Identifying the Viral Genes Encoding Envelope Glycoproteins for Differentiation of Cyprinid herpesvirus 3 Isolates

Abstract
:1. Introduction
2. Results and Discussion
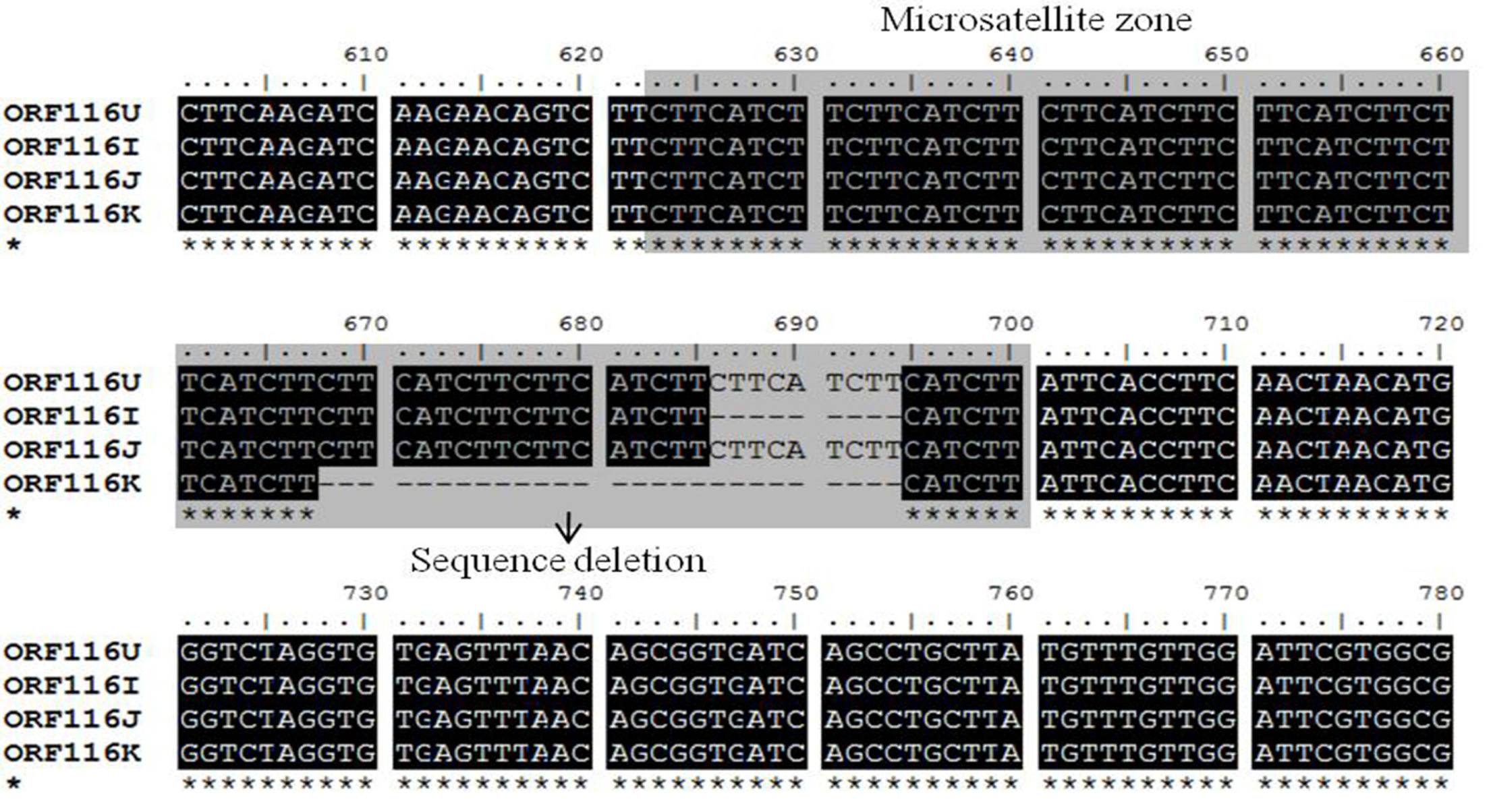
2.1. Analysis of CyHV-3 Envelope Glycoprotein Sequences



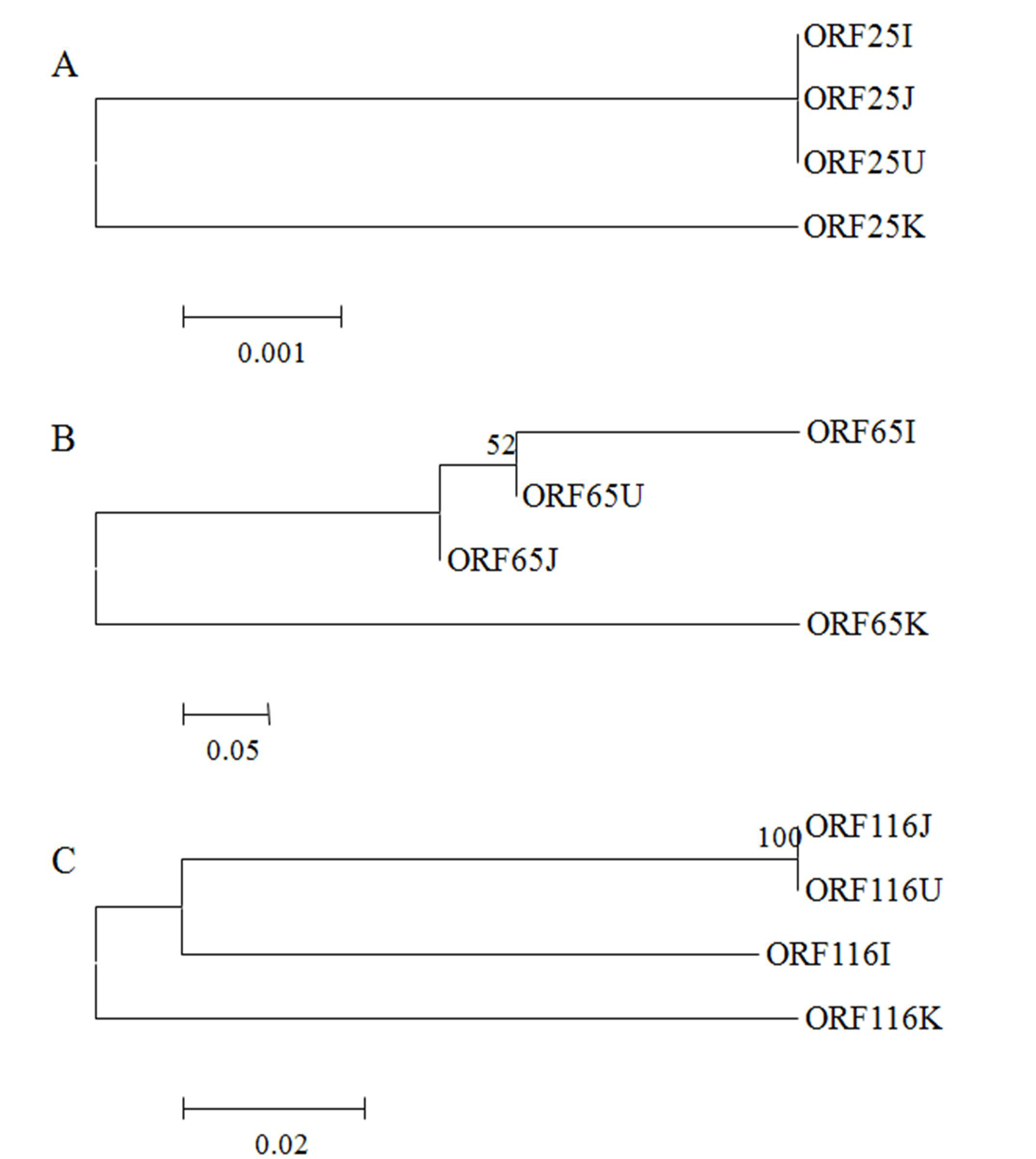
2.2. Phylogenetic Analysis of CyHV-3 Envelope Glycoprotein Genes

3. Experimental Section
3.1. Virus and Viral DNA
3.2. Amplification of Korean CyHV-3 Glycoprotein Genes
3.3. Analysis of Korean CyHV-3 Glycoprotein Genes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer name | Sequence (5’ → 3’ ) | Purpose |
|---|---|---|
| ORF25 | ||
| 25F1 | ATGACGGGTTGTGGGGTTTGG | Full length CDS |
| 25R1 | TTAGGGCCTCCGGGAAACCTG | |
| 25(m)F1 | GCGTCTCGGGAGATACTTTG | Internal region |
| 25(m)R1 | GGGCACTCCATCTCAAAGAC | |
| ORF65 | ||
| 65F1 | ATGGTCTCGCCGCTCGTCGTC | Full length CDS |
| 65R1 | CTACTTGATGGTCGCGGCGGC | |
| ORF116 | ||
| 116F1 | ATGAGACTTTTTCTCCTCGTC | Full length CDS |
| 116R1 | TCAAACTTTTGGTGATGAAAA |
3.4. Nucleotide Sequence Accession Numbers
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Whittington, R.; Chong, R. Global trade in ornamental fish from an Australian perspective: The case for revised import risk analysis and management strategies. Prev. Vet. Med. 2007, 81, 92–116. [Google Scholar]
- David, L.; Rothbard, S.; Rubinstein, I.; Katzman, H.; Hulata, G.; Hillel, J.; Lavi, U. Aspects of red and black color inheritance in the Japanese ornamental (Koi) carp (Cyprinus carpio L.). Aquaculture 2004, 233, 129–147. [Google Scholar] [CrossRef]
