Direct Inhibition of RNAse T2 Expression by the HTLV-1 Viral Protein Tax

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
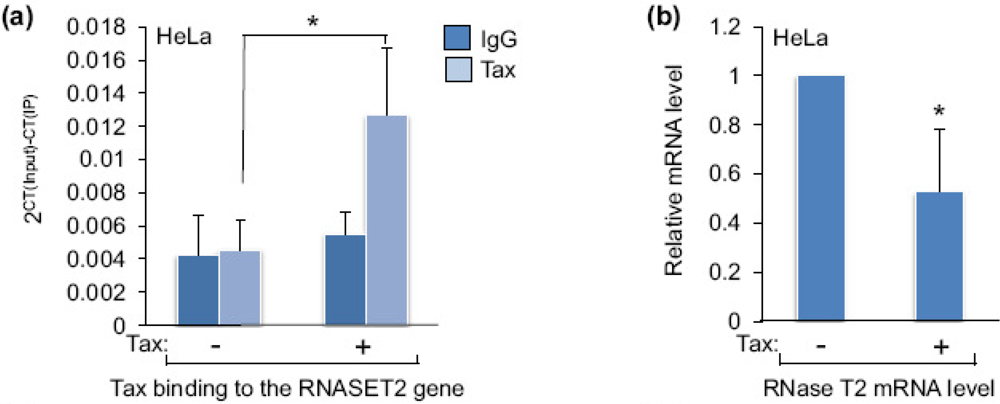
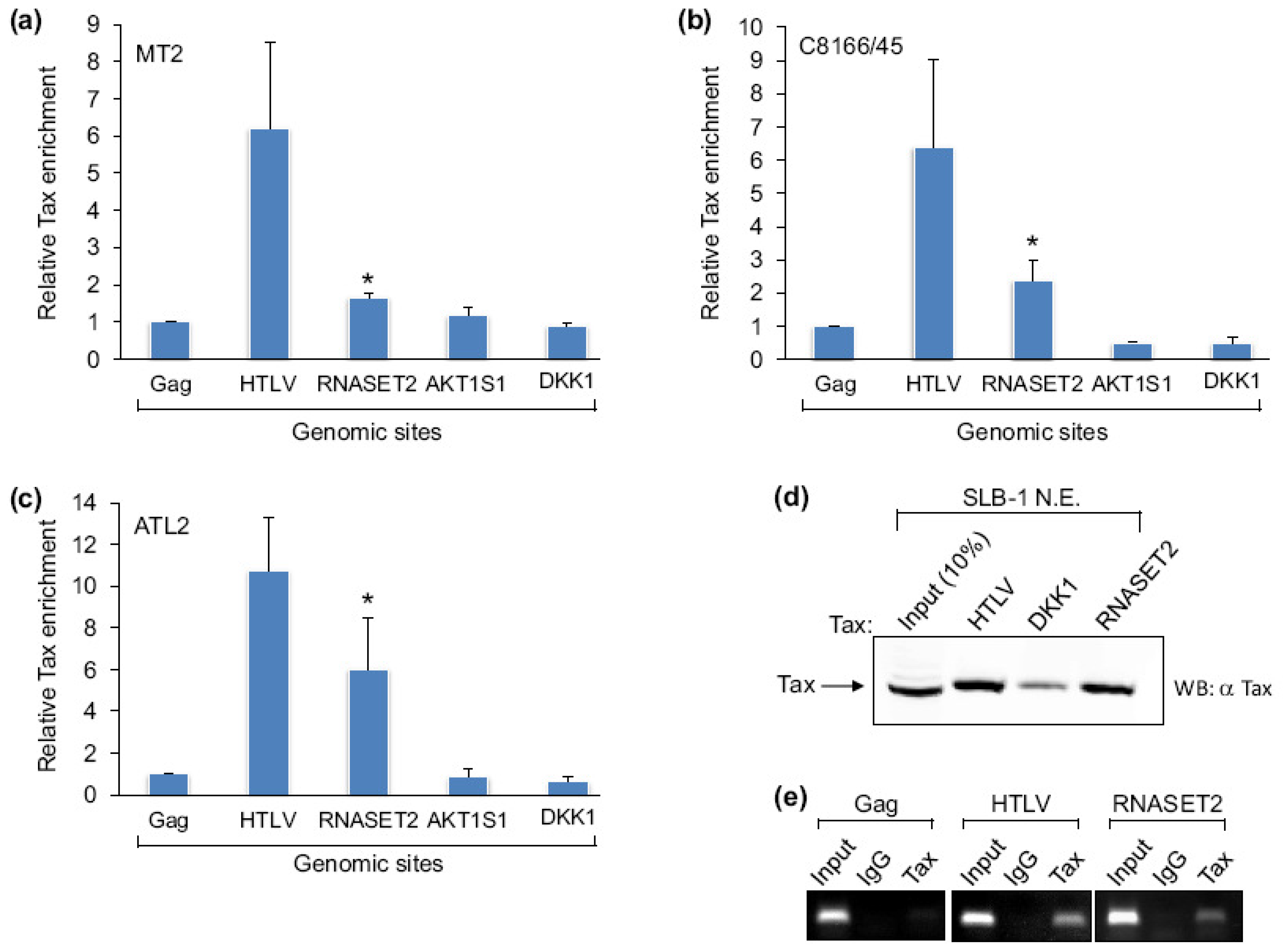
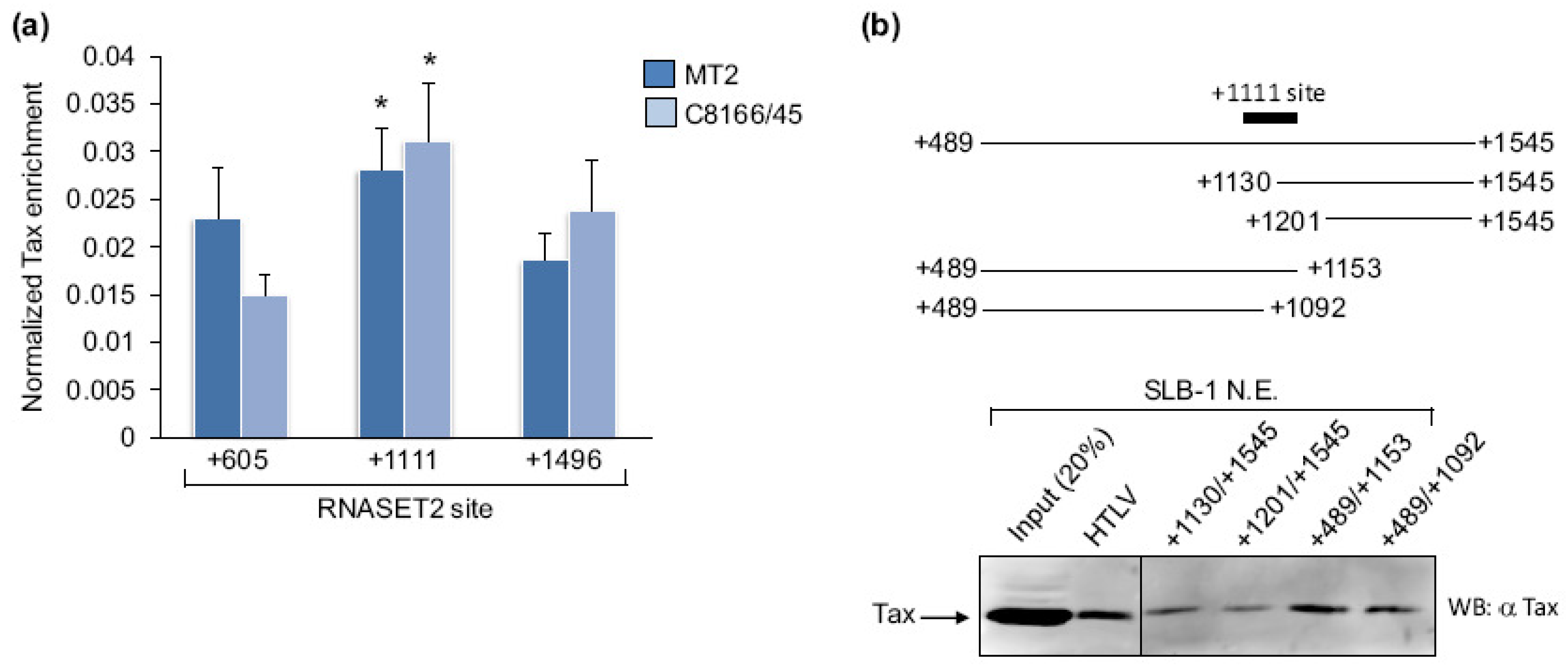
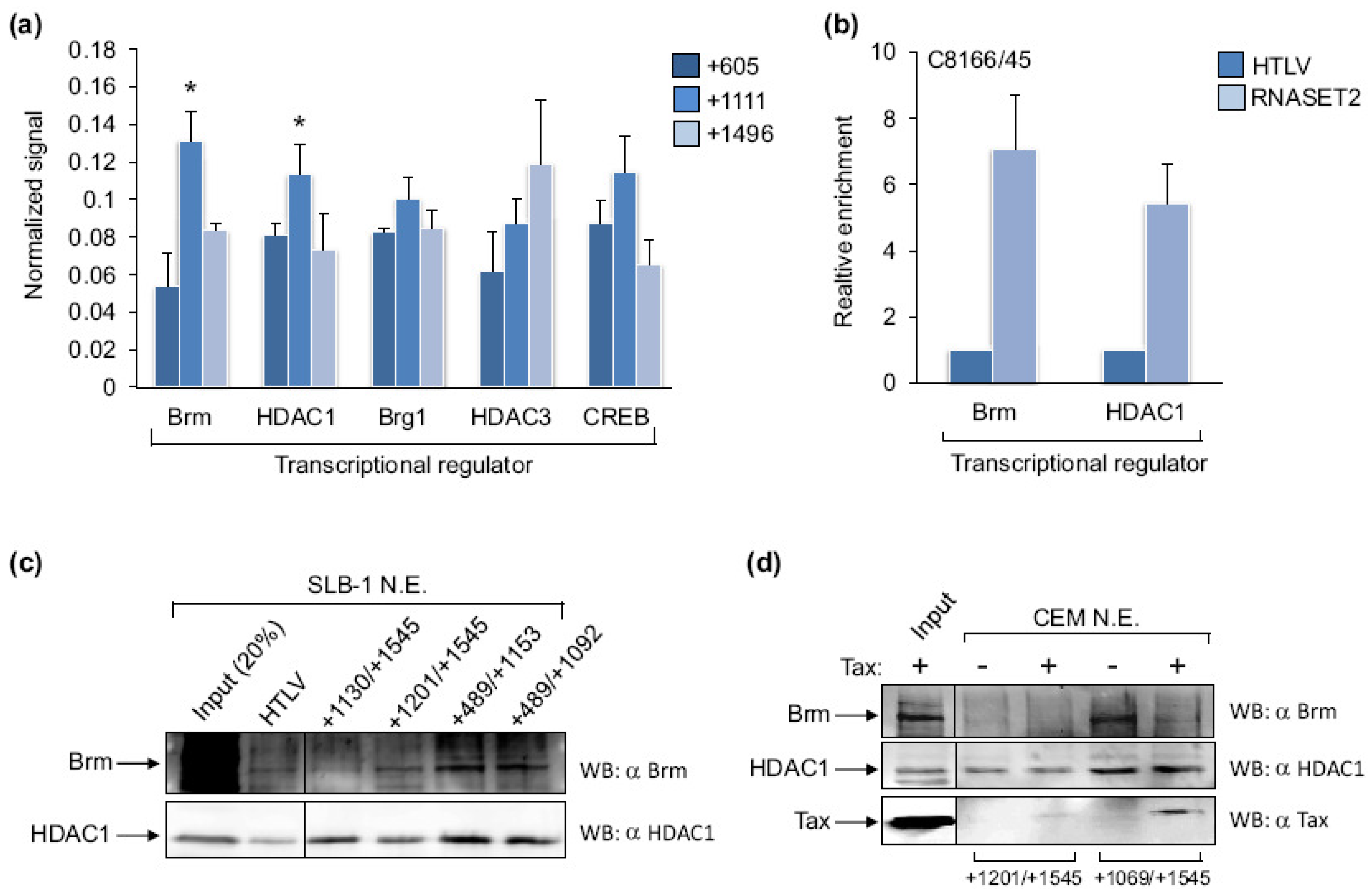
