HIV-1 Diversity in the Envelope Glycoproteins: Implications for Viral Entry Inhibition

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. HIV Entry and Its Inhibition

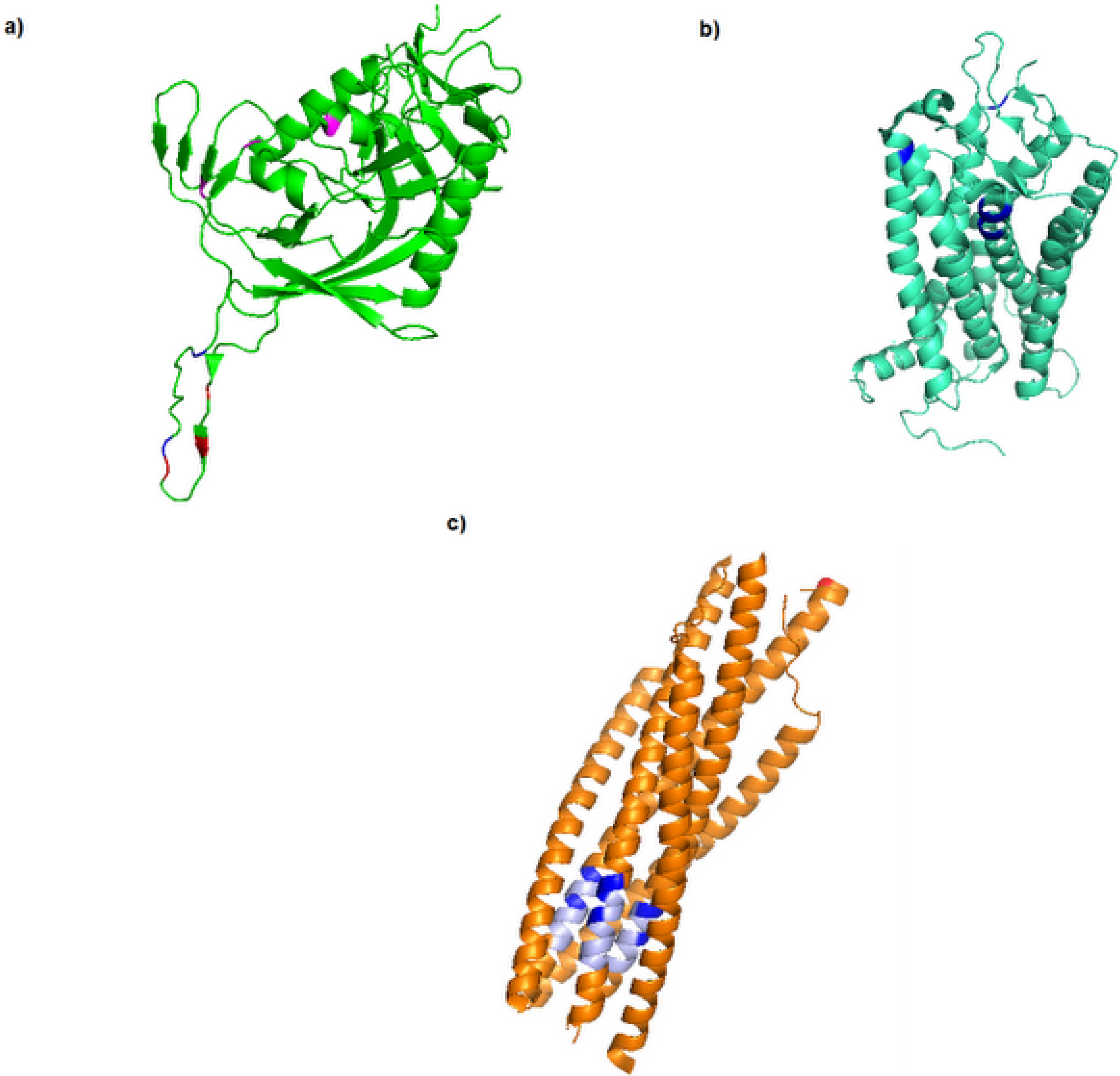
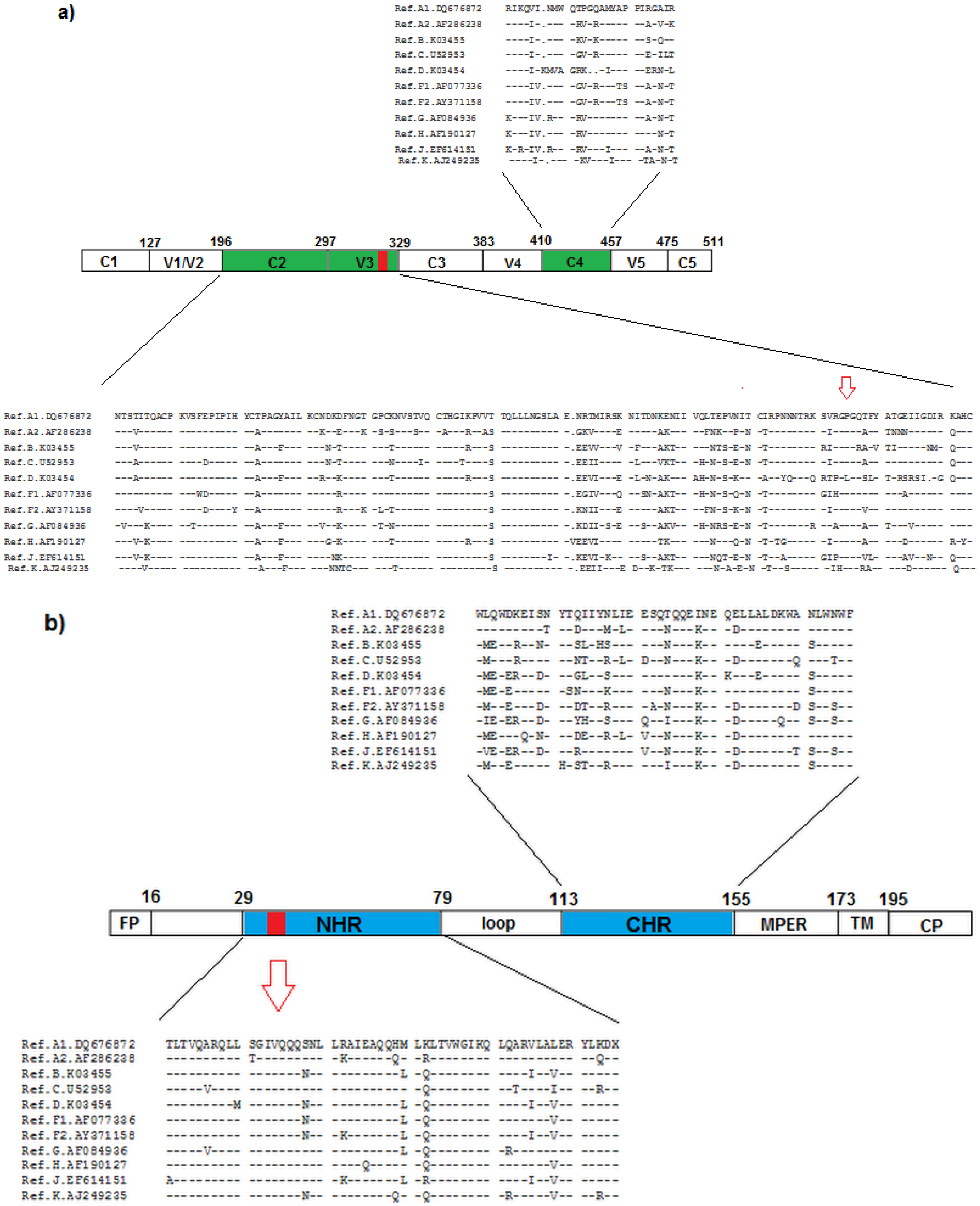
3. Diversity in the Envelope Glycoproteins

4. Resistance to Entry Inhibitors
Conflict of Interest
References
- Taylor, B.S.; Sobieszczyk, M.E.; McCutchan, F.E.; Hammer, S.M. The challenge of HIV-1 subtype diversity. N Engl. J. Med. 2008, 358, 1590–1602. [Google Scholar] [CrossRef]
- Sharp, P.M.; Hahn, B.H. Origins of HIV and the AIDS Pandemic. Cold Spring Harbor Laboratory Press 2011, 1, 1. [Google Scholar]
- Herbeck, J.T.; Nickle, D.C.; Learn, G.H.; Gottlieb, G.S.; Curlin, M.E.; Heath, L.; Mullins, J.I. Human immunodeficiency virus type 1 env evolves toward ancestral states upon transmission to a new host. J. Virol. 2006, 80, 1637–1644. [Google Scholar]
- Yang, Z.; Chakrabarti, B.K.; Xu, L.; Welcher, B.; Kong, W.; Leung, K.; Panet, A.; Mascola, J.R.; Nabel, G.J. Selective modification of variable loops alters tropism and enhances immunogenicity of human immunodeficiency virus type 1 envelope. J. Virol. 2004, 78, 4029–4036. [Google Scholar]
- Wainberg, M.A.; Brenner, B.G. Role of HIV subtype diversity in the development of resistance to antiviral drugs. Viruses 2010, 2, 2493–2508. [Google Scholar] [CrossRef]
- Wilen, C.B.; Tilton, J.C.; Doms, R.W. Molecular mechanisms of HIV entry. Viral Molecular Machines 2012, 726, 223–242. [Google Scholar] [CrossRef]
- Sundaravaradan, V.; Das, S.R.; Ramakrishnan, R.; Sehgal, S.; Gopalan, S.; Ahmad, N.; Jameel, S. Role of HIV-1 subtype C envelope V3 to V5 regions in viral entry, coreceptor utilization and replication efficiency in primary T-lymphocytes and monocyte-derived macrophages. Virol. J. 2007, 4, 126. [Google Scholar] [CrossRef]
- Caffrey, M. HIV envelope: Challenges and opportunities for development of entry inhibitors. Trends Microbiol. 2011, 19, 191–197. [Google Scholar] [CrossRef]
- Lobritz, M.A.; Ratcliff, A.N.; Arts, E.J. HIV-1 entry, inhibitors, and resistance. Viruses 2010, 2, 1069–1105. [Google Scholar] [CrossRef]
- Choisy, M.; Woelk, C.H.; Guégan, J.F.; Robertson, D.L. Comparative study of adaptive molecular evolution in different human immunodeficiency virus groups and subtypes. J. Virol. 2004, 78, 1962–1970. [Google Scholar] [CrossRef]
