Comparative Genomics of Korean Infectious Bronchitis Viruses (IBVs) and an Animal Model to Evaluate Pathogenicity of IBVs to the Reproductive Organs

Abstract
:1. Introduction
2. Results and Discussion
2.1. Complete Genome Sequence Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Location | The length of protein(aa) | ||
|---|---|---|---|---|
| SNU8067 | KM91 | SNU8067 | KM91 | |
| 1a | 529–12390 | 529–12387 | 3953 | 3952 |
| 1b | 12465–20423 | 12357–20420 | 2652 | 2688 |
| S | 20374–23883 | 20371–23862 | 1169 | 1163 |
| 3a | 23883–24056 | 23862–24008 | 57 | 48 |
| 3b | 24056–24250 | 23995–24183 | 64 | 62 |
| E | 24234–24560 | 24167–24496 | 108 | 109 |
| M | 24532–25209 | 24465–25145 | 225 | 226 |
| 5a | 25569–25766 | 25505–25702 | 65 | 65 |
| 5b | 25763–26011 | 25699–25947 | 82 | 82 |
| N | 25954–27183 | 25890–27119 | 409 | 409 |
| Strain | Nucleotide and amino acid sequence identity (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1a | 1b | S1 | S2 | 3a | 3b | E | M | 5a | 5b | N | |
| Beaudette | 90/92 a | 92/96 | 79/78 | 85/87 | 8584 | 86/72 | 84/89 | 89/90 | 92/92 | 96/92 | 90/93 |
| ArkDPI11 | 91/92 | 94/97 | 82/81 | 87/89 | 91/88 | 93/92 | 85/91 | 91/91 | 97/97 | 97/93 | 93/96 |
| H120 | 90/91 | 93/97 | 79/76 | 85/87 | 82/81 | 86/74 | 83/91 | 91/91 | 93/93 | 95/89 | 91/94 |
| SAIBK | 87/89 | 91/96 | 78/76 | 94/96 | 82/74 | 80/71 | 86/87 | 87/89 | 83/83 | 94/89 | 87/93 |
| GD S14 | 83/85 | 89/95 | 79/77 | 95/96 | 87/90 | 73/66 | 85/88 | 88/89 | 84/84 | 91/88 | 89/93 |
| Conn46 | 93/94 | 93/97 | 78/76 | 87/90 | 95/93 | 93/94 | 85/91 | 91/91 | 97/97 | 98/94 | 93/96 |
| M41 | 91/92 | 92/96 | 79/76 | 86/87 | 84/79 | 87/74 | 84/89 | 89/91 | 88/88 | 96/89 | 91/94 |
| LX4 | 84/86 | 90/96 | 76/74 | 90/93 | 80/71 | 73/65 | 84/90 | 90/91 | 81/81 | 91/90 | 89/94 |
| KM91 | 96/96b | 97/98 | 80/77 | 97/97 | 65/52 | 81/72 | 95/94 | 94/96 | 95/95 | 97/92 | 95/96 |
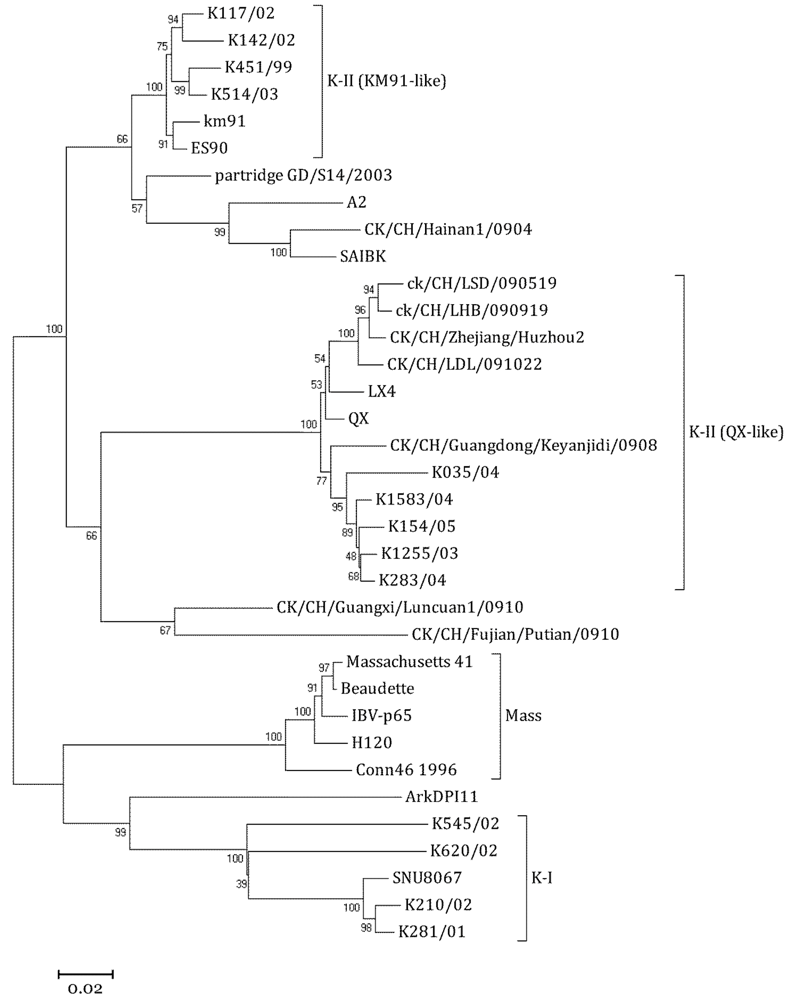
2.2. Phylogenetic Analysis of SNU8067
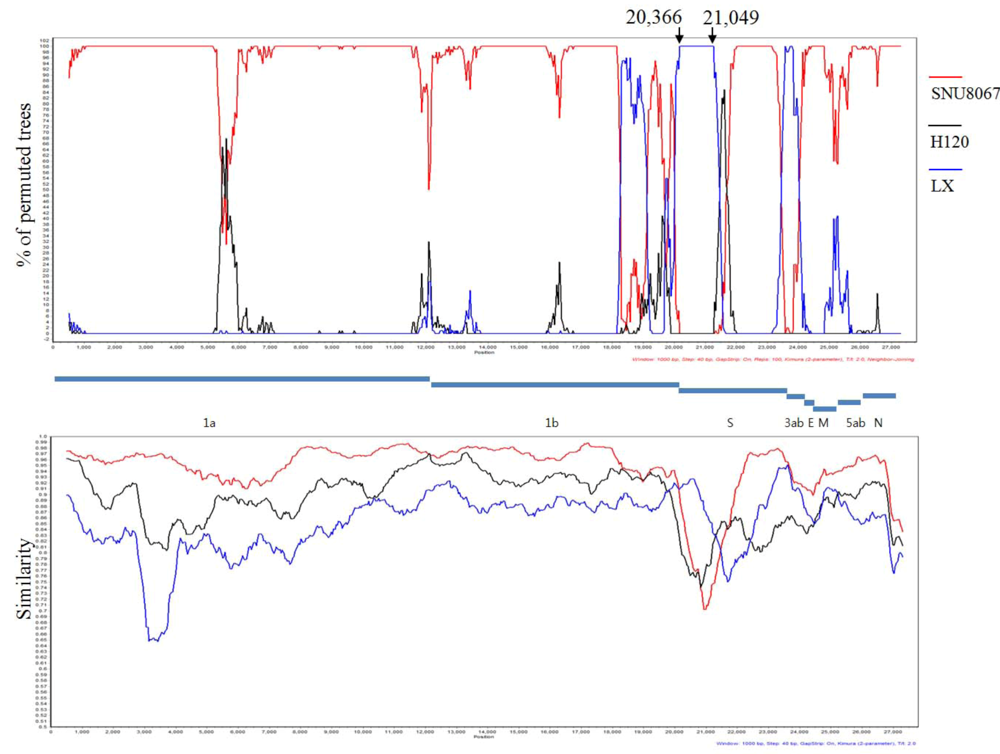
2.3. Computational Recombination Analysis
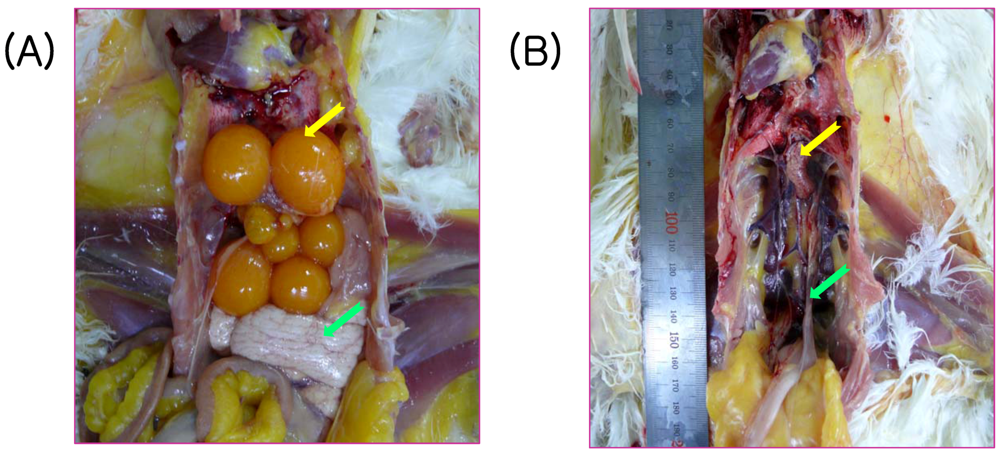
2.4. Vaccine Efficacy of a Commercial Inactivated Oil-Emulsion Vaccine against SNU8067


