Epidemiology and Transmission of Kaposi’s Sarcoma-Associated Herpesvirus

Abstract
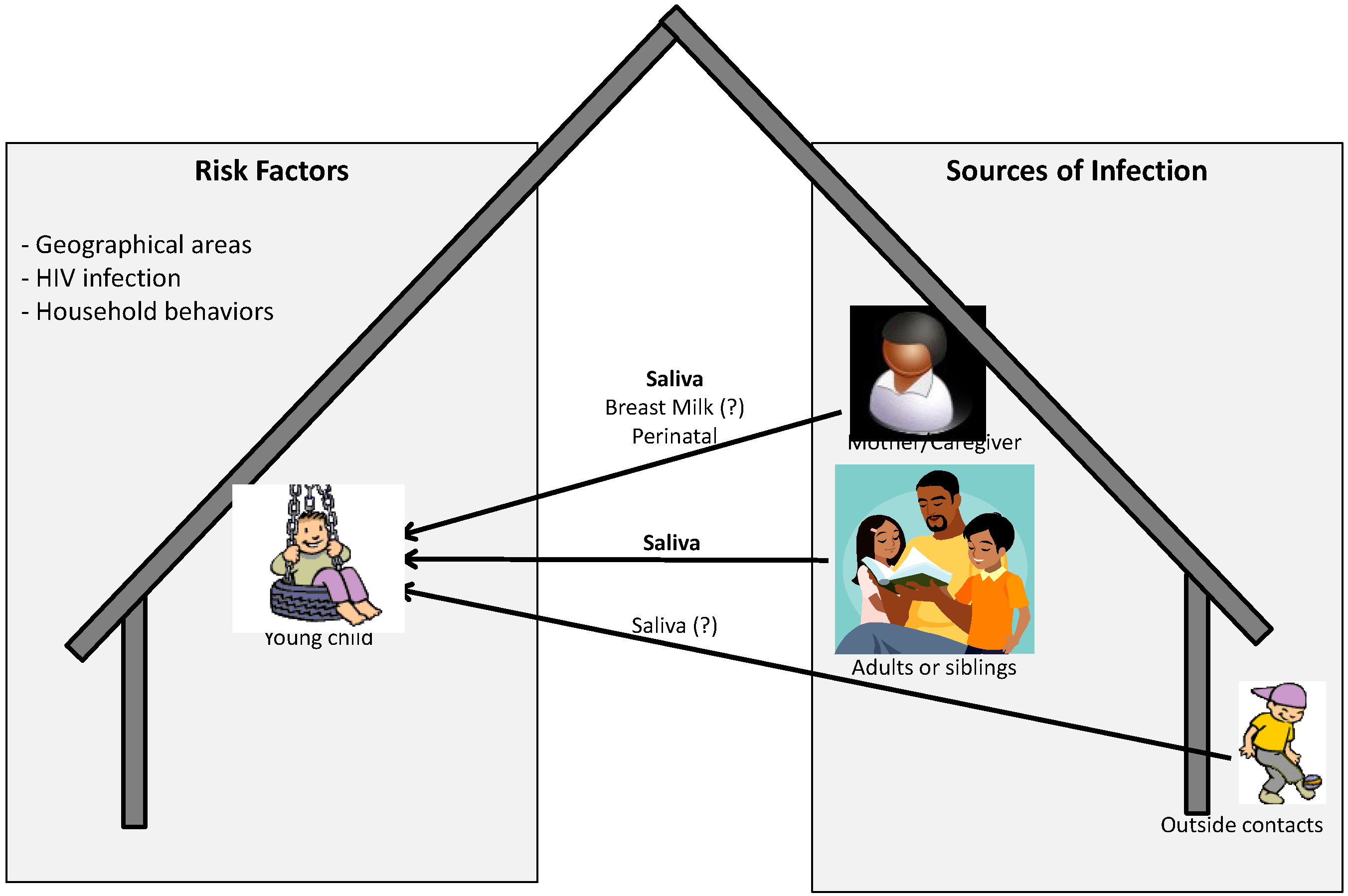
:1. Introduction
2. Modes of Transmission
3. Sources and Risk Factors Associated with Transmission

{kind=link}
| Reference (Year) | Country | Age (Years) | Percent Prevalence |
|---|---|---|---|
| Cao et al. (2014) [21] | Xinjiang, China | 0.5–5 | 48.3 |
| Wakeham et al. (2013) [64] | Entebbe, Uganda | 1 | 4 |
| 2 | 7 | ||
| 3 | 10 | ||
| 4 | 13 | ||
| 5 | 14 | ||
| Butler et al. (2011) [65] | Buziika B Parish, Uganda | 1.5–2 | 15.5 |
| 3–5 | 22.7 | ||
| 6–9 | 31.6 | ||
| 10–13 | 32.0 | ||
| Pfeiffer et al. (2010) [66] | Kampala, Uganda | 0–4 | 32.1 |
| 5–9 | 37.4 | ||
| 10–19 | 30.6 | ||
| Shirati District, Tanzania | 0–4 | 33.9 | |
| 5–9 | 33.7 | ||
| 10–19 | 32.4 | ||
| Lagos, Nigeria | 0–4 | 14.6 | |
| 5–9 | 2.3 | ||
| 10–19 | 83.1 | ||
| Butler et al. (2009) [38] | Kampala, Uganda | 2–8 | 10–30.6 |

4. Future Research
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Cesarman, E.; Chang, Y.; Moore, P.S.; Said, J.W.; Knowles, D.M. Kaposi’s sarcoma-associated herpesvirus-like DNA sequences in aids-related body-cavity-based lymphomas. N. Engl. J. Med. 1995, 332, 1186–1191. [Google Scholar] [CrossRef]
- Chang, Y.; Cesarman, E.; Pessin, M.S.; Lee, F.; Culpepper, J.; Knowles, D.M.; Moore, P.S. Identification of herpesvirus-like DNA sequences in aids-associated kaposi’s sarcoma. Science 1994, 266, 1865–1869. [Google Scholar] [CrossRef]
- Schulz, T.F. Kaposi’s sarcoma-associated herpesvirus (human herpesvirus-8). J. Gen. Virol. 1998, 79, 1573–1591. [Google Scholar]
- Soulier, J.; Grollet, L.; Oksenhendler, E.; Cacoub, P.; Cazals-Hatem, D.; Babinet, P.; d’Agay, M.F.; Clauvel, J.P.; Raphael, M.; Degos, L.; et al. Kaposi’s sarcoma-associated herpesvirus-like DNA sequences in multicentric castleman’s disease. Blood 1995, 86, 1276–1280. [Google Scholar]
- Sunil, M.; Reid, E.; Lechowicz, M.J. Update on hhv-8-associated malignancies. Curr. Infect. Dis. Rep. 2010, 12, 147–154. [Google Scholar] [CrossRef]
- Ziegler, J.L.; Bragg, K.; Abrams, D.; Beckstead, J.; Cogan, M.; Volberding, P.; Baer, D.; Wilkinson, L.; Rosenbaum, E.; Grant, K.; et al. High-grade non-hodgkin’s lymphoma in patients with aids. Ann. N. Y. Acad. Sci. 1984, 437, 412–419. [Google Scholar] [CrossRef]
- Cook-Mozaffari, P.; Newton, R.; Beral, V.; Burkitt, D.P. The geographical distribution of kaposi’s sarcoma and of lymphomas in africa before the aids epidemic. Br. J. Cancer 1998, 78, 1521–1528. [Google Scholar] [CrossRef]
- Blauvelt, A.; Sei, S.; Cook, P.M.; Schulz, T.F.; Jeang, K.T. Human herpesvirus 8 infection occurs following adolescence in the united states. J. Infect. Dis. 1997, 176, 771–774. [Google Scholar] [CrossRef]
- Dedicoat, M.; Newton, R. Review of the distribution of kaposi’s sarcoma-associated herpesvirus (kshv) in africa in relation to the incidence of kaposi’s sarcoma. Br. J. Cancer 2003, 88, 1–3. [Google Scholar] [CrossRef]