- Travers, S.A.A.; O'Connell, M.J.; McCormack, G.P.; McInerney, J.O. Evidence for heterogeneous selective pressures in the evolution of the env gene in different human immunodeficiency virus type 1 subtypes. J. Virol. 2005, 79, 1836–1841. [Google Scholar] [CrossRef]
- Klevytska, A.M.; Mracna, M.R.; Guay, L.; Becker-Pergola, G.; Furtado, M.; Zhang, L.; Jackson, J.B.; Eshleman, S.H. Analysis of length variation in the V1-V2 region of env in nonsubtype B HIV type 1 from Uganda. AIDS Res. Hum. Retrovir. 2002, 11, 791–796. [Google Scholar]
- Curlin, M.E.; Zioni, R.; Hawes, S.E.; Liu, Y.; Deng, W.; Gottlieb, G.S.; Zhu, T.; Mullins, J.I. HIV-1 envelope subregion length variation during disease progression. PLoS Pathogens 2010, 6, e1001228. [Google Scholar] [CrossRef]
- Cormier, E.G.; Dragic, T. The crown and stem of the V3 loop play distinct roles in human immunodeficiency virus type 1 envelope glycoprotein interactions with the CCR5 coreceptor. J. Virol. 2002, 76, 8953–8957. [Google Scholar] [CrossRef]
- Sander, O.; Sing, T.; Sommer, I.; Low, A.J.; Cheung, P.K.; Harrigan, P.R.; Lengauer, T.; Domingues, F.S. Structural descriptors of gp120 V3 loop for the prediction of HIV-1 coreceptor usage. PLoS Computational Biology 2007, 3, e58. [Google Scholar] [CrossRef]
- Gaschen, B.; Taylor, J.; Yusim, K.; Foley, B.; Gao, F.; Lang, D.; Novitsky, V.; Haynes, B.; Hahn, B.H.; Bhattacharya, T. Diversity considerations in HIV-1 vaccine selection. Science 2002, 296, 2354–2360. [Google Scholar] [CrossRef]
- Gnanakaran, S.; Lang, D.; Daniels, M.; Bhattacharya, T.; Derdeyn, C.A.; Korber, B. Clade-specific differences between human immunodeficiency virus type 1 clades B and C: Diversity and correlations in C3-V4 regions of gp120. J. Virol. 2007, 81, 4886–4891. [Google Scholar] [CrossRef]
- Rong, R.; Gnanakaran, S.; Decker, J.M.; Bibollet-Ruche, F.; Taylor, J.; Sfakianos, J.N.; Mokili, J.L.; Muldoon, M.; Mulenga, J.; Allen, S. Unique mutational patterns in the envelope α2 amphipathic helix and acquisition of length in gp120 hypervariable domains are associated with resistance to autologous neutralization of subtype C human immunodeficiency virus type 1. J. Virol. 2007, 81, 5658–5668. [Google Scholar]
