Baculovirus Nuclear Import: Open, Nuclear Pore Complex (NPC) Sesame

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Baculovirus: Classification, Structure and Life Cycle
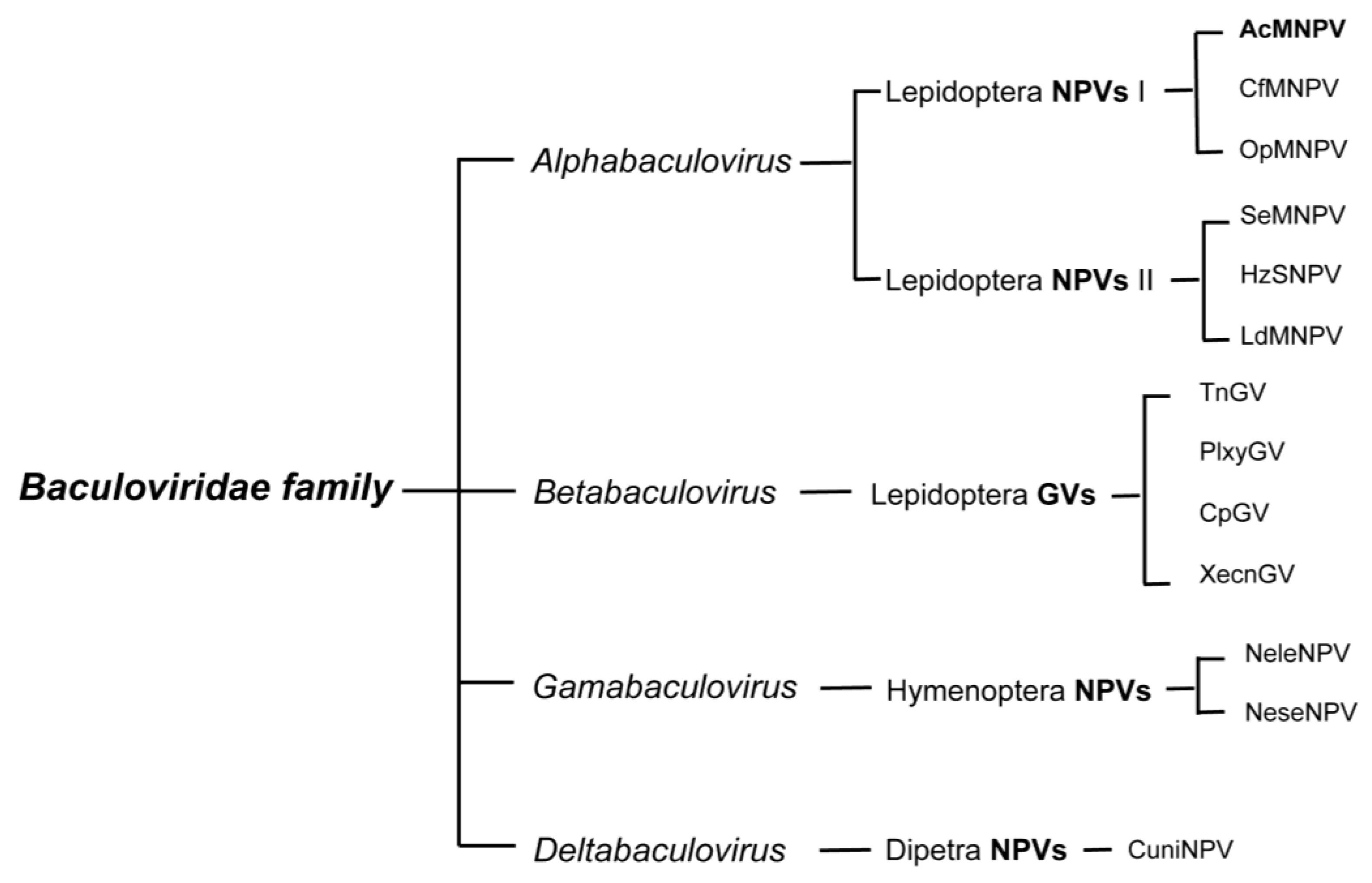
2.1. Baculovirus and Its Classification

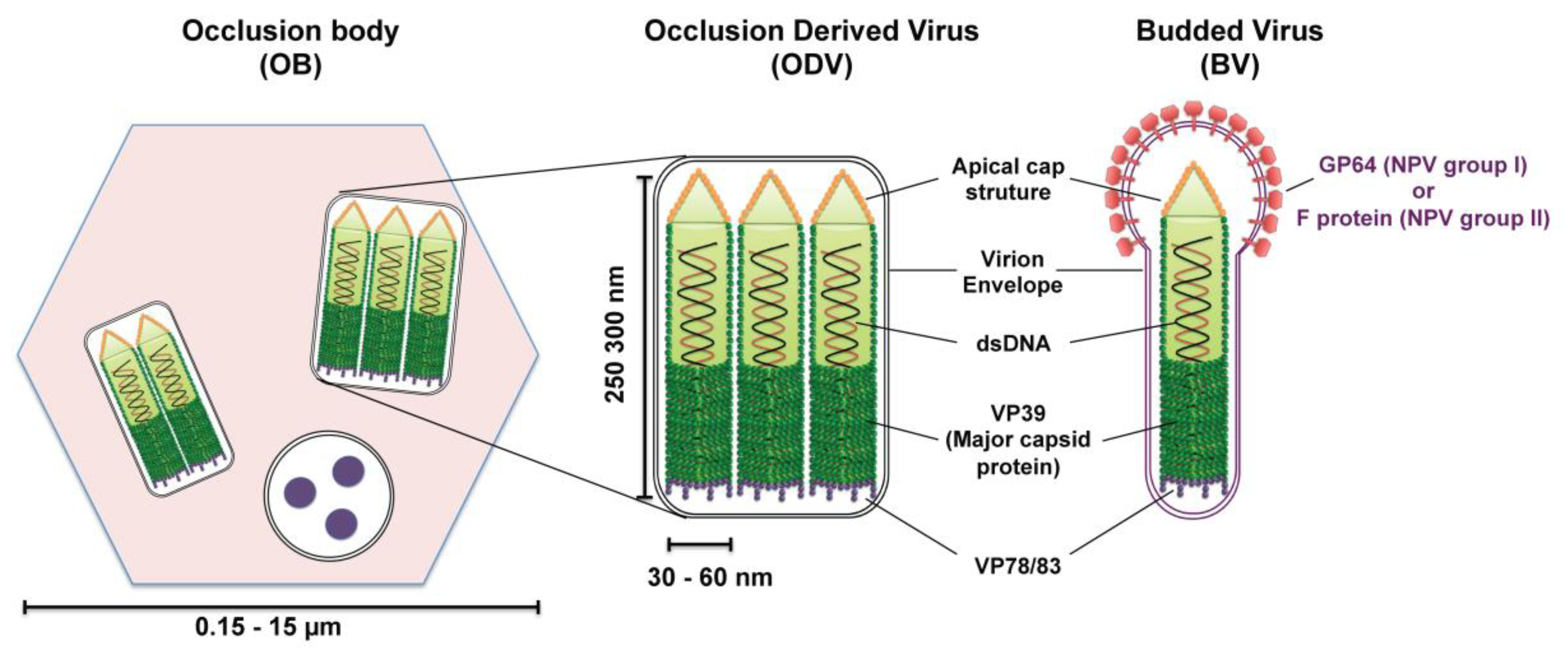
2.2. Baculovirus Structure

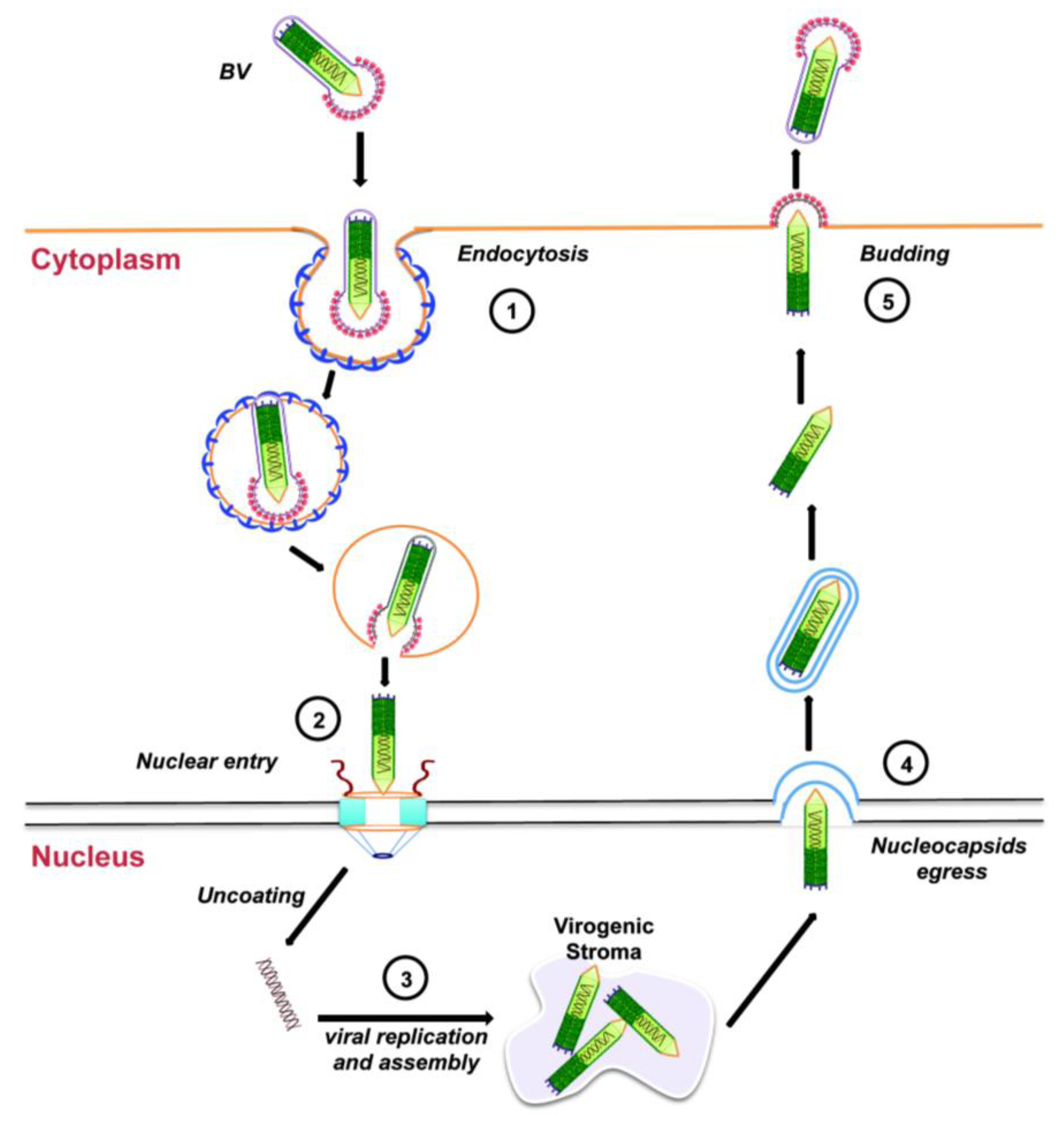
2.3. Budded Virus Infection Cycle

3. Baculovirus: Cellular Events before Nuclear Import
3.1. Cellular Entry of Budded Virus
3.2. Intracellular Trafficking towards the Nucleus
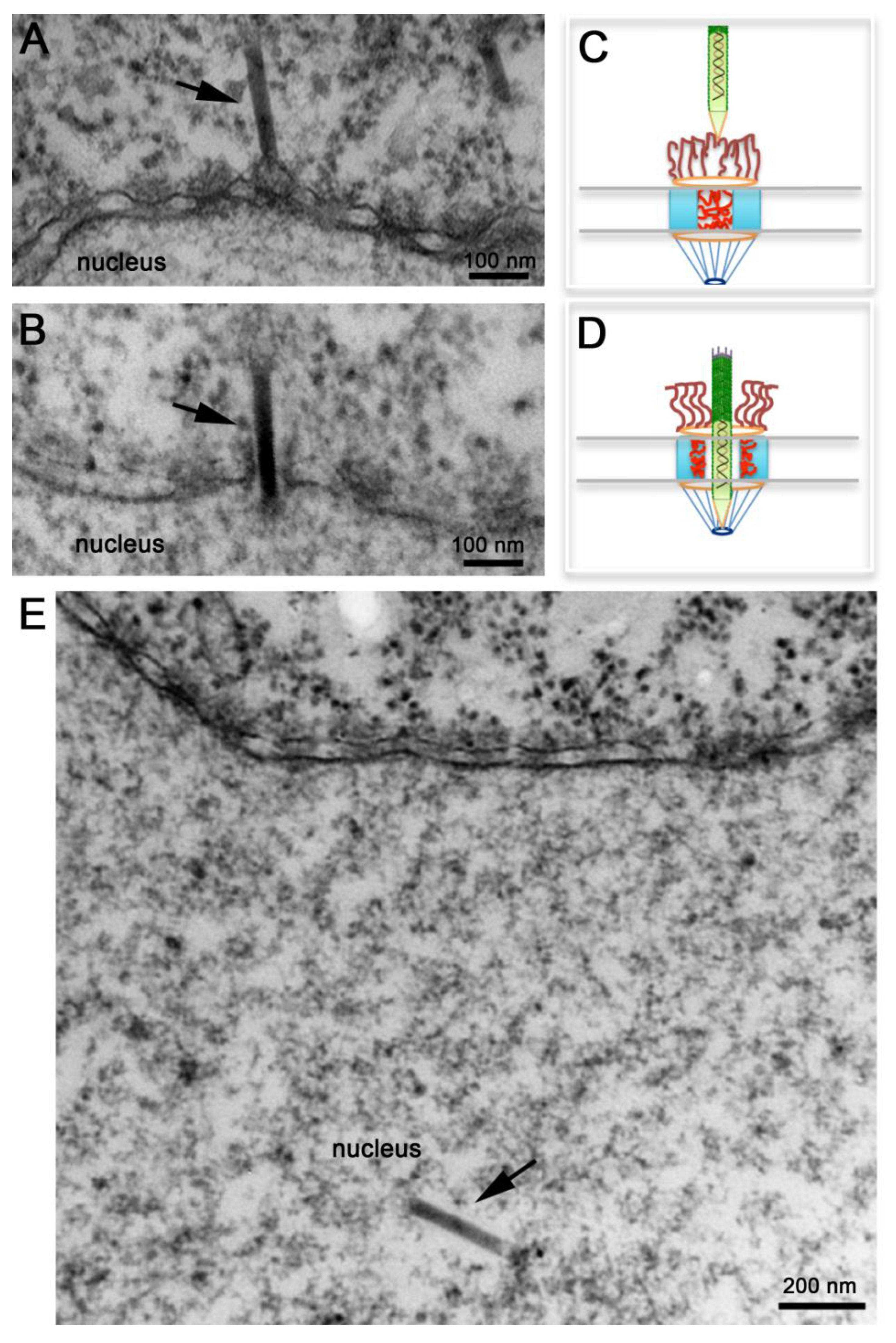
4. Nuclear Import of the Baculovirus Nucleocapsid
4.1. Early Studies from the 1960s to 1980s
4.2. Recent Advances in our Understanding of Baculovirus Nuclear Import Using AcMNPV
4.3. Using Baculovirus to Understand How the NPC Functions

5. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Van Beek, N.; Davis, D.C. Baculovirus insecticide production in insect larvae. Methods Mol. Biol. 2007, 388, 367–378. [Google Scholar] [CrossRef]
- Inceoglu, A.B.; Kamita, S.G.; Hinton, A.C.; Huang, Q.; Severson, T.F.; Kang, K.; Hammock, B.D. Recombinant baculoviruses for insect control. Pest. Manag. Sci. 2001, 57, 981–987. [Google Scholar] [CrossRef]
- Van Oers, M.M. Opportunities and challenges for the baculovirus expression system. J. Invertebr. Pathol. 2011, 107, S3–S15. [Google Scholar] [CrossRef]
- Hitchman, R.B.; Locanto, E.; Possee, R.D.; King, L.A. Optimizing the baculovirus expression vector system. Methods 2011, 55, 52–57. [Google Scholar] [CrossRef]
- Liu, C.Y.; Chen, H.Z.; Chao, Y.C. Maximizing baculovirus-mediated foreign proteins expression in mammalian cells. Curr. Gene. Ther. 2010, 10, 232–241. [Google Scholar] [CrossRef]
- Summers, M.D. Milestones leading to the genetic engineering of baculoviruses as expression vector systems and viral pesticides. Adv. Virus Res. 2006, 68, 3–73. [Google Scholar] [CrossRef]
- Cox, M.M. Recombinant protein vaccines produced in insect cells. Vaccine 2012, 30, 1759–1766. [Google Scholar] [CrossRef]
