Apple Latent Spherical Virus Vector as Vaccine for the Prevention and Treatment of Mosaic Diseases in Pea, Broad Bean, and Eustoma Plants by Bean Yellow Mosaic Virus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
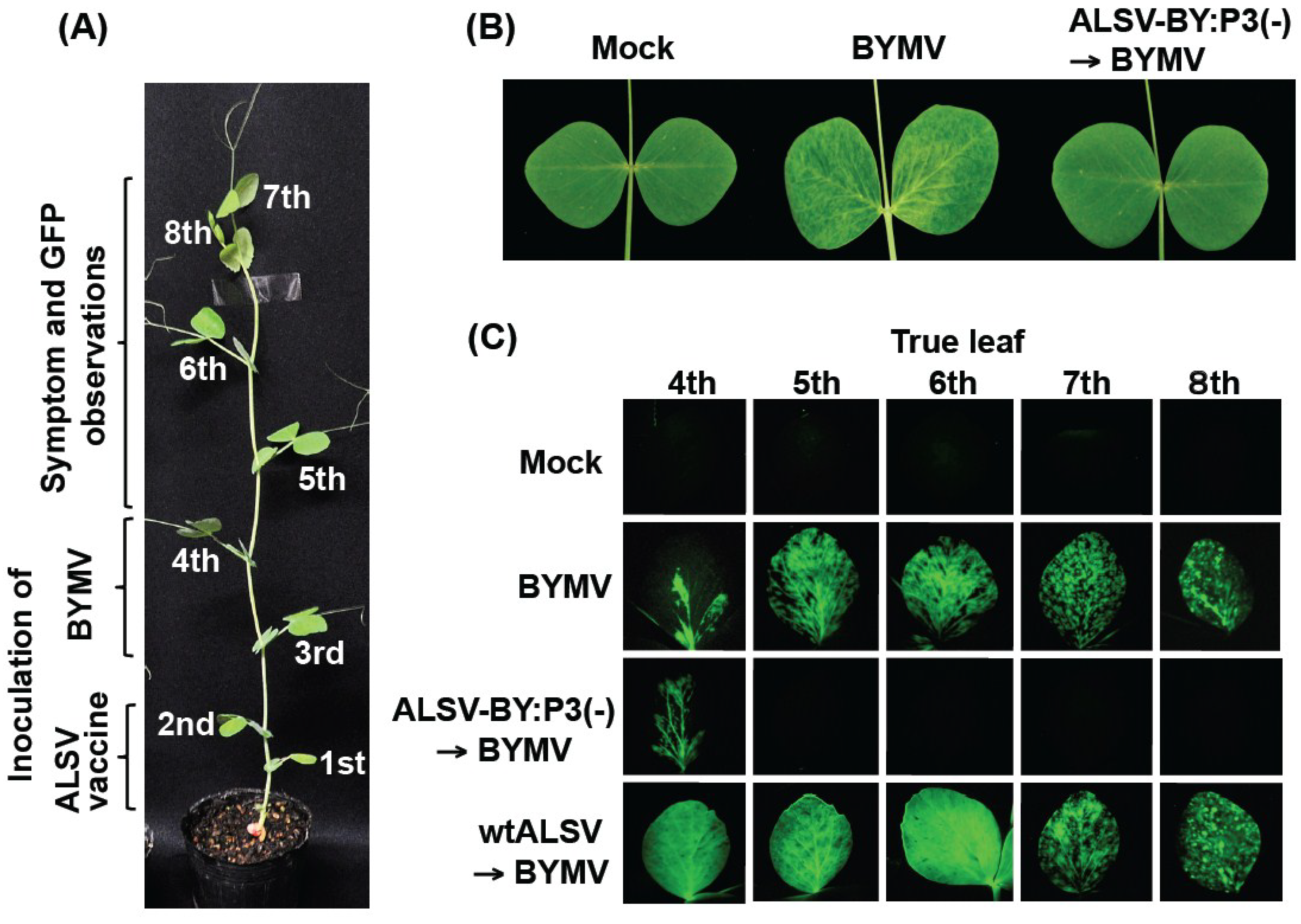
2.1. Effects of the ALSV Vaccine in Preventing Pea Mosaic Disease


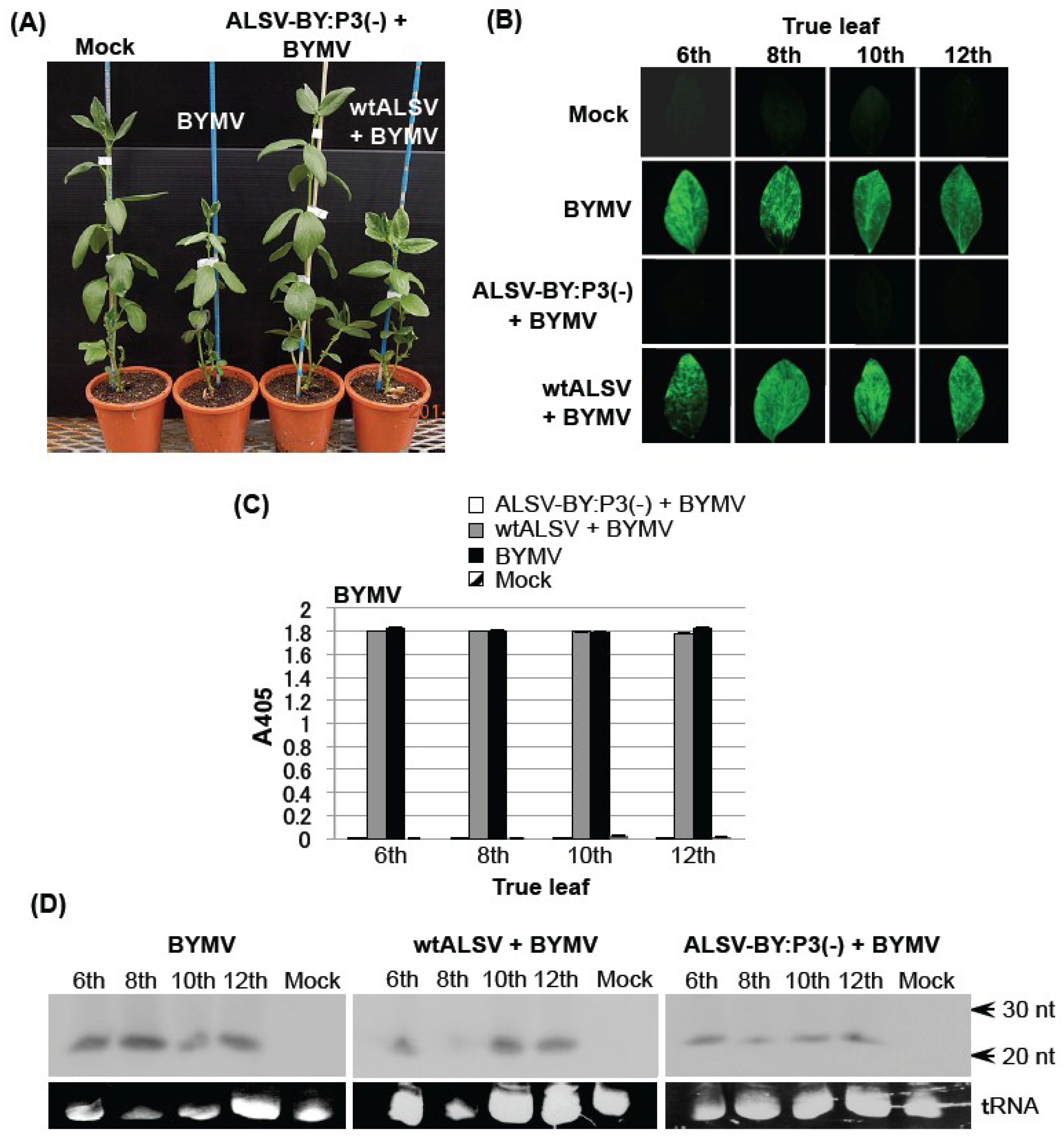
2.2. Effects of the ALSV Vaccine in Preventing Broad Bean Mosaic Disease

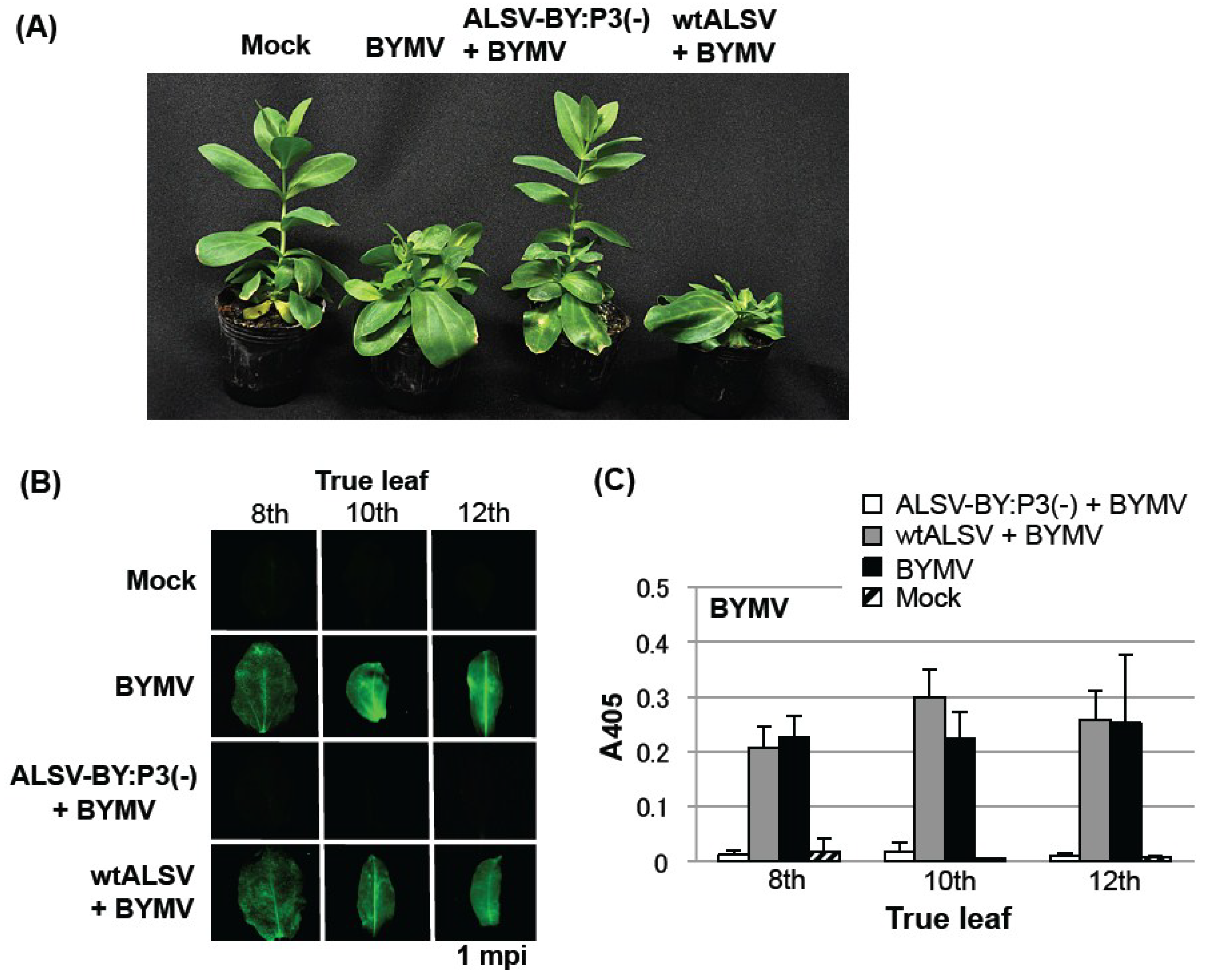
2.3. Effect of the ALSV Vaccine in Preventing Eustoma Mosaic Disease


2.4. Curative Effects of the ALSV Vaccine against Mosaic Diseases of Pea and Eustoma Plants

3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. ALSV Vaccine and BYMV
4.3. Preparation of ALSV Inocula
4.4. Viral Inoculation
4.5. Virus Detection
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Koganezawa, H.; Ito, T. Apple latent spherical virus. In Virus and Virus-Like Diseases of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T., Jalkmann, W., Eds.; APS Press: Minnesota, MN, USA, 2011; pp. 23–24. [Google Scholar]
- Li, C.; Yoshikawa, N.; Takahashi, T.; Ito, T.; Yoshida, K.; Koganezawa, H. Nucleotide sequence and genome organization of apple latent spherical virus: a new virus classified into the family Comoviridae. J. Gen. Virol. 2000, 81, 541–547. [Google Scholar]
- Igarashi, A.; Yamagata, K.; Sugai, T.; Takahashi, Y.; Sugawara, E.; Tamura, A.; Yaegashi, H.; Yamagishi, N.; Takahashi, T.; Isogai, M.; et al. Apple latent spherical virus vectors for reliable and effective virus-induced gene silencing among a broad range of plants including tobacco, tomato, Arabidopsis thaliana, cucurbits, and legumes. Virology 2009, 386, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Sasaki, N.; Isogai, M.; Yoshikawa, N. Stable expression of foreign proteins in herbaceous and apple plants using Apple latent spherical virus RNA2 vectors. Arch. Virol. 