| Challenge group a | Number of chickens | Lesion score b | |||||||
| Follicle | Oviduct | ||||||||
| − | + | ++ | +++ | − | + | ++ | +++ | ||
| Vaccinated | 10 | 7c | 1 | 2 | 0 | 8 | 0 | 2 | 0 |
| Non-vaccinated | 10 | 2c | 2 | 1 | 5 | 5 | 3 | 2 | 0 |


3. Experimental Section
3.1. Virus, Eggs, and Chickens
3.2. Virus Titration
3.3. RNA Extraction and Reverse Transcription Polymerase Chain Reaction (RT-PCR)
3.4. Sequencing and Genome Sequence Analysis
| Primer | Length (bp) | Primer for PCR | |||
|---|---|---|---|---|---|
| Position a | Forward-5' | position | Reverse-3' | ||
| 1 | 1494 | 1–26 | ACTTAAGATAGATATTAATATATATC | 1476–1494 | TCAGCTTTTTGRTCAAGCA |
| 2 | 2474 | 1247–1265 | GGAACTTGTCTTGCAAGYA | 3702–3720 | CCACAAAAGTCTGCAATTG |
| 3 | 1595 | 3623–3642 | GCATTGGAYGARTTTAAAGA | 5198–5217 | TCAACAACTGATTTRATACC |
| 4 | 1457 | 5045–5064 | GYCAGTTTTGAYRATCTTAC | 6484–6501 | AGCYCRCCAGCAACTTCA |
| 5 | 1812 | 6298–6317 | GGATRTAACATGTGAAGTKT | 8090–8109 | TCATTAAAYAAMACCCATTG |
| 6 | 1361 | 8027–8046 | AGCTATTGTAGRGGTAGTGT | 9368–9387 | CCAGTGTGTAATGCATTAGG |
| 7 | 1458 | 9244–9263 | CCCTGTYACTATGCGYTCTA | 10682–10701 | GTTGTRCAYTTTACATCACT |
| 8 | 1586 | 10547–10567 | GATCAATATAGGTATATGTGT | 12113–12132 | GCTATRTGKGCTCTACAATA |
| 9 | 1540 | 11993–12012 | CAACCTTTAGGTAAYTGTGT | 13513–13532 | AARCAAGAMGTTCTAAGATC |
| 10 | 1549 | 13366–13385 | ATAGCTACTTGTGGCTATCA | 14897–14914 | GCTCCATAACAGCYACAG |
| 11 | 1688 | 14653–14672 | GCCAAACARGGTCTTGTWGC | 16321–16340 | TCACCTACATACACAACATA |
| 12 | 1391 | 16183–16202 | AATGACACAGGCAAAAAGTA | 17554–17573 | GCTYTAGAACCRCAAGAACA |
| 13 | 1463 | 17464–17486 | GTTTCAGATTGYGTAGTKTTTGT | 18907–18926 | CGCTTRTAACACTGTGTAGA |
| 14 | 1521 | 18683–18702 | GAAAYATTCGCACACTRCCA | 20184–20203 | TTGCGTGCAGHGTTTTTCCA |
| 15 | 1845 | 20035–20054 | ACAGAGACAAGTTGGCAYGA | 21860–21879 | CCWGAMACTACAAACTGYTG |
| 16 | 1799 | 21581–21599 | AAGAGYGRTGGCTCTCGTA | 23360–23379 | GTAACTAYATCTCCTGCAGT |
| 17 | 1607 | 23158–23177 | TGCACCTAATGGYATAGTGT | 24745–24764 | GTAYTATCGCTGCGACAAGA |
| 18 | 1735 | 24666–24685 | TGWTAGTGTTATGGWGCTTT | 26381–26400 | GGAATGAARTCCCAACGGAA |