- Patel, M.B.; Hoffman, N.G.; Swanstrom, R. Subtype-specific conformational differences within the V3 region of subtype B and subtype C human immunodeficiency virus type 1 Env proteins. J. Virol. 2008, 82, 903–916. [Google Scholar] [CrossRef]
- Holguín, A.; de Arellano, E.R.; Soriano, V. Amino acid conservation in the gp41 transmembrane protein and natural polymorphisms associated with enfuvirtide resistance across HIV-1 variants. AIDS Res. Hum. Retrovir. 2007, 23, 1067–1074. [Google Scholar] [CrossRef]
- Holguín, A.; Faudon, J.L.; Labernardière, J.L.; Soriano, V. Susceptibility of HIV-1 non-B subtypes and recombinant variants to Enfuvirtide. J. Clin. Virol. 2007, 38, 176–180. [Google Scholar] [CrossRef]
- Sanders, R.W.; Korber, B.; Lu, M.; Berkhout, B.; Moore, J.P. Mutational analyses and natural variablility of the gp41 ectodomain. HIV Sequence Compendium 2002, 2002, 43–68. [Google Scholar]
- Razzolini, F.; Vicenti, I.; Saladini, F.; Micheli, V.; Romano, L.; Cargnel, A.; Zazzi, M. Natural variability in the HR-1 and HR-2 domains of HIV type 1 gp41 from different clades circulating in Italy. AIDS Res. Hum. Retrovir. 2007, 23, 558–563. [Google Scholar] [CrossRef]
- Kuhmann, S.E.; Pugach, P.; Kunstman, K.J.; Taylor, J.; Stanfield, R.L.; Snyder, A.; Strizki, J.M.; Riley, J.; Baroudy, B.M.; Wilson, I.A. Genetic and phenotypic analyses of human immunodeficiency virus type 1 escape from a small-molecule CCR5 inhibitor. J. Virol. 2004, 78, 2790–2807. [Google Scholar]
- Ogert, R.A.; Wojcik, L.; Buontempo, C.; Ba, L.; Buontempo, P.; Ralston, R.; Strizki, J.; Howe, J.A. Mapping resistance to the CCR5 co-receptor antagonist vicriviroc using heterologous chimeric HIV-1 envelope genes reveals key determinants in the C2-V5 domain of gp120. Virology 2008, 373, 387–399. [Google Scholar] [CrossRef]
- Putcharoen, O.; Lee, S.H.; Henrich, T.J.; Hu, Z.; Vanichanan, J.; Coakley, E.; Greaves, W.; Gulick, R.M.; Kuritzkes, D.R.; Tsibris, A. HIV-1 Clinical isolates resistant to CCR5 antagonists exhibit delayed entry kinetics that are corrected in the presence of drug. Science Signaling 2012, 86, 1119. [Google Scholar]
- Anastassopoulou, C.G.; Ketas, T.J.; Sanders, R.W.; Klasse, P.J.; Moore, J.P. Effects of sequence changes in the HIV-1 gp41 fusion peptide on CCR5 inhibitor resistance. Virology 2012, 482, 86–97. [Google Scholar]