2004, 149, 1541–1558. [Google Scholar] [PubMed]
- Sasaki, S.; Yamagishi, N.; Yoshikawa, N. Efficient virus-induced gene silencing in apple, pear and Japanese pear using Apple latent spherical virus vectors. Plant Methods 2011, 7, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, N.; Yoshikawa, N. Virus-induced gene silencing in soybean seeds and the emergence stage of soybean plants with Apple latent spherical virus vectors. Plant Mol. Biol. 2009, 71, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Yaegashi, H.; Yamatsuta, T.; Takahashi, T.; Li, C.; Isogai, M.; Kobori, T.; Ohki, S.; Yoshikawa, N. Characterization of virus-induced gene silencing in tobacco plants infected with apple latent spherical virus. Arch. Virol. 2007, 152, 1839–1849. [Google Scholar] [CrossRef]
- Tamura, A.; Kato, T.; Taki, A.; Sone, M.; Satoh, N.; Yamagishi, N.; Takahashi, T.; Ryo, B.-S.; Natsuaki, T.; Yoshikawa, N. Preventive and curative effects of Apple latent spherical virus vectors harboring part of the target virus genome against potyvirus and cucumovirus infections. Virology 2013, 446, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Hannon, G.J. RNA interference. Nature 2002, 418, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Plasterk, R.H. RNA silencing: the genome’s immune system. Science 2002, 296, 1263–1265. [Google Scholar] [CrossRef] [PubMed]