| 19 | 1220 | 26256–26275 | ATGGTATAGTGTGGGTTGCT | 27456–27475 | TGSTCTAACTCTAWACTAGC |
3.5. Vaccine Efficacy Test for the Inactivated Oil-Emulsion Vaccine against SNU8067
3.6. Enzyme-Linked Immunosorbent Assay (ELISA)
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef]
- Lai, M.M.C. Recombination in large RNA viruses: Coronaviruses. Semin. Virol. 1996, 7, 381–388. [Google Scholar] [CrossRef]
- Cavanagh, D.; Naqi, S.A. Infectious Bronchitis, 11th ed; Iowa State Press: Ames, IA, USA, 2003. [Google Scholar]
- Gough, R.E.; Randall, C.J.; Dagless, M.; Alexander, D.J.; Cox, W.J.; Pearson, D. A 'new' strain of infectious bronchitis virus infecting domestic fowl in Great Britain. Vet. Rec. 1992, 130, 493–494. [Google Scholar]
- Albassam, M.A.; Winterfield, R.W.; Thacker, H.L. Comparison of the nephropathogenicity of four strains of infectious bronchitis virus. Avian Dis. 1986, 30, 468–476. [Google Scholar] [CrossRef]
- Zhou, J.Y.; Hong, J. Isolation and identification and pathogenicity of virus causing proventricular-type infectious bronchitis. Chin. J. Anim. Poultry Infect. Dis. 1998, 20, 62–65. [Google Scholar]
- Lee, E.-K.; Jeon, W.-J.; Lee, Y.-J.; Jeong, O.-M.; Choi, J.-G.; Kwon, J.-H.; Choi, K.-S. Genetic Diversity of Avian Infectious Bronchitis Virus Isolates in Korea Between 2003 and 2006. Avian Dis. 2008, 52, 332–337. [Google Scholar] [CrossRef]
- Lim, T.H.; Kim, M.S.; Jang, J.H.; Lee, D.H.; Park, J.K.; Youn, H.N.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Live attenuated nephropathogenic infectious bronchitis virus vaccine provides broad cross protection against new variant strains. Poultry Sci. 2012, 91, 89–94. [Google Scholar] [CrossRef]
- Song, C.S.; Lee, Y.J.; Kim, J.H.; Sung, H.W.; Lee, C.W.; Izumiya, Y.; Miyazawa, T.; Jang, H.K.; Mikami, T. Epidemiological classification of infectious bronchitis virus isolated in Korea between 1986 and 1997. Avian Pathol. 1998, 27, 409–416. [Google Scholar] [CrossRef]
- Choi, K.S.; Lee, E.K.; Jeon, W.J.; Park, M.J.; Kim, J.W.; Kwon, J.H. Pathogenicity and antigenicity of a new variant of Korean nephropathogenic infectious bronchitis virus. J. Vet. Sci. 2009, 10, 357–359. [Google Scholar] [CrossRef]