- Araújo, L.A.L.; Junqueira, D.M.; de Medeiros, R.M.; Matte, M.C.C.; Almeida, S.E.M. Naturally occurring resistance mutations to HIV-1 entry inhibitors in subtypes B, C, and CRF31_BC. J. Clin. Virol. 2012, 54, 6–10. [Google Scholar] [CrossRef]
- Gonzalez, S.; Gondwe, C.; Tully, D.C.; Minhas, V.; Shea, D.; Kankasa, C.; M'soka, T.; Wood, C. Short communication: Antiretroviral therapy resistance mutations present in the HIV type 1 subtype C pol and env regions from therapy-naive patients in Zambia. AIDS Res. Hum. Retrovir. 2010, 26, 795–803. [Google Scholar] [CrossRef]
- Geretti, A.M.; Easterbrook, P. Antiretroviral resistance in clinical practice. Int. J. STD AIDS 2006, 12, 145–153. [Google Scholar] [CrossRef]
- Ray, N.; Blackburn, L.A.; Doms, R.W. HR-2 mutations in human immunodeficiency virus type 1 gp41 restore fusion kinetics delayed by HR-1 mutations that cause clinical resistance to enfuvirtide. J. Virol. 2009, 83, 2989–2995. [Google Scholar] [CrossRef]
- Xu, L.; Pozniak, A.; Wildfire, A.; Stanfield-Oakley, S.A.; Mosier, S.M.; Ratcliffe, D.; Workman, J.; Joall, A.; Myers, R.; Smit, E. Emergence and evolution of enfuvirtide resistance following long-term therapy involves heptad repeat 2 mutations within gp41. Antimicrob. Agents Chemother. 2005, 49, 1113–1119. [Google Scholar] [CrossRef]
- Baatz, F.; Nijhuis, M.; Lemaire, M.; Riedijk, M.; Wensing, A.M.J.; Servais, J.Y.; van Ham, P.M.; Hoepelman, A.I.M.; Koopmans, P.P.; Sprenger, H.G. Impact of the HIV-1 env genetic context outside HR1–HR2 on resistance to the fusion inhibitor enfuvirtide and viral infectivity in clinical isolates. PloS One 2011, 6, e21535. [Google Scholar]
- Labrosse, B.; Morand-Joubert, L.; Goubard, A.; Rochas, S.; Labernardière, J.L.; Pacanowski, J.; Meynard, J.L.; Hance, A.J.; Clavel, F.; Mammano, F. Role of the envelope genetic context in the development of enfuvirtide resistance in human immunodeficiency virus type 1-infected patients. J. Virol. 2006, 80, 8807–8819. [Google Scholar]
- Reeves, J.D.; Gallo, S.A.; Ahmad, N.; Miamidian, J.L.; Harvey, P.E.; Sharron, M.; Pöhlmann, S.; Sfakianos, J.N.; Derdeyn, C.A.; Blumenthal, R. Sensitivity of HIV-1 to entry inhibitors correlates with envelope/coreceptor affinity, receptor density, and fusion kinetics. Proc. Natl. Acad. Sci. 2002, 99, 16249–16254. [Google Scholar]