- Gao, S.J.; Kingsley, L.; Li, M.; Zheng, W.; Parravicini, C.; Ziegler, J.; Newton, R.; Rinaldo, C.R.; Saah, A.; Phair, J.; et al. Kshv antibodies among americans, italians and ugandans with and without kaposi’s sarcoma. Nat. Med. 1996, 2, 925–928. [Google Scholar] [CrossRef]
- Martro, E.; Bulterys, M.; Stewart, J.A.; Spira, T.J.; Cannon, M.J.; Thacher, T.D.; Bruns, R.; Pellett, P.E.; Dollard, S.C. Comparison of human herpesvirus 8 and epstein-barr virus seropositivity among children in areas endemic and non-endemic for kaposi’s sarcoma. J. Med. Virol. 2004, 72, 126–131. [Google Scholar] [CrossRef]
- Pellett, P.E.; Wright, D.J.; Engels, E.A.; Ablashi, D.V.; Dollard, S.C.; Forghani, B.; Glynn, S.A.; Goedert, J.J.; Jenkins, F.J.; Lee, T.H.; et al. Multicenter comparison of serologic assays and estimation of human herpesvirus 8 seroprevalence among us blood donors. Transfusion 2003, 43, 1260–1268. [Google Scholar] [CrossRef]
- Weiss, R.A.; Whitby, D.; Talbot, S.; Kellam, P.; Boshoff, C. Human herpesvirus type 8 and kaposi’s sarcoma. J. Natl. Cancer Inst. Monogr. 1998, 23, 51–54. [Google Scholar] [CrossRef]
- Stolka, K.; Ndom, P.; Hemingway-Foday, J.; Iriondo-Perez, J.; Miley, W.; Labo, N.; Stella, J.; Abassora, M.; Woelk, G.; Ryder, R.; et al. Risk factors for kaposi’s sarcoma among hiv-positive individuals in a case control study in cameroon. Cancer Epidemiol. 2014, 38, 137–143. [Google Scholar] [CrossRef]
- Biggar, R.J.; Whitby, D.; Marshall, V.; Linhares, A.C.; Black, F. Human herpesvirus 8 in brazilian amerindians: A hyperendemic population with a new subtype. J. Infect. Dis. 2000, 181, 1562–1568. [Google Scholar] [CrossRef]
- de Souza, V.A.; Sumita, L.M.; Nascimento, M.C.; Oliveira, J.; Mascheretti, M.; Quiroga, M.; Freire, W.S.; Tateno, A.; Boulos, M.; Mayaud, P.; et al. Human Herpesvirus-8 infection and oral shedding in Amerindian and non-Amerindian populations in the Brazilian amazon region. J. Infect. Dis. 2007, 196, 844–852. [Google Scholar] [CrossRef]
- De Sanjose, S.; Mbisa, G.; Perez-Alvarez, S.; Benavente, Y.; Sukvirach, S.; Hieu, N.T.; Shin, H.R.; Anh, P.T.; Thomas, J.; Lazcano, E.; et al. Geographic variation in the prevalence of kaposi sarcoma-associated herpesvirus and risk factors for transmission. J. Infect. Dis. 2009, 199, 1449–1456. [Google Scholar] [CrossRef]
- Fu, B.; Sun, F.; Li, B.; Yang, L.; Zeng, Y.; Sun, X.; Xu, F.; Rayner, S.; Guadalupe, M.; Gao, S.J.; et al. Seroprevalence of kaposi’s sarcoma-associated herpesvirus and risk factors in xinjiang, china. J. Med. Virol. 2009, 81, 1422–1431. [Google Scholar] [CrossRef]
- Mei, Q.; Ming, Z.W.; Ping, Y.X.; Hui, J.J.; Bin, Z.Y.; Hong, W.; Juan, L.; Zhe, C.Y.; Wei, T.; Han, Y. Hhv-8 seroprevalence in blood donors and hiv-positive individuals in shandong area, china. J. Infect. 2007, 55, 89–90. [Google Scholar] [CrossRef]
- Wang, H.; Liu, J.; Dilimulati; Li, L.; Ren, Z.; Wen, H.; Wang, X. Seroprevalence and risk factors of kaposi’s sarcoma-associated herpesvirus infection among the general uygur population from south and north region of xinjiang, china. Virol. J. 2011, 8, 539. [Google Scholar] [CrossRef]