- Rychlowska, M.; Gromadzka, B.; Bienkowska-Szewczyk, K.; Szewczyk, B. Application of baculovirus-insect cell expression system for human therapy. Curr. Pharm. Biotechnol. 2011, 12, 1840–1849. [Google Scholar] [CrossRef]
- Rivera-Gonzalez, G.C.; Swift, S.L.; Dussupt, V.; Georgopoulos, L.J.; Maitland, N.J. Baculoviruses as gene therapy vectors for human prostate cancer. J. Invertebr. Pathol. 2011, 107, S59–S70. [Google Scholar] [CrossRef]
- Hitchman, R.B.; Murguia-Meca, F.; Locanto, E.; Danquah, J.; King, L.A. Baculovirus as vectors for human cells and applications in organ transplantation. J. Invertebr. Pathol. 2011, 107, S49–S58. [Google Scholar] [CrossRef]
- Airenne, K.J.; Hu, Y.C.; Kost, T.A.; Smith, R.H.; Kotin, R.M.; Ono, C.; Matsuura, Y.; Wang, S.; Yla-Herttuala, S. Baculovirus: An insect-derived vector for diverse gene transfer applications. Mol. Ther. 2013, 21, 739–749. [Google Scholar] [CrossRef]
- Hu, Y.C. Baculovirus vectors for gene therapy. Adv. Virus Res. 2006, 68, 287–320. [Google Scholar] [CrossRef]
- Friesen, P.D. Insect viruses. In Field’s Virology, 5th ed.; Knipe, D.M., Howley, P.M., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Straus, S.E., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007. [Google Scholar]
- Herniou, E.A.; Arif, B.M.; Becnel, J.J.; Blissard, G.W.; Bonning, B.C.; Harrison, R.; Jehle, J.A.; Theilmann, D.A.; Vlak, J.M. Baculoviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: New York, NY, USA, 2011. [Google Scholar]
- Jehle, J.A.; Blissard, G.W.; Bonning, B.C.; Cory, J.S.; Herniou, E.A.; Rohrmann, G.F.; Theilmann, D.A.; Thiem, S.M.; Vlak, J.M. On the classification and nomenclature of baculoviruses: A proposal for revision. Arch. Virol. 2006, 151, 1257–1266. [Google Scholar] [CrossRef]
- Pearson, M.N.; Rohrmann, G.F. Transfer, incorporation, and substitution of envelope fusion proteins among members of the Baculoviridae, Orthomyxoviridae, and Metaviridae (insect retrovirus) families. J. Virol. 2002, 76, 5301–5304. [Google Scholar] [CrossRef]
- Rohrmann, G.F. Baculovirus Molecular Biology, 2nd ed.National Center for Biotechnology Information (NCBI): Bethesda, MD, USA, 2011. Available online: http://www.ncbi.nlm.nih.gov/ bookshelf/br.fcgi?book=bacvir/ (accessed on 1 May 2013).