2.1. Tax Is Enriched at the RNASET2 Gene
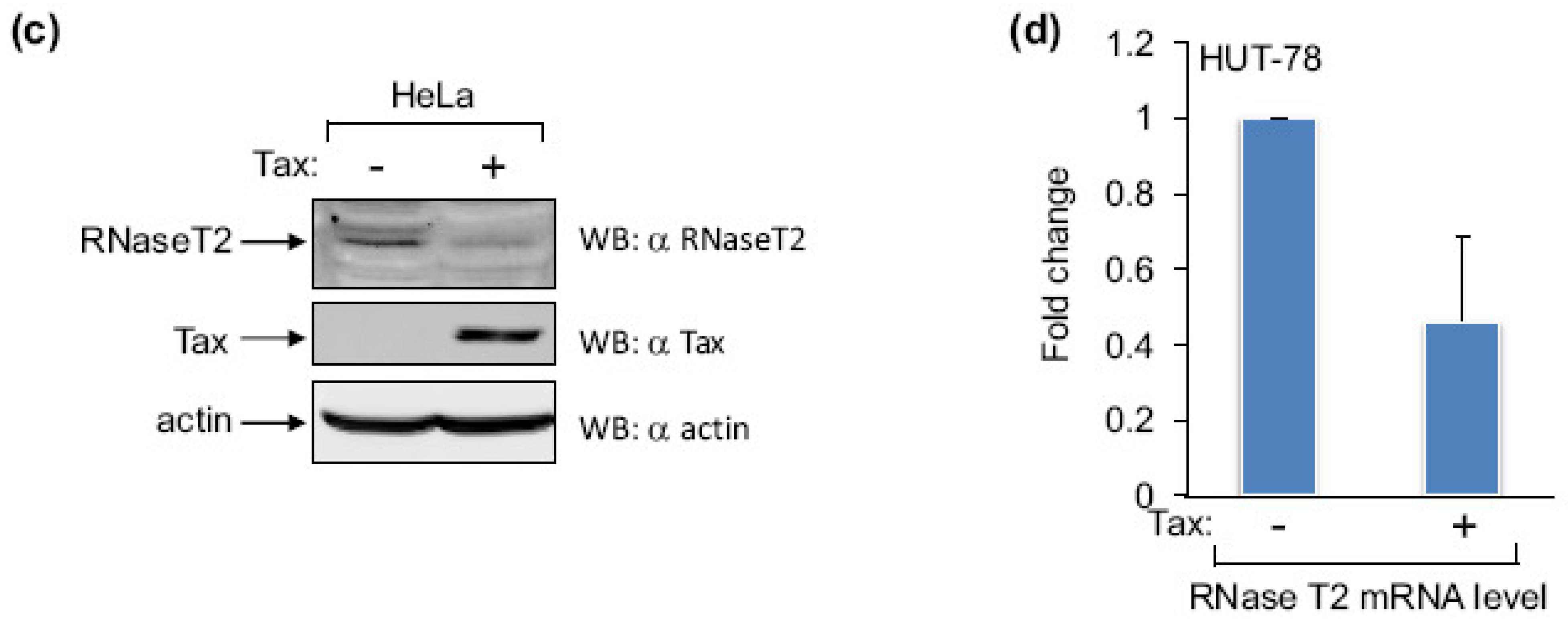
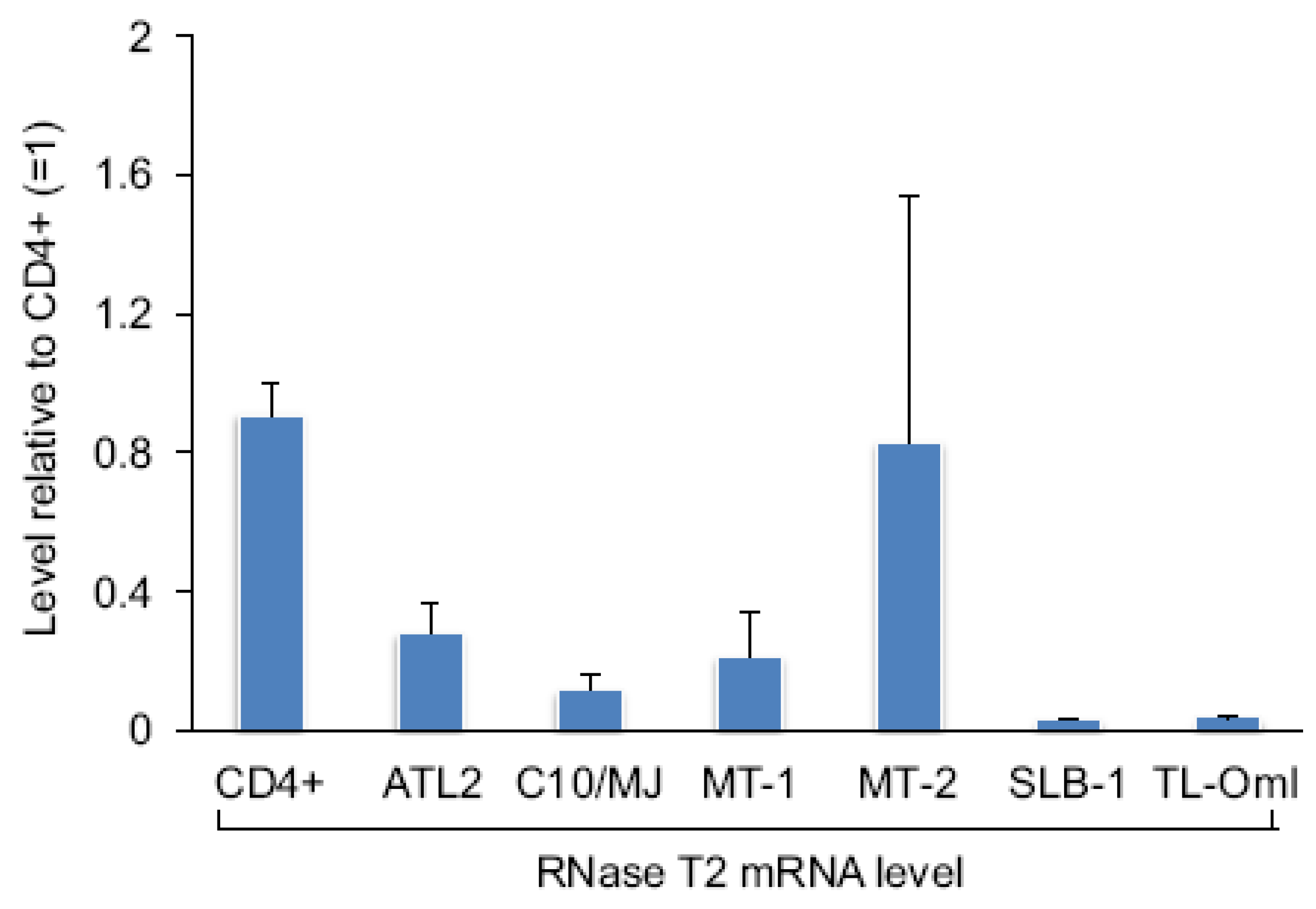
2.2. Tax Represses Expression of RNase T2
2.3. Brm1, HDAC1 and Tax Exhibit Overlapping Patterns of Enrichment on the RNASET2 Gene
3. Experimental Section
3.1. Cell Culture, Transfection and Western Blot Analysis
3.2. Chromatin Immunoprecipitation (ChIP) Assays and Microarray Analysis
3.3. Real-Time PCR
- vCRE-F, 5′-ATCATAAGCTCAGACCTCCGGGAA; vCRE-R,
- 5′-CCTGAGGACGGCTTGACAAACAT; Gag-F, 5-CGCTCATCACTGGCTTAACTTCCT; Gag-R,
- 5′-TGGTGGAAATCGTAACTGGAGGGA; RNaseT2+1496F,
- 5′-CGGTGCTGCAAAGAACTCCAACTT; RNaseT2+1496R,
- 5′-TCCTATTAACCCGATCATTACCCA; RNaseT2+1111F,
- 5′-AGCCAAGGGAAGGGATTCAGAACA; RNaseT2+1111R,
- 5′-TAATTTCAGCTTTCACTGCGGCGG; RNaseT2+605F,
- 5′-CGCTGCTCAACTTCCCAACTTCTT; RNaseT2+605R,
- 5′-CCCTAGAGCTTTCCACTGGGTGAA; DKK1-331F and DKK1-331R [28].