- Cao, Y.; Minhas, V.; Tan, X.; Huang, J.; Wang, B.; Zhu, M.; Gao, Y.; Zhao, T.; Yang, L.; Wood, C. High prevalence of early childhood infection by kaposi’s sarcoma-associated herpesvirus in a minority population in china. Clin. Microbiol. Infect. 2014, 20, 475–481. [Google Scholar] [CrossRef]
- Bayley, A.C. Occurrence, clinical behaviour and management of kaposi’s sarcoma in zambia. Cancer Surv. 1991, 10, 53–71. [Google Scholar]
- Patil, P.; Elem, B.; Zumla, A. Pattern of adult malignancies in zambia (1980–1989) in light of the human immunodeficiency virus type 1 epidemic. J. Trop. Med. Hyg. 1995, 98, 281–284. [Google Scholar]
- Patil, P.S.; Elem, B.; Gwavava, N.J.; Urban, M.I. The pattern of paediatric malignancy in zambia (1980–1989): A hospital-based histopathological study. J. Trop. Med. Hyg. 1992, 95, 124–127. [Google Scholar]
- Chintu, C.; Athale, U.H.; Patil, P.S. Childhood cancers in zambia before and after the hiv epidemic. Arch. Dis. Child 1995, 73, 100–104. [Google Scholar] [CrossRef]
- Chen, T.; Hudnall, S.D. Anatomical mapping of human herpesvirus reservoirs of infection. Mod. Pathol. 2006, 19, 726–737. [Google Scholar] [CrossRef]
- Kedes, D.H.; Operskalski, E.; Busch, M.; Kohn, R.; Flood, J.; Ganem, D. The seroepidemiology of human herpesvirus 8 (kaposi’s sarcoma-associated herpesvirus): Distribution of infection in ks risk groups and evidence for sexual transmission. Nat. Med. 1996, 2, 918–924. [Google Scholar] [CrossRef]
- Mantina, H.; Kankasa, C.; Klaskala, W.; Brayfield, B.; Campbell, J.; Du, Q.; Bhat, G.; Kasolo, F.; Mitchell, C.; Wood, C. Vertical transmission of kaposi’s sarcoma-associated herpesvirus. Int. J. Cancer 2001, 94, 749–752. [Google Scholar] [CrossRef]
- Mbulaiteye, S.M.; Pfeiffer, R.M.; Whitby, D.; Brubaker, G.R.; Shao, J.; Biggar, R.J. Human herpesvirus 8 infection within families in rural tanzania. J. Infect. Dis. 2003, 187, 1780–1785. [Google Scholar] [CrossRef]
- Cannon, M.J.; Dollard, S.C.; Smith, D.K.; Klein, R.S.; Schuman, P.; Rich, J.D.; Vlahov, D.; Pellett, P.E. Blood-borne and sexual transmission of human herpesvirus 8 in women with or at risk for human immunodeficiency virus infection. N. Engl. J. Med. 2001, 344, 637–643. [Google Scholar] [CrossRef]
- Martin, J.N.; Ganem, D.E.; Osmond, D.H.; Page-Shafer, K.A.; Macrae, D.; Kedes, D.H. Sexual transmission and the natural history of human herpesvirus 8 infection. N. Engl. J. Med. 1998, 338, 948–954. [Google Scholar] [CrossRef]
- Melbye, M.; Cook, P.M.; Hjalgrim, H.; Begtrup, K.; Simpson, G.R.; Biggar, R.J.; Ebbesen, P.; Schulz, T.F. Risk factors for kaposi’s-sarcoma-associated herpesvirus (kshv/hhv-8) seropositivity in a cohort of homosexual men, 1981–1996. Int. J. Cancer 1998, 77, 543–548. [Google Scholar] [CrossRef]
- Dukers, N.H.; Renwick, N.; Prins, M.; Geskus, R.B.; Schulz, T.F.; Weverling, G.J.; Coutinho, R.A.; Goudsmit, J. Risk factors for human herpesvirus 8 seropositivity and seroconversion in a cohort of homosexual men. Am. J. Epidemiol. 2000, 151, 213–224. [Google Scholar] [CrossRef]