- Bretzinger, A.; Fischer-Scherl, T.; Oumouma, M.; Hoffmann, R.; Truyen, U. Mass mortalities in koi, Cyprinus carpio, associated with gill and skin disease. Bull. Eur. Assoc. Fish Pathol. 1999, 19, 182–185. [Google Scholar]
- Waltzek, T.B.; Kelley, G.O.; Stone, D.M.; Way, K.; Hanson, L.; Fukuda, H.; Hirono, I.; Aoki, T.; Davison, A.J.; Hedrick, R.P. Koi herpesvirus represents a third cyprinid herpesvirus (CyHV-3) in the family Herpesviridae. J. Gen. Virol. 2005, 86, 1659–1667. [Google Scholar]
- Davison, A.J. Herpesvirus systematics. Vet. Microbiol. 2010, 143, 52–69. [Google Scholar]
- Davison, A.J.; Eberle, R.; Hayward, G.S.; McGeoch, D.J.; Minson, A.C.; Pellett, P.E.; Roizman, B.; Studdert, M.J.; Thiry, E. Herpesviridae. In Virus Taxonomy: Eighth Report of the International Committee on Taxonomy of Viruses, 1st; Fauquet, C.M., Mayo, A.A., Maniloff, J., Desselberger, U., Ball, L.A., Eds.; Elsevier: Academic Press, Amsterdam, The Netherlands, 2005; pp. 193–212. [Google Scholar]
- Aoki, T.; Hirono, I.; Kurokawa, K.; Fukuda, H.; Nahary, R.; Eldar, A.; Davison, A.J.; Waltzek, T.B.; Bercovier, H.; Hedrick, R.P. Genome sequences of three koi herpesvirus isolates representing the expanding distribution of an emerging disease threatening koi and common carp worldwide. J. Virol. 2007, 81, 5058–5065. [Google Scholar]
- Davison, A.J.; Davison, M.D. Identification of structural proteins of channel catfish virus by mass spectrometry. Virology 1995, 206, 1035–1043. [Google Scholar]
- Kucuktas, H.; Brady, Y.J.; Tuzun, S. Cloning and expression of a putative glycoprotein gene of channel catfish virus using baculovirus expression system. Dis. Aquat. Organ. 1998, 34, 231–237. [Google Scholar]