- Wang, R.; Deng, F.; Hou, D.; Zhao, Y.; Guo, L.; Wang, H.; Hu, Z. Proteomics of the Autographa californica nucleopolyhedrovirus budded virions. J. Virol. 2010, 84, 7233–7242. [Google Scholar] [CrossRef]
- Braunagel, S.C.; Russell, W.K.; Rosas-Acosta, G.; Russell, D.H.; Summers, M.D. Determination of the protein composition of the occlusion-derived virus of Autographa californica nucleopolyhedrovirus. Proc. Natl. Acad. Sci. USA 2003, 100, 9797–9802. [Google Scholar]
- Hou, D.; Zhang, L.; Deng, F.; Fang, W.; Wang, R.; Liu, X.; Guo, L.; Rayner, S.; Chen, H.W.; Hu, Z. Comparative proteomics reveal fundamental structural and functional differences between the two progeny phenotypes of a baculovirus. J. Virol. 2013, 87, 829–839. [Google Scholar] [CrossRef]
- Zhou, J.; Blissard, G.W. Identification of a GP64 subdomain involved in receptor binding by budded virions of the baculovirus Autographa californica multicapsid nucleopolyhedrovirus. J. Virol. 2008, 82, 4449–4460. [Google Scholar] [CrossRef]
- Hefferon, K.L.; Oomens, A.G.; Monsma, S.A.; Finnerty, C.M.; Blissard, G.W. Host cell receptor binding by baculovirus GP64 and kinetics of virion entry. Virology 1999, 258, 455–468. [Google Scholar] [CrossRef]
- Oomens, A.G.; Blissard, G.W. Requirement for GP64 to drive efficient budding of Autographa californica multicapsid nucleopolyhedrovirus. Virology 1999, 254, 297–314. [Google Scholar] [CrossRef]
- Long, G.; Pan, X.; Kormelink, R.; Vlak, J.M. Functional entry of baculovirus into insect and mammalian cells is dependent on clathrin-mediated endocytosis. J. Virol. 2006, 80, 8830–8833. [Google Scholar] [CrossRef]
- Matilainen, H.; Rinne, J.; Gilbert, L.; Marjomaki, V.; Reunanen, H.; Oker-Blom, C. Baculovirus entry into human hepatoma cells. J. Virol. 2005, 79, 15452–15459. [Google Scholar] [CrossRef]
- Laakkonen, J.P.; Makela, A.R.; Kakkonen, E.; Turkki, P.; Kukkonen, S.; Peranen, J.; Yla-Herttuala, S.; Airenne, K.J.; Oker-Blom, C.; Vihinen-Ranta, M.; et al. Clathrin-independent entry of baculovirus triggers uptake of E. coli in non-phagocytic human cells. PLoS One 2009, 4, e5093. [Google Scholar] [CrossRef]