- Vance, V.; Vaucheret, H. RNA silencing in plants-defense and counterdefense. Science 2001, 292, 2277–2280. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. RNA silencing as a plant immune system against viruses. Trends Genet. 2001, 17, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Induction and suppression of RNA silencing: Insights from viral infections. Nat. Rev. Genet. 2005, 6, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, P.M.; Wang, M.-B.; Lough, T. Gene silencing as an adaptive defence against viruses. Nature 2001, 411, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Lecellier, C.-H.; Voinnnet, O. RNA silencing: no mercy for viruses. Immunol. Rev. 2004, 198, 285–303. [Google Scholar] [CrossRef] [PubMed]
- Mlotshwa, S.; Voinnet, O.; Mette, M.F.; Matzke, M.; Vaucheret, H.; Ding, S.W.; Pruss, G.; Vance, V.B. RNA silencing and the mobile silencing signal. Plant Cell 2002, 14, S289–S301. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O.; Baulcombe, D.C. Systemic signaling in gene silencing. Nature 1997, 389, 553. [Google Scholar] [CrossRef] [PubMed]
- Culver, J.N. Tobamovirus cross protection using a potexvirus vector. Virology 1996, 226, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Ratcliff, F.G.; MacFarlane, S.A.; Baulcombe, D.C. Gene silencing without DNA: RNA-mediated cross-protection between viruses. Plant Cell 1999, 11, 1207–1216. [Google Scholar] [CrossRef] [PubMed]
- Taki, A.; Yamagishi, N.; Yoshikawa, N. Development of apple latent spherical virus-based vaccines against three tospoviruses. Virus Res. 2013, 176, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.J.; Zerbini, F.M.; French, R.; Rabenstein, F.; Strenger, D.C.; Valkonen, J.P.T. Potyviridae. In Virus Taxonomy Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.G., Adams, M.J., Carstens, E.B., Lefkowits, E.J., Eds.; Academic Press: Tokyo, Japan, 2012; pp. 1069–1089. [Google Scholar]