- Lim, T.-H.; Lee, H.-J.; Lee, D.-H.; Lee, Y.-N.; Park, J.-K.; Youn, H.-N.; Kim, M.-S.; Lee, J.-B.; Park, S.-Y.; Choi, I.-S.; et al. An emerging recombinant cluster of nephropathogenic strains of avian infectious bronchitis virus in Korea. Infect. Genet. Evol. 2011, 11, 678–685. [Google Scholar] [CrossRef]
- Hofstad, M.S. Cross-immunity in chickens using seven isolates of avian infectious bronchitis virus. Avian Dis. 1981, 25, 650–654. [Google Scholar] [CrossRef]
- Cook, J.K.; Smith, H.W.; Huggins, M.B. Infectious bronchitis immunity: Its study in chickens experimentally infected with mixtures of infectious bronchitis virus and Escherichia coli. J. Gen. Virol. 1986, 67, 1427–1434. [Google Scholar] [CrossRef]
- Klieve, A.V.; Cumming, R.B. Immunity and cross-protection to nephritis produced by Australian infectious bronchitis viruses used as vaccines. Avian Pathol. 1988, 17, 829–839. [Google Scholar] [CrossRef]
- Sevoian, M.; Levine, P.P. Effects of infectious bronchitis on the reproductive tracts, egg production, and egg quality of laying chickens. Avian Dis. 1957, 1, 136–164. [Google Scholar] [CrossRef]
- Martin, D.P.; Lemey, P.; Lott, M.; Moulton, V.; Posada, D.; Lefeuvre, P. RDP3: A flexible and fast computer program for analyzing recombination. Bioinformatics 2010, 26, 2462–2463. [Google Scholar]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J. Virol. 1999, 73, 152–160. [Google Scholar]
- Hamilton, M.A.; Russo, R.C.; Thurston, R.V. Trimmed Spearman-Karber method for estimating median lethal concentrations in toxicity bioassays. Environ. Sci. Tech. 1977, 11, 714–719. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Roberts, R.J.; Vincze, T.; Posfai, J.; Macelis, D. ChromasPro. Nucleic Acids Res. 2003, 31, 418–420. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Heath, L.; van der Walt, E.; Varsani, A.; Martin, D.P. Recombination patterns in aphthoviruses mirror those found in other picornaviruses. J. Virol. 2006, 80, 11827–11832. [Google Scholar] [CrossRef]
- SPSS, version 18.0; PASW Statistics for Windows; SPSS Inc.: Chicago, IL, USA, 2009.