- Blissard, G.W.; Wenz, J.R. Baculovirus GP64 envelope glycoprotein is sufficient to mediate pH-dependent membrane fusion. J. Virol. 1992, 66, 6829–6835. [Google Scholar]
- Dong, S.; Wang, M.; Qiu, Z.; Deng, F.; Vlak, J.M.; Hu, Z.; Wang, H. Autographa californica multicapsid nucleopolyhedrovirus efficiently infects Sf9 cells and transduces mammalian cells via direct fusion with the plasma membrane at low pH. J. Virol. 2010, 84, 5351–5359. [Google Scholar] [CrossRef]
- Kataoka, C.; Kaname, Y.; Taguwa, S.; Abe, T.; Fukuhara, T.; Tani, H.; Moriishi, K.; Matsuura, Y. Baculovirus GP64-mediated entry into mammalian cells. J. Virol. 2012, 86, 2610–2620. [Google Scholar] [CrossRef]
- Ward, B.M. The taking of the cytoskeleton one two three: How viruses utilize the cytoskeleton during egress. Virology 2011, 411, 244–250. [Google Scholar] [CrossRef]
- Greber, U.F.; Way, M. A superhighway to virus infection. Cell 2006, 124, 741–754. [Google Scholar] [CrossRef] [Green Version]
- Radtke, K.; Dohner, K.; Sodeik, B. Viral interactions with the cytoskeleton: A hitchhiker’s guide to the cell. Cell Microbiol. 2006, 8, 387–400. [Google Scholar] [CrossRef]
- Xu, H.; Yao, L.; Lu, S.; Qi, Y. Host filamentous actin is associated with Heliothis armigera single nucleopolyhedrosis virus (HaSNPV) nucleocapsid transport to the host nucleus. Curr. Microbiol. 2007, 54, 199–206. [Google Scholar] [CrossRef]
- Ohkawa, T.; Volkman, L.E.; Welch, M.D. Actin-based motility drives baculovirus transit to the nucleus and cell surface. J. Cell Biol. 2010, 190, 187–195. [Google Scholar] [CrossRef]
- Charlton, C.A.; Volkman, L.E. Penetration of Autographa californica nuclear polyhedrosis virus nucleocapsids into IPLB Sf 21 cells induces actin cable formation. Virology 1993, 197, 245–254. [Google Scholar] [CrossRef]
- Charlton, C.A.; Volkman, L.E. Sequential rearrangement and nuclear polymerization of actin in baculovirus-infected Spodoptera frugiperda cells. J. Virol. 1991, 65, 1219–1227. [Google Scholar]