- Description of Plant Viruses, No. 40. Available online: http://www.dpvweb.net/dpv/showdpv.php?dpvno=040 (accessed on 25 September 2014).
- Wylie, S.J.; Coutts, B.A.; Jones, M.G.K.; Jones, R.A.C. Phylogenetic analysis of bean yellow mosaic virus isolates from four continents: Relationship between the seven groups found and their hosts and origins. Plant Dis. 2008, 92, 1596–1603. [Google Scholar] [CrossRef]
- Sasaya, T.; Nozu, Y.; Koganezawa, H. Biological and serological comparisons of bean yellow mosaic virus (BYMV) isolates in Japan. Ann. Phytopath. Soc. Jpn. 1998, 64, 24–33. [Google Scholar] [CrossRef]
- Shukla, D.D.; Ward, C.W. Amino acid sequence homology of coat proteins as a basis for identification and classification of the potyvirus group. J. Gen. Virol. 1988, 69, 2703–2710. [Google Scholar] [CrossRef]
- Nicaise, V.; Gallois, J.L.; Chafiai, F.; Allen, L.M.; Schrdi-Levraud, V.; Browning, K.S.; Candresse, T.; Caranta, C.; Le Gall, O.; German-Retana, S. Coordinated and selective recruitment of elF4E and elF4G factors for potyvirus infection in Arabidopsis thaliana. FEBS Lett. 2007, 581, 1041–1046. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Kohalmi, S.E.; Svircev, A.; Wang, A.; Sanfacon, H.; Tian, L. Silencing of the host factor elF(iso)4E gene confers plum pox virus resistance in plum. PLoS One 2013, 8, e50627. [Google Scholar] [CrossRef] [PubMed]
- Plant Diseases in Japan; Kishi, K. (Ed.) Zenkoku Nouson Kyouiku Kyoukai Press: Tokyo, Japan, 1998.