- Smith, N.A.; Sabin, C.A.; Gopal, R.; Bourboulia, D.; Labbet, W.; Boshoff, C.; Barlow, D.; Band, B.; Peters, B.S.; de Ruiter, A.; et al. Serologic evidence of human herpesvirus 8 transmission by homosexual but not heterosexual sex. J. Infect. Dis. 1999, 180, 600–606. [Google Scholar] [CrossRef]
- Eltom, M.A.; Mbulaiteye, S.M.; Dada, A.J.; Whitby, D.; Biggar, R.J. Transmission of human herpesvirus 8 by sexual activity among adults in lagos, nigeria. AIDS 2002, 16, 2473–2478. [Google Scholar] [CrossRef]
- Lavreys, L.; Chohan, B.; Ashley, R.; Richardson, B.A.; Corey, L.; Mandaliya, K.; Ndinya-Achola, J.O.; Kreiss, J.K. Human herpesvirus 8: Seroprevalence and correlates in prostitutes in mombasa, kenya. J. Infect. Dis. 2003, 187, 359–363. [Google Scholar] [CrossRef]
- Baeten, J.M.; Chohan, B.H.; Lavreys, L.; Rakwar, J.P.; Ashley, R.; Richardson, B.A.; Mandaliya, K.; Bwayo, J.J.; Kreiss, J.K. Correlates of human herpesvirus 8 seropositivity among heterosexual men in kenya. AIDS 2002, 16, 2073–2078. [Google Scholar] [CrossRef]
- Butler, L.M.; Dorsey, G.; Hladik, W.; Rosenthal, P.J.; Brander, C.; Neilands, T.B.; Mbisa, G.; Whitby, D.; Kiepiela, P.; Mosam, A.; et al. Kaposi sarcoma-associated herpesvirus (kshv) seroprevalence in population-based samples of african children: Evidence for at least 2 patterns of kshv transmission. J. Infect. Dis. 2009, 200, 430–438. [Google Scholar] [CrossRef]
- Malope, B.I.; MacPhail, P.; Mbisa, G.; MacPhail, C.; Stein, L.; Ratshikhopha, E.M.; Ndhlovu, L.; Sitas, F.; Whitby, D. No evidence of sexual transmission of kaposi’s sarcoma herpes virus in a heterosexual south african population. Aids 2008, 22, 519–526. [Google Scholar] [CrossRef]
- Zhang, T.; Yang, Y.; Yu, F.; Zhao, Y.; Lin, F.; Minhas, V.; Wood, C.; He, N. Kaposi`s sarcoma associated herpesvirus infection among female sex workers and general population women in shanghai, china: A cross-sectional study. BMC Infect. Dis. 2014, 14, 58. [Google Scholar] [CrossRef]
- Zavitsanou, A.; Sypsa, V.; Petrodaskalaki, M.; Psichogiou, M.; Katsoulidou, A.; Boletis, J.; Hadjiconstantinou, V.; Karalis, D.; Kalapothaki, V.; Hatzakis, A. Human herpesvirus 8 infection in hemodialysis patients. Am. J. Kidney Dis. 2006, 47, 167–170. [Google Scholar] [CrossRef]
- Whitby, D.; Howard, M.R.; Tenant-Flowers, M.; Brink, N.S.; Copas, A.; Boshoff, C.; Hatzioannou, T.; Suggett, F.E.; Aldam, D.M.; Denton, A.S.; et al. Detection of kaposi sarcoma associated herpesvirus in peripheral blood of hiv-infected individuals and progression to kaposi’s sarcoma. Lancet 1995, 346, 799–802. [Google Scholar] [CrossRef]
- Hladik, W.; Dollard, S.C.; Mermin, J.; Fowlkes, A.L.; Downing, R.; Amin, M.M.; Banage, F.; Nzaro, E.; Kataaha, P.; Dondero, T.J.; et al. Transmission of human herpesvirus 8 by blood transfusion. N. Engl. J. Med. 2006, 355, 1331–1338. [Google Scholar] [CrossRef]