- Lanier, L.M.; Volkman, L.E. Actin binding and nucleation by Autographa california M nucleopolyhedrovirus. Virology 1998, 243, 167–177. [Google Scholar] [CrossRef]
- Machesky, L.M.; Insall, R.H.; Volkman, L.E. WASP homology sequences in baculoviruses. Trends Cell Biol. 2001, 11, 286–287. [Google Scholar] [CrossRef]
- Goley, E.D.; Welch, M.D. The Arp2/3 complex: An actin nucleator comes of age. Nat. Rev. Mol. Cell Biol. 2006, 7, 713–726. [Google Scholar] [CrossRef]
- van Loo, N.D.; Fortunati, E.; Ehlert, E.; Rabelink, M.; Grosveld, F.; Scholte, B.J. Baculovirus infection of nondividing mammalian cells: Mechanisms of entry and nuclear transport of capsids. J. Virol. 2001, 75, 961–970. [Google Scholar]
- Goley, E.D.; Ohkawa, T.; Mancuso, J.; Woodruff, J.B.; D'Alessio, J.A.; Cande, W.Z.; Volkman, L.E.; Welch, M.D. Dynamic nuclear actin assembly by Arp2/3 complex and a baculovirus WASP-like protein. Science 2006, 314, 464–467. [Google Scholar] [CrossRef]
- Cohen, S.; Au, S.; Pante, N. How viruses access the nucleus. Biochim. Biophys. Acta 2011, 1813, 1634–1645. [Google Scholar] [CrossRef]
- Au, V.; Yu, M.; Carstens, E.B. Characterization of a baculovirus nuclear localization signal domain in the late expression factor 3 protein. Virology 2009, 385, 209–217. [Google Scholar] [CrossRef]
- Chen, Z.; Carsten, E.B. Identification of domains in Autographa californica multiple nucleopolyhedrovirus late expression factor 3 required for nuclear transport of P143. J. Virol. 2005, 79, 10915–10922. [Google Scholar] [CrossRef]
- Marek, M.; Merten, O.W.; Galibert, L.; Vlak, J.M.; van Oers, M.M. Baculovirus VP80 protein and the F-actin cytoskeleton interact and connect the viral replication factory with the nuclear periphery. J. Virol. 2011, 85, 5350–5362. [Google Scholar] [CrossRef]
- Summers, M.D. Apparent in vivo pathway of granulosis virus invasion and infection. J. Virol. 1969, 4, 188–190. [Google Scholar]
- Summers, M.D. Electron microscopic observations on granulosis virus entry, uncoating and replication processes during infection of the midgut cells of trichoplusia ni. J. Ultrastruct. Res. 1971, 35, 606–625. [Google Scholar]