- Kwon, H.J.; Lee, D.W.; Ahn, Y.K.; Yoon, J.W.; Kim, S.J. Presence of infectious bronchitis virus in Korea before 1986. Korean J. Vet. Res. 2001, 41, 59–65. [Google Scholar]
- Lee, E.K.; Jeon, W.J.; Lee, Y.J.; Jeong, O.M.; Choi, J.G.; Kwon, J.H.; Choi, K.S. Genetic diversity of avian infectious bronchitis virus isolates in Korea between 2003 and 2006. Avian Dis. 2008, 52, 332–337. [Google Scholar] [CrossRef]
- Lim, T.H.; Kim, M.S.; Jang, J.H.; Lee, D.H.; Park, J.K.; Youn, H.N.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Live attenuated nephropathogenic infectious bronchitis virus vaccine provides broad cross protection against new variant strains. Poult. Sci. 2012, 91, 89–94. [Google Scholar] [CrossRef]
- Lee, Y.O.; Kim, J.H.; Kim, J.H.; Mo, I.P.; Yoon, H.J.; Choi, S.H.; Nam, G.S. The first outbreak of infectious bronchitis in Korea. Korean J. Vet. Res. 1986, 26, 277–282. [Google Scholar]
- Cavanagh, D.; Davis, P.J.; Darbyshire, J.H.; Peters, R.W. Coronavirus IBV: Virus retaining spike glycopolypeptide S2 but not S1 is unable to induce virus-neutralizing or haemagglutination-inhibiting antibody, or induce chicken tracheal protection. J. Gen. Virol. 1986, 67, 1435–1442. [Google Scholar] [CrossRef]
- Gelb, J., Jr.; Keeler, C.L., Jr.; Nix, W.A.; Rosenberger, J.K.; Cloud, S.S. Antigenic and S-1 genomic characterization of the Delaware variant serotype of infectious bronchitis virus. Avian Dis. 1997, 41, 661–669. [Google Scholar] [CrossRef]
- Kant, A.; Koch, G.; van Roozelaar, D.J.; Kusters, J.G.; Poelwijk, F.A.; van der Zeijst, B.A. Location of antigenic sites defined by neutralizing monoclonal antibodies on the S1 avian infectious bronchitis virus glycopolypeptide. J. Gen. Virol. 1992, 73, 591–596. [Google Scholar] [CrossRef]
- Boots, A.M.; Benaissa-Trouw, B.J.; Hesselink, W.; Rijke, E.; Schrier, C.; Hensen, E.J. Induction of anti-viral immune responses by immunization with recombinant-DNA encoded avian coronavirus nucleocapsid protein. Vaccine 1992, 10, 119–124. [Google Scholar] [CrossRef]
- Timms, L.M.; Bracewell, C.D. Cell mediated and humoral immune response of chickens to inactivated oil-emulsion infectious bronchitis vaccine. Res. Vet. Sci. 1983, 34, 224–230. [Google Scholar]
- Cavanagh, D.; Davis, P.J.; Cook, J.K.; Li, D.; Kant, A.; Koch, G. Location of the amino acid differences in the S1 spike glycoprotein subunit of closely related serotypes of infectious bronchitis virus. Avian Pathol. 1992, 21, 33–43. [Google Scholar] [CrossRef]
- Kim, J.H. Serological Differentiation, Pathogenicity and Immunogenicity of Avian Infectious Bronchitis Virus Isolated in Korea; Seoul National University: Seoul, Korea, 1994. [Google Scholar]
- Hodgson, T.; Britton, P.; Cavanagh, D. Neither the RNA nor the proteins of open reading frames 3a and 3b of the coronavirus infectious bronchitis virus are essential for replication. J. Virol. 2006, 80, 296–305. [Google Scholar] [CrossRef]