3.4. Immobilized Template-Binding Assays
4. Conclusions
Acknowledgements
References and Notes
- Mahieux, R.; Gessain, A. Adult T-cell leukemia/lymphoma and HTLV-1. Curr. Hematol. Malig. Rep. 2007, 2, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Azran, I.; Schavinsky-Khrapunsky, Y.; Aboud, M. Role of Tax protein in human T-cell leukemia virus type-I leukemogenicity. Retrovirology 2004, 1, 20. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, H.; Sawa, H.; Lewis, M.J.; Orba, Y.; Sheehy, N.; Yamamoto, Y.; Ichinohe, T.; Tsunetsugu-Yokota, Y.; Katano, H.; Takahashi, H.; et al. Thymus-derived leukemia-lymphoma in mice transgenic for the Tax gene of human T-lymphotropic virus type I. Nat. Med. 2006, 12, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Marriott, S.J.; Semmes, O.J. Impact of HTLV-I Tax on cell cycle progression and the cellular DNA damage repair response. Oncogene 2005, 24, 5986–5995. [Google Scholar] [CrossRef]
- Polakowski, N.; Lemasson, I. Regulation of HTLV-1 transcription by viral and cellular proteins. In Recent Advances in Human Retroviruses: Principles of Replication and Pathogenesis. Advances in Retroviral Research; Lever, A.M.L., Jeang, K.T., Berkhout, B., Eds.; World Scientific Publishing Co.: Singapore, 2010; pp. 129–169. [Google Scholar]
- Wang, D.; Guo, M.X.; Hu, H.M.; Zhao, Z.Z.; Qiu, H.L.; Shao, H.J.; Zhu, C.G.; Xue, L.; Shi, Y.B.; Li, W.X. Human T-cell leukemia virus type 1 oncoprotein tax represses ZNF268 expression through the cAMP-responsive element-binding protein/activating transcription factor pathway. J. Biol. Chem. 2008, 283, 16299–16308. [Google Scholar] [CrossRef] [PubMed]
- Kibler, K.V.; Jeang, K.T. CREB/ATF-dependent repression of cyclin a by human T-cell leukemia virus type 1 Tax protein. J. Virol. 2001, 75, 2161–2173. [Google Scholar] [CrossRef] [PubMed]
- Van Orden, K.; Nyborg, J.K. Insight into the tumor suppressor function of CBP through the viral oncoprotein Tax. Gene Expr. 2000, 9, 29–36. [Google Scholar] [CrossRef]
- Riou, P.; Bex, F.; Gazzolo, L. The human T cell leukemia/lymphotropic virus type 1 Tax protein represses MyoD-dependent transcription by inhibiting MyoD-binding to the KIX domain of p300. A potential mechanism for Tax-mediated repression of the transcriptional activity of basic helix-loop-helix factors. J. Biol. Chem. 2000, 275, 10551–10560. [Google Scholar]
- Colgin, M.A.; Nyborg, J.K. The human T-cell leukemia virus type 1 oncoprotein Tax inhibits the transcriptional activity of c-Myb through competition for the CREB binding protein. J. Virol. 1998, 72, 9396–9399. [Google Scholar] [CrossRef]
- Nicot, C.; Opavsky, R.; Mahieux, R.; Johnson, J.M.; Brady, J.N.; Wolff, L.; Franchini, G. Tax oncoprotein trans-represses endogenous B-myb promoter activity in human T cells. AIDS Res. Hum. Retroviruses 2000, 16, 1629–1632. [Google Scholar] [CrossRef]
- Boxus, M.; Twizere, J.C.; Legros, S.; Dewulf, J.F.; Kettmann, R.; Willems, L. The HTLV-1 Tax interactome. Retrovirology 2008, 5, 76. [Google Scholar] [CrossRef] [PubMed]
- Chlichlia, K.; Khazaie, K. HTLV-1 Tax: Linking transformation, DNA damage and apoptotic T-cell death. Chem. Biol. Interact. 2010, 188, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.L.; Giam, C.Z. Activation of the anaphase promoting complex by HTLV-1 tax leads to senescence. EMBO J. 2006, 25, 1741–1752. [Google Scholar] [CrossRef]
- Tripp, A.; Banerjee, P.; Sieburg, M.; Planelles, V.; Li, F.; Feuer, G. Induction of cell cycle arrest by human T-cell lymphotropic virus type 1 Tax in hematopoietic progenitor (CD34+) cells: Modulation of p21cip1/waf1 and p27kip1 expression. J. Virol. 2005, 79, 14069–14078. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Kotomura, N.; Ho, Y.K.; Zhi, H.; Bixler, S.; Schell, M.J.; Giam, C.Z. Complex cell cycle abnormalities caused by human T-lymphotropic virus type 1 Tax. J. Virol. 2011, 85, 3001–3009. [Google Scholar] [CrossRef]