- Sodeik, B.; Ebersold, M.W.; Helenius, A. Microtubule-mediated transport of incoming herpes simplex virus 1 capsids to the nucleus. J. Cell Biol. 1997, 136, 1007–1021. [Google Scholar] [CrossRef]
- Ojala, P.M.; Sodeik, B.; Ebersold, M.W.; Kutay, U.; Helenius, A. Herpes simplex virus type 1 entry into host cells: Reconstitution of capsid binding and uncoating at the nuclear pore complex in vitro. Mol. Cell Biol. 2000, 20, 4922–4931. [Google Scholar] [CrossRef]
- Raghow, R.; Grace, T.D. Studies on a nuclear polyhedrosis virus in bombyx mori cells in vitro. 1. Multiplication kinetics and ultrastructural studies. J. Ultrastruct. Res. 1974, 47, 384–399. [Google Scholar] [CrossRef]
- Wang, X.; Kelly, D.C. Baculovirus replicaton: Uptake of trichoplusia ni nuclear polyhedrosis virus particles by insect cells. J. Gen. Virol. 1984, 66, 541–550. [Google Scholar] [CrossRef]
- Kawanishi, C.Y.; Summers, M.D.; Stoltz, D.B.; Arnott, H.J. Entry of an insect virus in vivo by fusion of viral envelope and microvillus membrane. J. Invertebr. Pathol. 1972, 20, 104–108. [Google Scholar] [CrossRef]
- Carstens, E.B.; Tjia, S.T.; Doerfler, W. Infection of Spodoptera frugiperda cells with Autographa californica nuclear polyhedrosis virus I. Synthesis of intracellular proteins after virus infection. Virology 1979, 99, 386–398. [Google Scholar] [CrossRef]
- Knudson, D.L.; Harrap, K.A. Replication of a nuclear polyhedrosis virus in a continuous cell culture of Spodoptera frugiperda: Microscopy study of the sequence of events of the virus infection. J. Virol. 1976, 17, 254–268. [Google Scholar]
- Bassemir, U.; Miltenburger, H.G.; David, P. Morphogenesis of nuclear polyhedrosis virus from Autographa californica in a cell line from Mamestra brassicae (cabbage moth). Further aspects on baculovirus assembly. Cell Tissue Res. 1983, 228, 587–595. [Google Scholar]
- Hirumi, H.; Hirumi, K.; McIntosh, A.H. Morphogenesis of a nuclear polyhedrosis virus of the alfalfa looper in a continuous cabbage looper cell line. Ann. N. Y. Acad. Sci. 1975, 226, 302–326. [Google Scholar] [CrossRef]