- Hladik, W.; Pellett, P.E.; Hancock, J.; Downing, R.; Gao, H.; Packel, L.; Mimbe, D.; Nzaro, E.; Mermin, J. Association between transfusion with human herpesvirus 8 antibody-positive blood and subsequent mortality. J. Infect. Dis. 2012, 206, 1497–1503. [Google Scholar] [CrossRef]
- Lisco, A.; Barbierato, M.; Fiore, J.R.; Gasperini, P.; Favia, A.; Volpe, A.; Chironna, M.; Pastore, G.; Chieco-Bianchi, L.; Calabro, M.L. Pregnancy and human herpesvirus 8 reactivation in human immunodeficiency virus type 1-infected women. J. Clin. Microbiol. 2006, 44, 3863–3871. [Google Scholar] [CrossRef]
- Gutierrez-Ortega, P.; Hierro-Orozco, S.; Sanchez-Cisneros, R.; Montano, L.F. Kaposi’s sarcoma in a 6-day-old infant with human immunodeficiency virus. Arch. Dermatol. 1989, 125, 432–433. [Google Scholar] [CrossRef]
- Ahlfors, K.; Ivarsson, S.A. Cytomegalovirus in breast milk of swedish milk donors. Scand. J. Infect. Dis. 1985, 17, 11–13. [Google Scholar] [CrossRef]
- Dworsky, M.; Yow, M.; Stagno, S.; Pass, R.F.; Alford, C. Cytomegalovirus infection of breast milk and transmission in infancy. Pediatrics 1983, 72, 295–299. [Google Scholar]
- Fujisaki, H.; Tanaka-Taya, K.; Tanabe, H.; Hara, T.; Miyoshi, H.; Okada, S.; Yamanishi, K. Detection of human herpesvirus 7 (hhv-7) DNA in breast milk by polymerase chain reaction and prevalence of hhv-7 antibody in breast-fed and bottle-fed children. J. Med. Virol. 1998, 56, 275–279. [Google Scholar] [CrossRef]
- Hamprecht, K.; Vochem, M.; Baumeister, A.; Boniek, M.; Speer, C.P.; Jahn, G. Detection of cytomegaloviral DNA in human milk cells and cell free milk whey by nested pcr. J. Virol. Methods 1998, 70, 167–176. [Google Scholar] [CrossRef]
- Junker, A.K.; Thomas, E.E.; Radcliffe, A.; Forsyth, R.B.; Davidson, A.G.; Rymo, L. Epstein-barr virus shedding in breast milk. Am. J. Med. Sci. 1991, 302, 220–223. [Google Scholar] [CrossRef]
- Dedicoat, M.; Newton, R.; Alkharsah, K.R.; Sheldon, J.; Szabados, I.; Ndlovu, B.; Page, T.; Casabonne, D.; Gilks, C.F.; Cassol, S.A.; et al. Mother-to-child transmission of human herpesvirus-8 in south africa. J. Infect. Dis. 2004, 190, 1068–1075. [Google Scholar] [CrossRef]
- Brayfield, B.P.; Kankasa, C.; West, J.T.; Muyanga, J.; Bhat, G.; Klaskala, W.; Mitchell, C.D.; Wood, C. Distribution of kaposi sarcoma-associated herpesvirus/human herpesvirus 8 in maternal saliva and breast milk in zambia: Implications for transmission. J. Infect. Dis. 2004, 189, 2260–2270. [Google Scholar] [CrossRef]
- Boldogh, I.; Szaniszlo, P.; Bresnahan, W.A.; Flaitz, C.M.; Nichols, M.C.; Albrecht, T. Kaposi’s sarcoma herpesvirus-like DNA sequences in the saliva of individuals infected with human immunodeficiency virus. Clin. Infect. Dis. 1996, 23, 406–407. [Google Scholar] [CrossRef]
- Koelle, D.M.; Huang, M.L.; Chandran, B.; Vieira, J.; Piepkorn, M.; Corey, L. Frequent detection of kaposi’s sarcoma-associated herpesvirus (human herpesvirus 8) DNA in saliva of human immunodeficiency virus-infected men: Clinical and immunologic correlates. J. Infect. Dis. 1997, 176, 94–102. [Google Scholar] [CrossRef]