- Acquati, F.; Morelli, C.; Cinquetti, R.; Bianchi, M.G.; Porrini, D.; Varesco, L.; Gismondi, V.; Rocchetti, R.; Talevi, S.; Possati, L.; et al. Cloning and characterization of a senescence inducing and class II tumor suppressor gene in ovarian carcinoma at chromosome region 6q27. Oncogene 2001, 20, 980–988. [Google Scholar] [CrossRef]
- Luhtala, N.; Parker, R. T2 Family ribonucleases: Ancient enzymes with diverse roles. Trends Biochem. Sci. 2010, 35, 253–259. [Google Scholar] [CrossRef]
- Trubia, M.; Sessa, L.; Taramelli, R. Mammalian Rh/T2/S-glycoprotein ribonuclease family genes: Cloning of a human member located in a region of chromosome 6 (6q27) frequently deleted in human malignancies. Genomics 1997, 42, 342–344. [Google Scholar] [CrossRef]
- Tsukasaki, K.; Krebs, J.; Nagai, K.; Tomonaga, M.; Koeffler, H.P.; Bartram, C.R.; Jauch, A. Comparative genomic hybridization analysis in adult T-cell leukemia/lymphoma: Correlation with clinical course. Blood 2001, 97, 3875–3881. [Google Scholar] [CrossRef]
- Liu, Y.; Emilion, G.; Mungall, A.J.; Dunham, I.; Beck, S.; Le Meuth-Metzinger, V.G.; Shelling, A.N.; Charnock, F.M.; Ganesan, T.S. Physical and transcript map of the region between D6S264 and D6S149 on chromosome 6q27, the minimal region of allele loss in sporadic epithelial ovarian cancer. Oncogene 2002, 21, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Morin, P.J. A novel homozygous deletion at chromosomal band 6q27 in an ovarian cancer cell line delineates the position of a putative tumor suppressor gene. Cancer Lett. 2001, 173, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Monti, L.; Rodolfo, M.; Lo Russo, G.; Noonan, D.; Acquati, F.; Taramelli, R. RNASET2 as a tumor antagonizing gene in a melanoma cancer model. Oncol. Res. 2008, 17, 69–74. [Google Scholar] [CrossRef]
- Acquati, F.; Possati, L.; Ferrante, L.; Campomenosi, P.; Talevi, S.; Bardelli, S.; Margiotta, C.; Russo, A.; Bortoletto, E.; Rocchetti, R.; et al. Tumor and metastasis suppression by the human RNASET2 gene. Int. J. Oncol. 2005, 26, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhawar, V.K.; Kaur, G.; Kaur, G.P.; Deriel, J.K.; Kandpal, R.P.; Athwal, R.S. Chromosome 6 encoded RNaseT2 protein is a cell growth regulator. J. Cell Mol. Med. 2010, 14, 1146–1155. [Google Scholar] [CrossRef] [PubMed]
- Acquati, F.; Bertilaccio, S.; Grimaldi, A.; Monti, L.; Cinquetti, R.; Bonetti, P.; Lualdi, M.; Vidalino, L.; Fabbri, M.; Sacco, M.G.; et al. Microenvironmental control of malignancy exerted by RNASET2, a widely conserved extracellular RNase. Proc. Natl. Acad. Sci. U. S. A. 2010, 108, 1104–1109. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, P.; Roiz, L.; Angelkovitch, B.; Schwartz, B.; Shoseyov, O. A recombinant human RNASET2 glycoprotein with antitumorigenic and antiangiogenic characteristics: Expression, purification, and characterization. Cancer 2006, 107, 2760–2769. [Google Scholar] [CrossRef]
- Polakowski, N.; Gregory, H.; Mesnard, J.M.; Lemasson, I. Expression of a protein involved in bone resorption, Dkk1, is activated by HTLV-1 bZIP factor through its activation domain. Retrovirology 2010, 7, 61. [Google Scholar] [CrossRef]
- Cartharius, K.; Frech, K.; Grote, K.; Klocke, B.; Haltmeier, M.; Klingenhoff, A.; Frisch, M.; Bayerlein, M.; Werner, T. MatInspector and beyond: Promoter analysis based on transcription factor binding sites. Bioinformatics 2005, 21, 2933–2942. [Google Scholar] [CrossRef]
- Mori, N.; Fujii, M.; Ikeda, S.; Yamada, Y.; Tomonaga, M.; Ballard, D.W.; Yamamoto, N. Constitutive activation of NF-kappaB in primary adult T-cell leukemia cells. Blood 1999, 93, 2360–2368. [Google Scholar]