- Dubin, G.; Socolof, E.; Frank, I.; Friedman, H. Herpes simplex virus type 1 Fc receptor protects infected cells from antibody-dependent cellular cytotoxicity. J. Virol. 1991, 65, 7046–7050. [Google Scholar]
- Dunowska, M.; Letchworth, G.J.; Collins, J.K.; DeMartini, J.C. Ovine herpesvirus-2 glycoprotein B sequences from tissues of ruminant malignant catarrhal fever cases and healthy sheep are highly conserved. J. Gen. Virol. 2001, 82, 2785–2790. [Google Scholar]
- Kaashoek, M.; Moerman, A.; Madic, J.; Rijsewijk, F.; Quak, J.; Gielkens, A.; Van Oirschot, J. A conventionally attenuated glycoprotein E-negative strain of bovine herpesvirus type 1 is an efficacious and safe vaccine. Vaccine 1994, 12, 439–444. [Google Scholar] [CrossRef]
- Davis, C.L.; Field, D.; Metzgar, D.; Saiz, R.; Morin, P.A.; Smith, I.L.; Spector, S.A.; Wills, C. Numerous length polymorphisms at short tandem repeats in human cytomegalovirus. J. Virol. 1999, 73, 6265–6270. [Google Scholar]
- Picone, O.; Ville, Y.; Costa, J.M.; Rouzioux, C.; Leruez-Ville, M. Human cytomegalovirus (HCMV) short tandem repeats analysis in congenital infection. J. Clin. Virol. 2005, 32, 254–256. [Google Scholar] [CrossRef]
- Deback, C.; Boutolleau, D.; Depienne, C.; Luyt, C.E.; Bonnafous, P.; Gautheret, A.; Garrigue, I.; Agut, H. Utilization of microsatellite polymorphism for differentiating herpes simplex virus type 1 strains. J. Clin. Microbiol. 2009, 47, 533–540. [Google Scholar]
- Segarra, A.; Pepin, J.F.; Arzul, I.; Morga, B.; Faury, N.; Renault, T. Detection and description of a particular Ostreid herpesvirus 1 genotype associated with massive mortality outbreaks of Pacific oysters, Crassostrea gigas, in France in 2008. Virus Res. 2010, 153, 92–99. [Google Scholar] [CrossRef]