- Casais, R.; Davies, M.; Cavanagh, D.; Britton, P. Gene 5 of the avian coronavirus infectious bronchitis virus is not essential for replication. J. Virol. 2005, 79, 8065–8078. [Google Scholar] [CrossRef]
- Shi, P.; Yu, L.; Fu, Y.X.; Huang, J.F.; Zhang, K.Q.; Zhang, Y.P. Evolutionary implications of Avian Infectious Bronchitis Virus (AIBV) analysis. Cell Res. 2006, 16, 323–327. [Google Scholar] [CrossRef]
- Tang, X.; Li, G.; Vasilakis, N.; Zhang, Y.; Shi, Z.; Zhong, Y.; Wang, L.F.; Zhang, S. Differential stepwise evolution of SARS coronavirus functional proteins in different host species. BMC Evol. Biol. 2009, 9, 52. [Google Scholar]
- Wang, L.; Junker, D.; Collisson, E.W. Evidence of natural recombination within the S1 gene of infectious bronchitis virus. Virology 1993, 192, 710–716. [Google Scholar] [CrossRef]
- Brooks, J.E.; Rainer, A.C.; Parr, R.L.; Woolcock, P.; Hoerr, F.; Collisson, E.W. Comparisons of envelope through 5B sequences of infectious bronchitis coronaviruses indicates recombination occurs in the envelope and membrane genes. Virus Res. 2004, 100, 191–198. [Google Scholar] [CrossRef]
- McKinley, E.T.; Jackwood, M.W.; Hilt, D.A.; Kissinger, J.C.; Robertson, J.S.; Lemke, C.; Paterson, A.H. Attenuated live vaccine usage affects accurate measures of virus diversity and mutation rates in avian coronavirus infectious bronchitis virus. Virus Res. 2011, 158, 225–234. [Google Scholar] [CrossRef]
- Crinion, R.A.; Ball, R.A.; Hofstad, M.S. Abnormalities in laying chickens following exposure to infectious bronchitis virus at one day old. Avian Dis. 1971, 15, 42–48. [Google Scholar] [CrossRef]
- Box, P.G.; Ellis, K.R. Infectious bronchitis in laying hens: Interference with response to emulsion vaccine by attenuated live vaccine. Avian Pathol. 1985, 14, 9–22. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hong, S.-M.; Kwon, H.-J.; Kim, I.-H.; Mo, M.-L.; Kim, J.-H. Comparative Genomics of Korean Infectious Bronchitis Viruses (IBVs) and an Animal Model to Evaluate Pathogenicity of IBVs to the Reproductive Organs. Viruses 2012, 4, 2670-2683. https://doi.org/10.3390/v4112670
Hong S-M, Kwon H-J, Kim I-H, Mo M-L, Kim J-H. Comparative Genomics of Korean Infectious Bronchitis Viruses (IBVs) and an Animal Model to Evaluate Pathogenicity of IBVs to the Reproductive Organs. Viruses. 2012; 4(11):2670-2683. https://doi.org/10.3390/v4112670
Chicago/Turabian StyleHong, Seung-Min, Hyuk-Joon Kwon, Il-Hwan Kim, Mei-Lan Mo, and Jae-Hong Kim. 2012. "Comparative Genomics of Korean Infectious Bronchitis Viruses (IBVs) and an Animal Model to Evaluate Pathogenicity of IBVs to the Reproductive Organs" Viruses 4, no. 11: 2670-2683. https://doi.org/10.3390/v4112670
APA StyleHong, S. -M., Kwon, H. -J., Kim, I. -H., Mo, M. -L., & Kim, J. -H. (2012). Comparative Genomics of Korean Infectious Bronchitis Viruses (IBVs) and an Animal Model to Evaluate Pathogenicity of IBVs to the Reproductive Organs. Viruses, 4(11), 2670-2683. https://doi.org/10.3390/v4112670