- Hironaka, N.; Mochida, K.; Mori, N.; Maeda, M.; Yamamoto, N.; Yamaoka, S. Tax-independent constitutive IkappaB kinase activation in adult T-cell leukemia cells. Neoplasia 2004, 6, 266–278. [Google Scholar] [CrossRef]
- Narlikar, G.J.; Fan, H.Y.; Kingston, R.E. Cooperation between complexes that regulate chromatin structure and transcription. Cell 2002, 108, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Bottazzi, M.E.; de la Fuente, C.; Deng, L.; Gitlin, S.D.; Maddukuri, A.; Dadgar, S.; Li, H.; Vertes, A.; Pumfery, A.; et al. Protein profile of tax-associated complexes. J. Biol. Chem. 2004, 279, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Ego, T.; Ariumi, Y.; Shimotohno, K. The interaction of HTLV-1 Tax with HDAC1 negatively regulates the viral gene expression. Oncogene 2002, 21, 7241–7246. [Google Scholar] [CrossRef]
- Villanueva, R.; Iglesias, A.H.; Camelo, S.; Sanin, L.C.; Gray, S.G.; Dangond, F. Histone deacetylase 3 represses HTLV-1 tax transcription. Oncol. Rep. 2006, 16, 581–585. [Google Scholar] [CrossRef]
- Riccio, A. New endogenous regulators of class I histone deacetylases. Sci. Signal 2010, 3, pe1. [Google Scholar] [CrossRef]
- Clerc, I.; Polakowski, N.; Andre-Arpin, C.; Cook, P.; Barbeau, B.; Mesnard, J.M.; Lemasson, I. An interaction between the human T cell leukemia virus type 1 basic leucine zipper factor (HBZ) and the KIX domain of p300/CBP contributes to the down-regulation of tax-dependent viral transcription by HBZ. J. Biol. Chem. 2008, 283, 23903–23913. [Google Scholar] [CrossRef] [PubMed]
- Lemasson, I.; Polakowski, N.J.; Laybourn, P.J.; Nyborg, J.K. Tax-dependent displacement of nucleosomes during transcriptional activation of human T-cell leukemia virus type 1. J. Biol. Chem. 2006, 281, 13075–13082. [Google Scholar] [CrossRef] [PubMed]
- Rousset, R.; Desbois, C.; Bantignies, F.; Jalinot, P. Effects on NF-kappa B1/p105 processing of the interaction between the HTLV-1 transactivator Tax and the proteasome. Nature 1996, 381, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Lemasson, I.; Nyborg, J.K. Human T-cell leukemia virus type I Tax repression of p73beta is mediated through competition for the C/H1 domain of CBP. J. Biol. Chem. 2001, 276, 15720–15727. [Google Scholar] [CrossRef]
- Lemasson, I.; Polakowski, N.; Laybourn, P.J.; Nyborg, J.K. Transcription factor binding and histone modifications on the integrated proviral promoter in HTLV-I-infected T-cells. J. Biol. Chem. 2002, 277, 49459–49465. [Google Scholar] [CrossRef]
- Oberley, M.J.; Tsao, J.; Yau, P.; Farnham, P.J. High-throughput screening of chromatin immunoprecipitates using CpG-island microarrays. Methods Enzymol. 2004, 376, 315–334. [Google Scholar] [PubMed]
- Cross, S.H.; Charlton, J.A.; Nan, X.; Bird, A.P. Purification of CpG islands using a methylated DNA binding column. Nat. Genet. 1994, 6, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Dynlacht, B.D. Use of chromatin immunoprecipitation assays in genome-wide location analysis of mammalian transcription factors. Methods Enzymol. 2004, 376, 304–315. [Google Scholar] [PubMed]
- Yie, J.; Senger, K.; Thanos, D. Mechanism by which the IFN-beta enhanceosome activates transcription. Proc. Natl. Acad. Sci. U. S. A. 1999, 96, 13108–13113. [Google Scholar] [CrossRef] [PubMed]
- Dignam, J.D.; Lebovitz, R.M.; Roeder, R.G. Accurate transcription initiation by RNA polymerase II in a soluble extract from isolated mammalian nuclei. Nucleic Acids Res. 1983, 11, 1475–1489. [Google Scholar] [CrossRef]
- Zhao, L.J.; Giam, C.Z. Interaction of the human T-cell lymphotrophic virus type I (HTLV-I) transcriptional activator Tax with cellular factors that bind specifically to the 21-base-pair repeats in the HTLV-I enhancer. Proc. Natl. Acad. Sci. U. S. A. 1991, 88, 11445–11449. [Google Scholar] [CrossRef]