- Granados, R.R. Early events in the infection of Heliothis zea midgut cells by baculovirus invasion and infection. Virology 1978, 90, 170–174. [Google Scholar] [CrossRef]
- Granados, R.R.; Lawler, K.A. In vivo pathway of Autographa californica baculovirus invasion and infection. Virology 1981, 108, 297–308. [Google Scholar] [CrossRef]
- Pante, N.; Kann, M. Nuclear pore complex is able to transport macromolecules with diameters of about 39 nm. Mol. Biol. Cell 2002, 13, 425–434. [Google Scholar] [CrossRef]
- Cohen, S.; Etingov, I.; Pante, N. Effect of viral infection on the nuclear envelope and nuclear pore complex. Int. Rev. Cell Mol. Biol. 2012, 299, 117–159. [Google Scholar] [CrossRef]
- Gorlich, D.; Pante, N.; Kutay, U.; Aebi, U.; Bischoff, F.R. Identification of different roles for RanGDP and RanGTP in nuclear protein import. EMBO J. 1996, 15, 5584–5594. [Google Scholar]
- Au, S.; Pante, N. Nuclear transport of baculovirus: Revealing the nuclear pore complex passage. J. Struct. Biol. 2012, 177, 90–98. [Google Scholar] [CrossRef]
- Pante, N. Use of intact Xenopus oocytes in nucleocytoplasmic transport studies. Methods Mol. Biol. 2006, 322, 301–314. [Google Scholar] [CrossRef]
- Au, S.; Cohen, S.; Pante, N. Microinjection of Xenopus laevis oocytes as a system for studying nuclear transport of viruses. Methods 2010, 51, 114–120. [Google Scholar] [CrossRef]
- Grossman, E.; Medalia, O.; Zwerger, M. Functional architecture of the nuclear pore complex. Annu. Rev. Biophys. 2012, 41, 557–584. [Google Scholar] [CrossRef]
- Pante, N.; Aebi, U. Sequential binding of import ligands to distinct nucleopore regions during their nuclear import. Science 1996, 273, 1729–1732. [Google Scholar] [CrossRef]
- Wente, S.R.; Rout, M.P. The nuclear pore complex and nuclear transport. Cold Spring Harb. Perspect. Biol. 2010, 2, a000562. [Google Scholar] [CrossRef]
- Ribbeck, K.; Gorlich, D. The permeability barrier of nuclear pore complexes appears to operate via hydrophobic exclusion. EMBO J. 2002, 21, 2664–2671. [Google Scholar] [CrossRef]