- Webster-Cyriaque, J.; Edwards, R.H.; Quinlivan, E.B.; Patton, L.; Wohl, D.; Raab-Traub, N. Epstein-barr virus and human herpesvirus 8 prevalence in human immunodeficiency virus-associated oral mucosal lesions. J. Infect. Dis. 1997, 175, 1324–1332. [Google Scholar] [CrossRef]
- Casper, C.; Krantz, E.; Selke, S.; Kuntz, S.R.; Wang, J.; Huang, M.L.; Pauk, J.S.; Corey, L.; Wald, A. Frequent and asymptomatic oropharyngeal shedding of human herpesvirus 8 among immunocompetent men. J. Infect. Dis. 2007, 195, 30–36. [Google Scholar] [CrossRef]
- Phipps, W.; Saracino, M.; Selke, S.; Huang, M.L.; Jaoko, W.; Mandaliya, K.; Wald, A.; Casper, C.; McClelland, R.S. Oral hhv-8 replication among women in mombasa, kenya. J. Med. Virol. 2014, 86, 1759–1765. [Google Scholar] [CrossRef]
- DeSantis, S.M.; Pau, C.P.; Archibald, L.K.; Nwanyanwu, O.C.; Kazembe, P.N.; Dobbie, H.; Jarvis, W.R.; Jason, J. Demographic and immune correlates of human herpesvirus 8 seropositivity in malawi, africa. Int. J. Infect. Dis. 2002, 6, 266–271. [Google Scholar] [CrossRef]
- Plancoulaine, S.; Abel, L.; Tregouet, D.; Duprez, R.; van Beveren, M.; Tortevoye, P.; Froment, A.; Gessain, A. Respective roles of serological status and blood specific antihuman herpesvirus 8 antibody levels in human herpesvirus 8 intrafamilial transmission in a highly endemic area. Cancer Res. 2004, 64, 8782–8787. [Google Scholar] [CrossRef]
- Minhas, V.; Crabtree, K.L.; Chao, A.; M’Soka, T.J.; Kankasa, C.; Bulterys, M.; Mitchell, C.D.; Wood, C. Early childhood infection by human herpesvirus 8 in zambia and the role of human immunodeficiency virus type 1 coinfection in a highly endemic area. Am. J. Epidemiol. 2008, 168, 311–320. [Google Scholar] [CrossRef]
- Alkharsah, K.R.; Dedicoat, M.; Blasczyk, R.; Newton, R.; Schulz, T.F. Influence of hla alleles on shedding of kaposi sarcoma-associated herpesvirus in saliva in an african population. J. Infect. Dis. 2007, 195, 809–816. [Google Scholar] [CrossRef]
- Olp, L.N.; Shea, D.M.; White, M.K.; Gondwe, C.; Kankasa, C.; Wood, C. Early childhood infection of kaposi’s sarcoma-associated herpesvirus in zambian households: A molecular analysis. Int. J. Cancer 2012, 132, 1182–1190. [Google Scholar]
- Wakeham, K.; Webb, E.L.; Sebina, I.; Nalwoga, A.; Muhangi, L.; Miley, W.; Johnston, W.T.; Ndibazza, J.; Whitby, D.; Newton, R.; et al. Risk Factors for Seropositivity to Kaposi Sarcoma-Associated Herpesvirus among Children in Uganda. J. Acquir. Immune Defic. Syndr. 2013, 63, 228–233. [Google Scholar] [CrossRef]
- Butler, L.M.; Were, W.A.; Balinandi, S.; Downing, R.; Dollard, S.; Neilands, T.B.; Gupta, S.; Rutherford, G.W.; Mermin, J. Human Herpesvirus 8 Infection in Children and Adults in a Population-Based Study in Rural Uganda. J. Infect. Dis. 2011, 203, 625–634. [Google Scholar] [CrossRef]
- Pfeiffer, R.M.; Wheeler, W.A.; Mbisa, G.; Whitby, D.; Goedert, J.J.; de The, G.; Mbulaiteye, S.M. Geographic Heterogeneity of Prevalence of the Human Herpesvirus 8 in Sub-Saharan Africa: Clues about Etiology. Ann. Epidemiol. 2010, 20, 958–963. [Google Scholar] [CrossRef]