- Kim, Y.M.; Geiger, T.R.; Egan, D.I.; Sharma, N.; Nyborg, J.K. The HTLV-1 tax protein cooperates with phosphorylated CREB, TORC2 and p300 to activate CRE-dependent cyclin D1 transcription. Oncogene 2010, 29, 2142–2152. [Google Scholar] [CrossRef]
- Cheng, J.; Kydd, A.R.; Nakase, K.; Noonan, K.M.; Murakami, A.; Tao, H.; Dwyer, M.; Xu, C.; Zhu, Q.; Marasco, W.A. Negative regulation of the SH2-homology containing protein-tyrosine phosphatase-1 (SHP-1) P2 promoter by the HTLV-1 Tax oncoprotein. Blood 2007, 110, 2110–2120. [Google Scholar] [CrossRef]
- de la Fuente, C.; Gupta, M.V.; Klase, Z.; Strouss, K.; Cahan, P.; McCaffery, T.; Galante, A.; Soteropoulos, P.; Pumfery, A.; Fujii, M.; et al. Involvement of HTLV-I Tax and CREB in aneuploidy: A bioinformatics approach. Retrovirology 2006, 3, 43. [Google Scholar] [CrossRef]
- Jeang, K.T.; Widen, S.G.; Semmes, O.J.; Wilson, S.H. HTLV-I trans-activator protein, Tax, is a trans-repressor of the human b-polymerase gene. Science 1990, 247, 1082–1084. [Google Scholar] [CrossRef]
- Lemasson, I.; Robert-Hebmann, V.; Hamaia, S.; Duc Dodon, M.; Gazzolo, L.; Devaux, C. Transrepression of lck gene expression by human T-cell leukemia virus type 1-encoded p40Tax. J. Virol. 1997, 71, 1975–1983. [Google Scholar] [CrossRef] [PubMed]
- Uittenbogaard, M.N.; Giebler, H.A.; Reisman, D.; Nyborg, J.K. Transcriptional repression of p53 by human T-cell leukemia virus type I Tax protein. J. Biol. Chem. 1995, 270, 28503–28506. [Google Scholar] [CrossRef]
- Suzuki, T.; Narita, T.; Uchida-Toita, M.; Yoshida, M. Down-regulation of the INK4 family of cyclin-dependent kinase inhibitors by tax protein of HTLV-1 through two distinct mechanisms. Virology 1999, 259, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.T.; Chun, A.C.; Ching, Y.P.; Jeang, K.T.; Jin, D.Y. Human T-cell leukemia virus oncoprotein tax represses nuclear receptor-dependent transcription by targeting coactivator TAX1BP1. Cancer Res. 2007, 67, 1072–1081. [Google Scholar] [CrossRef] [PubMed]
- Kinjo, T.; Ham-Terhune, J.; Peloponese, J.M., Jr.; Jeang, K.T. Induction of reactive oxygen species by human T-cell leukemia virus type 1 tax correlates with DNA damage and expression of cellular senescence marker. J. Virol. 2010, 84, 5431–5437. [Google Scholar] [CrossRef] [PubMed]
- Los, M.; Khazaie, K.; Schulze-Osthoff, K.; Baeuerle, P.A.; Schirrmacher, V.; Chlichlia, K. Human T cell leukemia virus-I (HTLV-I) Tax-mediated apoptosis in activated T cells requires an enhanced intracellular prooxidant state. J. Immunol. 1998, 161, 3050–3055. [Google Scholar] [CrossRef]
- Thompson, D.M.; Parker, R. The RNase Rny1p cleaves tRNAs and promotes cell death during oxidative stress in Saccharomyces cerevisiae. J. Cell Biol. 2009, 185, 43–50. [Google Scholar] [CrossRef]
- Masutani, H.; Hirota, K.; Sasada, T.; Ueda-Taniguchi, Y.; Taniguchi, Y.; Sono, H.; Yodoi, J. Transactivation of an inducible anti-oxidative stress protein, human thioredoxin by HTLV-I Tax. Immunol. Lett. 1996, 54, 67–71. [Google Scholar] [CrossRef]






© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Polakowski, N.; Han, H.; Lemasson, I. Direct Inhibition of RNAse T2 Expression by the HTLV-1 Viral Protein Tax. Viruses 2011, 3, 1485-1500. https://doi.org/10.3390/v3081485
Polakowski N, Han H, Lemasson I. Direct Inhibition of RNAse T2 Expression by the HTLV-1 Viral Protein Tax. Viruses. 2011; 3(8):1485-1500. https://doi.org/10.3390/v3081485
Chicago/Turabian StylePolakowski, Nicholas, Hongjin Han, and Isabelle Lemasson. 2011. "Direct Inhibition of RNAse T2 Expression by the HTLV-1 Viral Protein Tax" Viruses 3, no. 8: 1485-1500. https://doi.org/10.3390/v3081485
APA StylePolakowski, N., Han, H., & Lemasson, I. (2011). Direct Inhibition of RNAse T2 Expression by the HTLV-1 Viral Protein Tax. Viruses, 3(8), 1485-1500. https://doi.org/10.3390/v3081485