- Derdeyn, C.A.; Decker, J.M.; Sfakianos, J.N.; Wu, X.; O'Brien, W.A.; Ratner, L.; Kappes, J.C.; Shaw, G.M.; Hunter, E. Sensitivity of human immunodeficiency virus type 1 to the fusion inhibitor T-20 is modulated by coreceptor specificity defined by the V3 loop of gp120. J. Virol. 2000, 74, 8358–8367. [Google Scholar]
- Derdeyn, C.A.; Decker, J.M.; Sfakianos, J.N.; Zhang, Z.; O'Brien, W.A.; Ratner, L.; Shaw, G.M.; Hunter, E. Sensitivity of human immunodeficiency virus type 1 to fusion inhibitors targeted to the gp41 first heptad repeat involves distinct regions of gp41 and is consistently modulated by gp120 interactions with the coreceptor. J. Virol. 2001, 75, 8605–8614. [Google Scholar] [CrossRef]
- Heil, M.L.; Decker, J.M.; Sfakianos, J.N.; Shaw, G.M.; Hunter, E.; Derdeyn, C.A. Determinants of human immunodeficiency virus type 1 baseline susceptibility to the fusion inhibitors enfuvirtide and T-649 reside outside the peptide interaction site. J. Virol. 2004, 78, 7582–7589. [Google Scholar] [CrossRef]
- Cilliers, T.; Patience, T.; Pillay, C.; Papathanasopoulos, M.; Morris, L. Sensitivity of HIV type 1 subtype C isolates to the entry inhibitor T-20. AIDS Res. Hum. Retrovir. 2004, 20, 477–482. [Google Scholar] [CrossRef]
- Yu, X.; Yuan, L.; Huang, Y.; Xu, W.; Fang, Z.; Liu, S.; Shao, Y.; Jiang, S.; Ma, L. Susceptibility of HIV-1 subtypes B′, CRF07_BC and CRF01_AE that are predominantly circulating in China to HIV-1 entry inhibitors. PloS One 2011, 6, e17605. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Araújo, L.A.L.; Almeida, S.E.M. HIV-1 Diversity in the Envelope Glycoproteins: Implications for Viral Entry Inhibition. Viruses 2013, 5, 595-604. https://doi.org/10.3390/v5020595
Araújo LAL, Almeida SEM. HIV-1 Diversity in the Envelope Glycoproteins: Implications for Viral Entry Inhibition. Viruses. 2013; 5(2):595-604. https://doi.org/10.3390/v5020595
Chicago/Turabian StyleAraújo, Leonardo Augusto Luvison, and Sabrina E. M. Almeida. 2013. "HIV-1 Diversity in the Envelope Glycoproteins: Implications for Viral Entry Inhibition" Viruses 5, no. 2: 595-604. https://doi.org/10.3390/v5020595
APA StyleAraújo, L. A. L., & Almeida, S. E. M. (2013). HIV-1 Diversity in the Envelope Glycoproteins: Implications for Viral Entry Inhibition. Viruses, 5(2), 595-604. https://doi.org/10.3390/v5020595