- Coluzzi, M.; Manno, D.; Guzzinati, S.; Tognazzo, S.; Zambon, P.; Arca, B.; Costantini, C.; Ascoli, V. The bloodsucking arthropod bite as possible cofactor in the transmission of human herpesvirus-8 infection and in the expression of kaposi’s sarcoma disease. Parassitologia 2002, 44, 123–129. [Google Scholar]
- Lin, C.J.; Katongole-Mbidde, E.; Byekwaso, T.; Orem, J.; Rabkin, C.S.; Mbulaiteye, S.M. Intestinal parasites in kaposi sarcoma patients in uganda: Indication of shared risk factors or etiologic association. Am. J. Trop. Med. Hyg. 2008, 78, 409–412. [Google Scholar]
- Whitby, D.; Marshall, V.A.; Bagni, R.K.; Miley, W.J.; McCloud, T.G.; Hines-Boykin, R.; Goedert, J.J.; Conde, B.A.; Nagashima, K.; Mikovits, J.; et al. Reactivation of kaposi’s sarcoma-associated herpesvirus by natural products from kaposi’s sarcoma endemic regions. Int. J. Cancer 2007, 120, 321–328. [Google Scholar] [CrossRef]
- Ziegler, J.L. Endemic kaposi’s sarcoma in africa and local volcanic soils. Lancet 1993, 342, 1348–1351. [Google Scholar] [CrossRef]
- Renne, R.; Dittmer, D.; Kedes, D.; Schmidt, K.; Desrosiers, R.C.; Luciw, P.A.; Ganem, D. Experimental transmission of kaposi’s sarcoma-associated herpesvirus (kshv/hhv-8) to siv-positive and siv-negative rhesus macaques. J. Med. Primatol. 2004, 33, 1–9. [Google Scholar] [CrossRef]
- Chang, H.; Wachtman, L.M.; Pearson, C.B.; Lee, J.S.; Lee, H.R.; Lee, S.H.; Vieira, J.; Mansfield, K.G.; Jung, J.U. Non-human primate model of kaposi’s sarcoma-associated herpesvirus infection. PLoS Pathog. 2009, 5, e1000606. [Google Scholar] [CrossRef]
- Brehm, M.A.; Shultz, L.D.; Greiner, D.L. Humanized mouse models to study human diseases. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 120–125. [Google Scholar] [CrossRef]
- Wege, A.K.; Melkus, M.W.; Denton, P.W.; Estes, J.D.; Garcia, J.V. Functional and phenotypic characterization of the humanized blt mouse model. Curr. Top. Microbiol. Immunol. 2008, 324, 149–165. [Google Scholar]
- Wang, L.X.; Kang, G.; Kumar, P.; Lu, W.; Li, Y.; Zhou, Y.; Li, Q.; Wood, C. Humanized-blt mouse model of kaposi’s sarcoma-associated herpesvirus infection. Proc. Natl. Acad. Sci. USA 2014, 111, 3146–3151. [Google Scholar] [CrossRef]
- Labo, N.; Miley, W.; Marshall, V.; Gillette, W.; Esposito, D.; Bess, M.; Turano, A.; Uldrick, T.; Polizzotto, M.N.; Wyvill, K.M.; et al. Heterogeneity and breadth of host antibody response to kshv infection demonstrated by systematic analysis of the kshv proteome. PLoS Pathog. 2014, 10, e1004046. [Google Scholar] [CrossRef]
- Lennette, E.T.; Blackbourn, D.J.; Levy, J.A. Antibodies to human herpesvirus type 8 in the general population and in kaposi’s sarcoma patients. Lancet 1996, 348, 858–861. [Google Scholar] [CrossRef]
- Kimball, L.E.; Casper, C.; Koelle, D.M.; Morrow, R.; Corey, L.; Vieira, J. Reduced levels of neutralizing antibodies to kaposi sarcoma-associated herpesvirus in persons with a history of kaposi sarcoma. J. Infect. Dis. 2004, 189, 2016–2022. [Google Scholar] [CrossRef]
