. Future Virol. 2006, 1, 733–745. [Google Scholar] [CrossRef]
- Contreras, X.; Barboric, M.; Lenasi, T.; Peterlin, B.M. Hmba releases p-tefb from hexim1 and 7sk snrna via pi3k/akt and activates HIV transcription. PLoS Pathog. 2007, 3, 1459–1469. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Shen, K.; Lin, M.; Chen, P.; Lin, C.; Chang, G.D.; Chen, H. Gcma regulates the syncytin-mediated trophoblastic fusion. J. Biol. Chem. 2002, 277, 50062–50068. [Google Scholar] [CrossRef] [PubMed]
- Prudhomme, S.; Oriol, G.; Mallet, F. A retroviral promoter and a cellular enhancer define a bipartite element which controls env ervwe1 placental expression. J. Virol. 2004, 78, 12157–12168. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.H.; Richardson, B.D.; Hubert, M.A.; Handwerger, S. Isolation and characterization of the human syncytin gene promoter. Biol. Reprod. 2004, 70, 694–701. [Google Scholar] [CrossRef]
- Mameli, G.; Astone, V.; Khalili, K.; Serra, C.; Sawaya, B.E.; Dolei, A. Regulation of the syncytin-1 promoter in human astrocytes by multiple sclerosis-related cytokines. Virology 2007, 362, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Schon, U.; Seifarth, W.; Baust, C.; Hohenadl, C.; Erfle, V.; Leib-Mosch, C. Cell type-specific expression and promoter activity of human endogenous retroviral long terminal repeats. Virology 2001, 279, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Shibata, H.; Fujisawa, J.I.; Inoue, H.; Hakura, A.; Tsukahara, T.; Fujii, M. Human t-cell leukemia virus type 1 tax protein transforms rat fibroblasts via two distinct pathways. J. Virol. 1997, 71, 4445–4451. [Google Scholar] [CrossRef]
- Rousset, R.; Desbois, C.; Bantignies, F.; Jalinot, P. Effects on nf-kappa b1/p105 processing of the interaction between the HTLV-1 transactivator tax and the proteasome. Nature 1996, 381, 328–331. [Google Scholar] [CrossRef]
- Walton, K.M.; Rehfuss, R.P.; Chrivia, J.C.; Lochner, J.E.; Goodman, R.H. A dominant repressor of cyclic adenosine 3′,5′-monophosphate (camp)-regulated enhancer-binding protein activity inhibits the camp-mediated induction of the somatostatin promoter in vivo. Mol. Endocrinol. 1992, 6, 647–655. [Google Scholar]
- Gachon, F.; Thebault, S.; Peleraux, A.; Devaux, C.; Mesnard, J.M. Molecular interactions involved in the transactivation of the human T-cell leukemia virus type 1 promoter mediated by tax and creb-2 (atf-4). Mol. Cell Biol. 2000, 20, 3470–3481. [Google Scholar] [CrossRef]
- Balada, E.; Ordi-Ros, J.; Vilardell-Tarres, M. Molecular mechanisms mediated by human endogenous retroviruses (HERVs) in autoimmunity. Rev. Med. Virol. 2009, 19, 273–286. [Google Scholar] [CrossRef] [PubMed]





© 2011 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Toufaily, C.; Landry, S.; Leib-Mosch, C.; Rassart, E.; Barbeau, B. Activation of LTRs from Different Human Endogenous Retrovirus (HERV) Families by the HTLV-1 Tax Protein and T-Cell Activators. Viruses 2011, 3, 2146-2159. https://doi.org/10.3390/v3112146
Toufaily C, Landry S, Leib-Mosch C, Rassart E, Barbeau B. Activation of LTRs from Different Human Endogenous Retrovirus (HERV) Families by the HTLV-1 Tax Protein and T-Cell Activators. Viruses. 2011; 3(11):2146-2159. https://doi.org/10.3390/v3112146
Chicago/Turabian StyleToufaily, Chirine, Sebastien Landry, Christine Leib-Mosch, Eric Rassart, and Benoit Barbeau. 2011. "Activation of LTRs from Different Human Endogenous Retrovirus (HERV) Families by the HTLV-1 Tax Protein and T-Cell Activators" Viruses 3, no. 11: 2146-2159. https://doi.org/10.3390/v3112146
APA StyleToufaily, C., Landry, S., Leib-Mosch, C., Rassart, E., & Barbeau, B. (2011). Activation of LTRs from Different Human Endogenous Retrovirus (HERV) Families by the HTLV-1 Tax Protein and T-Cell Activators. Viruses, 3(11), 2146-2159. https://doi.org/10.3390/v3112146