- Orellana, R.G.; Fan, F.F. Nodule infection by bean yellow mosaic virus in Phaseolus vulgaris. Appl. Environ. Microbiol. 1978, 36, 814–818. [Google Scholar] [PubMed]
- Rohani, B.; Habibi, M.K.; Mosahebi, G. Nodule infection by bean yellow mosaic virus in Vicia faba and molecular characterization of it. Commun. Agric. Appl. Biol. Sci. 2008, 73, 303–306. [Google Scholar] [PubMed]
- Walkey, D.G.A.; Lecoq, H.; Collier, R.; Dobson, S. Studies on the control of zucchini yellow mosaic virus in courgettes by mild strain protection. Plant Pathol. 1992, 41, 762–771. [Google Scholar] [CrossRef]
- Wen, F.; Lister, R.M.; Fattouh, F.A. Cross-protection among strains of barley yellow dwarf virus. J. Gen. Virol. 1991, 72, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, N.; Iwate University, Morioka, Japan. Unpublished work. 2014.
- Maejima, K.; Himeno, M.; Komatsu, K.; Takinami, Y.; Hashimoto, M.; Takahashi, S.; Yamaji, Y.; Oshima, K.; Namba, S. Molecular epidemiology of Plum pox virus in Japan. Phytopathology 2011, 101, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Maejima, K.; Hoshi, H.; Hashimoto, M.; Himeno, M.; Kawanishi, T.; Komatsu, K.; Yamaji, Y.; Hamamoto, H.; Namba, S. First report of plum pox virus infecting Japanese apricot (Prunus mume Sieb. et Zucc.) in Japan. J. Gen. Plant Pathol. 2010, 76, 229–231. [Google Scholar] [CrossRef]
- Takahashi, T.; Sugawara, T.; Yamatsuta, T.; Isogai, M.; Natsuaki, T.; Yoshikawa, N. Analysis of the spatial distribution of identical and two distinct virus populations differently labeled with cyan and yellow fluorescent proteins in coinfected plants. Phytopathology 2007, 97, 1200–1206. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, N.; Sasaki, S.; Yoshikawa, N. Highly efficient method for inoculation of apple viruses to apple seedlings. Julius-Kuhn-Archiv. 2010, 427, 226–229. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Satoh, N.; Kon, T.; Yamagishi, N.; Takahashi, T.; Natsuaki, T.; Yoshikawa, N. Apple Latent Spherical Virus Vector as Vaccine for the Prevention and Treatment of Mosaic Diseases in Pea, Broad Bean, and Eustoma Plants by Bean Yellow Mosaic Virus. Viruses 2014, 6, 4242-4257. https://doi.org/10.3390/v6114242
Satoh N, Kon T, Yamagishi N, Takahashi T, Natsuaki T, Yoshikawa N. Apple Latent Spherical Virus Vector as Vaccine for the Prevention and Treatment of Mosaic Diseases in Pea, Broad Bean, and Eustoma Plants by Bean Yellow Mosaic Virus. Viruses. 2014; 6(11):4242-4257. https://doi.org/10.3390/v6114242
Chicago/Turabian StyleSatoh, Nozomi, Tatsuya Kon, Noriko Yamagishi, Tsubasa Takahashi, Tomohide Natsuaki, and Nobuyuki Yoshikawa. 2014. "Apple Latent Spherical Virus Vector as Vaccine for the Prevention and Treatment of Mosaic Diseases in Pea, Broad Bean, and Eustoma Plants by Bean Yellow Mosaic Virus" Viruses 6, no. 11: 4242-4257. https://doi.org/10.3390/v6114242
APA StyleSatoh, N., Kon, T., Yamagishi, N., Takahashi, T., Natsuaki, T., & Yoshikawa, N. (2014). Apple Latent Spherical Virus Vector as Vaccine for the Prevention and Treatment of Mosaic Diseases in Pea, Broad Bean, and Eustoma Plants by Bean Yellow Mosaic Virus. Viruses, 6(11), 4242-4257. https://doi.org/10.3390/v6114242