- Inoue, N.; Spira, T.; Lam, L.; Corchero, J.L.; Luo, W. Comparison of serologic responses between kaposi’s sarcoma-positive and -negative men who were seropositive for both human herpesvirus 8 and human immunodeficiency virus. J. Med. Virol. 2004, 74, 202–206. [Google Scholar] [CrossRef]
- Kumar, P.; Kuwa, N.Y.; Minhas, V.; Marimo, C.; Shea, D.M.; Kankasa, C.; Wood, C. Higher levels of neutralizing antibodies against KSHV in KS patients compared to asymptomatic individuals from Zambia. PLoS One 2013, 8, e71254. [Google Scholar] [CrossRef]
- Lepone, L.; Rappocciolo, G.; Knowlton, E.; Jais, M.; Piazza, P.; Jenkins, F.J.; Rinaldo, C.R. Monofunctional and polyfunctional CD8+ T cell responses to human Herpesvirus 8 lytic and latency proteins. Clin. Vaccine Immunol. 2010, 17, 1507–1516. [Google Scholar] [CrossRef]
- Robey, R.C.; Mletzko, S.; Gotch, F.M. The T-Cell immune response against kaposi's sarcoma-associated herpesvirus. Adv. Virol. 2010, 2010, 340356. [Google Scholar] [CrossRef]
- Carrieri, M.P.; Pradier, C.; Piselli, P.; Piche, M.; Rosenthal, E.; Heudier, P.; Durant, J.; Serraino, D. Reduced incidence of kaposi’s sarcoma and of systemic non-hodgkin’s lymphoma in hiv-infected individuals treated with highly active antiretroviral therapy. Int. J. Cancer 2003, 103, 142–144. [Google Scholar] [CrossRef]
- Franceschi, S.; Maso, L.D.; Rickenbach, M.; Polesel, J.; Hirschel, B.; Cavassini, M.; Bordoni, A.; Elzi, L.; Ess, S.; Jundt, G.; et al. Kaposi sarcoma incidence in the swiss hiv cohort study before and after highly active antiretroviral therapy. Br. J. Cancer 2008, 99, 800–804. [Google Scholar] [CrossRef]
- Ngalamika, O.; Minhas, V.; Wood, C. Kaposi's sarcoma at the university teaching hospital, Lusaka, Zambia in the antiretroviral therapy era. Int. J. Cancer 2014. [Google Scholar] [CrossRef]
- Bohlius, J.; Valeri, F.; Maskew, M.; Prozesky, H.; Garone, D.; Sengayi, M.; Fox, M.P.; Davies, M.A.; Egger, M. Kaposi’s sarcoma in hiv-infected patients in south africa: Multicohort study in the antiretroviral therapy era. Int. J. Cancer 2014, 135, 2644–2652. [Google Scholar] [CrossRef]
- Stebbing, J.; Portsmouth, S.; Bower, M. Insights into the molecular biology and sero-epidemiology of kaposi's sarcoma. Curr. Opin. Infect. Dis. 2003, 16, 25–31. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minhas, V.; Wood, C. Epidemiology and Transmission of Kaposi’s Sarcoma-Associated Herpesvirus. Viruses 2014, 6, 4178-4194. https://doi.org/10.3390/v6114178
Minhas V, Wood C. Epidemiology and Transmission of Kaposi’s Sarcoma-Associated Herpesvirus. Viruses. 2014; 6(11):4178-4194. https://doi.org/10.3390/v6114178
Chicago/Turabian StyleMinhas, Veenu, and Charles Wood. 2014. "Epidemiology and Transmission of Kaposi’s Sarcoma-Associated Herpesvirus" Viruses 6, no. 11: 4178-4194. https://doi.org/10.3390/v6114178
APA StyleMinhas, V., & Wood, C. (2014). Epidemiology and Transmission of Kaposi’s Sarcoma-Associated Herpesvirus. Viruses, 6(11), 4178-4194. https://doi.org/10.3390/v6114178