- Frey, S.; Richter, R.P.; Gorlich, D. FG-rich repeats of nuclear pore proteins form a three-dimensional meshwork with hydrogel-like properties. Science 2006, 314, 815–817. [Google Scholar] [CrossRef]
- Lim, R.Y.; Huang, N.P.; Koser, J.; Deng, J.; Lau, K.H.; Schwarz-Herion, K.; Fahrenkrog, B.; Aebi, U. Flexible phenylalanine-glycine nucleoporins as entropic barriers to nucleocytoplasmic transport. Proc. Natl. Acad. Sci. USA 2006, 103, 9512–9517. [Google Scholar]
- Lim, R.Y.; Fahrenkrog, B.; Koser, J.; Schwarz-Herion, K.; Deng, J.; Aebi, U. Nanomechanical basis of selective gating by the nuclear pore complex. Science 2007, 318, 640–643. [Google Scholar] [CrossRef]
- Lim, R.Y.; Koser, J.; Huang, N.P.; Schwarz-Herion, K.; Aebi, U. Nanomechanical interactions of phenylalanine-glycine nucleoporins studied by single molecule force-volume spectroscopy. J. Struct. Biol. 2007, 159, 277–289. [Google Scholar] [CrossRef]
- Yamada, J.; Phillips, J.L.; Patel, S.; Goldfien, G.; Calestagne-Morelli, A.; Huang, H.; Reza, R.; Acheson, J.; Krishnan, V.V.; Newsam, S.; et al. A bimodal distribution of two distinct categories of intrinsically disordered structures with separate functions in FG nucleoporins. Mol. Cell. Proteomics 2010, 9, 2205–2224. [Google Scholar] [CrossRef]
- Akey, C.W. Visualization of transport-related configurations of the nuclear pore transporter. Biophys. J. 1990, 58, 341–355. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Au, S.; Wu, W.; Panté, N. Baculovirus Nuclear Import: Open, Nuclear Pore Complex (NPC) Sesame. Viruses 2013, 5, 1885-1900. https://doi.org/10.3390/v5071885
Au S, Wu W, Panté N. Baculovirus Nuclear Import: Open, Nuclear Pore Complex (NPC) Sesame. Viruses. 2013; 5(7):1885-1900. https://doi.org/10.3390/v5071885
Chicago/Turabian StyleAu, Shelly, Wei Wu, and Nelly Panté. 2013. "Baculovirus Nuclear Import: Open, Nuclear Pore Complex (NPC) Sesame" Viruses 5, no. 7: 1885-1900. https://doi.org/10.3390/v5071885
APA StyleAu, S., Wu, W., & Panté, N. (2013). Baculovirus Nuclear Import: Open, Nuclear Pore Complex (NPC) Sesame. Viruses, 5(7), 1885-1900. https://doi.org/10.3390/v5071885