- Gomez, D.K.; Joh, S.J.; Jang, H.; Shin, S.P.; Choresca, C.H., Jr.; Han, J.E.; Kim, J.H.; Jun, J.W.; Park, S.C. Detection of Koi Herpesvirus (KHV) from Koi (Cyprinus carpio koi) Broodstock in Korea. Aquaculture 2011, 311, 42–47. [Google Scholar] [CrossRef]
- Kado, C.I.; Liu, S.T. Rapid procedure for detection and isolation of large and small plasmids. J. Bacteriol. 1981, 145, 1365–1373. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.M.; Eyers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Han, J.E.; Kim, J.H.; Renault, T.; Jr., C.C.; Shin, S.P.; Jun, J.W.; Park, S.C. Identifying the Viral Genes Encoding Envelope Glycoproteins for Differentiation of Cyprinid herpesvirus 3 Isolates. Viruses 2013, 5, 568-576. https://doi.org/10.3390/v5020568
Han JE, Kim JH, Renault T, Jr. CC, Shin SP, Jun JW, Park SC. Identifying the Viral Genes Encoding Envelope Glycoproteins for Differentiation of Cyprinid herpesvirus 3 Isolates. Viruses. 2013; 5(2):568-576. https://doi.org/10.3390/v5020568
Chicago/Turabian StyleHan, Jee Eun, Ji Hyung Kim, Tristan Renault, Casiano Choresca Jr., Sang Phil Shin, Jin Woo Jun, and Se Chang Park. 2013. "Identifying the Viral Genes Encoding Envelope Glycoproteins for Differentiation of Cyprinid herpesvirus 3 Isolates" Viruses 5, no. 2: 568-576. https://doi.org/10.3390/v5020568
APA StyleHan, J. E., Kim, J. H., Renault, T., Jr., C. C., Shin, S. P., Jun, J. W., & Park, S. C. (2013). Identifying the Viral Genes Encoding Envelope Glycoproteins for Differentiation of Cyprinid herpesvirus 3 Isolates. Viruses, 5(2), 568-576. https://doi.org/10.3390/v5020568