Role of Human Cytomegalovirus Tegument Proteins in Virion Assembly

Abstract
:1. Introduction
2. Overview of Replication and Assembly

{kind=link}
| Gene Designation | Relative Abundance a | Phenotype b |
|---|---|---|
| Inner Tegument | ||
| UL32 (pp150) | 9.1% | Essential [39,55] |
| UL48 | 12.6% | Essential [39]; Severe growth defect (>104 fold drop in titer) [55] |
| Nuclear Tegument | ||
| UL26 | 0.1% | Non-essential, Small plaque phenotype with severe growth defect [39,55] |
| UL35 | 0.5% | Moderate growth defect (1,000-fold decrease in titer), Essential at low multiplicity [56] |
| UL82 (pp71) | 8.9% | Non-essential, Severe growth defect at low multiplicity [57,58] |
| UL83 (pp65) | 15.4% | Non-essential [59], Important for replication in macrophages [60], Slight growth defect (10-fold) at low multiplicity in fibroblasts [61] |
| UL94 | 1.2% | Essential [39,62] |
| UL97 | 0.1% | Non-essential, Severe growth defect [63] |
| TRS1 | 0.6% | Non-essential, Moderate growth defect (~200-fold decrease in titer) [64] |
| Cytoplasmic Tegument | ||
| UL71 | 0.1% | Essential [39] |
| UL99 (pp28) | Unknown | Essential [39,55,65] |
3. The Inner Tegument Proteins
3.1. pp150 (ppUL32)
3.2. pUL48
4. The Nuclear Tegument Proteins
4.1. pUL26
4.2. pUL35
| Protein Name | Protein Function | # of Peptides Matched | Ion Score b (−10Log(P)) | Sequence Coverage |
|---|---|---|---|---|
| pTRS1 | Transcriptional Regulation/Immune Evasion [12] | 78 | 2439 | 44% |
| pIRS1 | Transcriptional Regulation/Immune Evasion [12] | 60 | 1809 | 41% |
| pUL44 | DNA Processivity Factor [94] | 66 | 2413 | 82% |
| pUL86 | Major Capsid Protein [95] | 5 | 339 | 8% |
| ppUL97 | Viral Kinase [96] | 5 | 272 | 8% |
| pp65 | Tegument protein [12] | 236 | 2580 | 71% |
| pUL50 | Nuclear Egress [97] | 9 | 276 | 26% |
| pUL52 | Genome Cleavage and Packaging [98] | 7 | 256 | 14% |
| pUL56 | Terminase Subunit [99,100] | 11 | 385 | 26% |
| pUL88 | Tegument Protein [100] | 7 | 135 | 10% |
| pUL35 | Tegument Protein [12] | 6 | 289 | 29% |
4.3. pp71 (ppUL82)
4.4. pp65 (ppUL83)

4.5. pUL94
4.6. ppUL97
4.7. pTRS1
5. The Cytoplasmic Tegument Proteins
5.1. pUL71
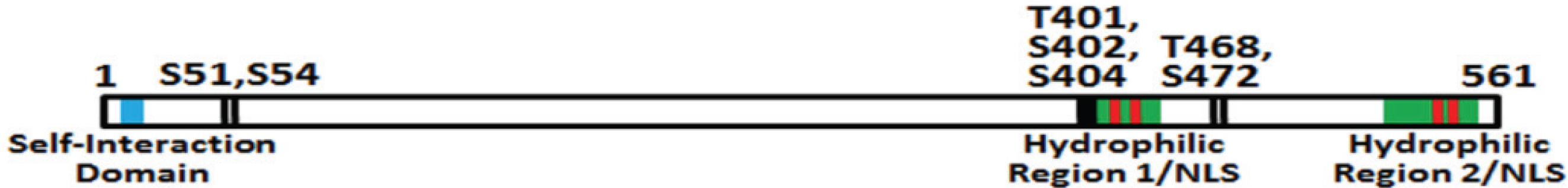
5.2. pp28
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Cannon, M.J.; Schmid, D.S.; Hyde, T.B. Review of cytomegalovirus seroprevalence and demographic characteristics associated with infection. Rev. Med. Virol. 2010, 20, 202–213. [Google Scholar] [CrossRef]
- Drew, W.L. Cytomegalovirus infection in patients with AIDS. Clin. Infect. Dis. 1992, 14, 608–615. [Google Scholar] [CrossRef]
- Griffiths, P.D.; Clark, D.A.; Emery, V.C. Betaherpesviruses in transplant recipients. J. Antimicrob. Chemother. 2000, 45, 29–34. [Google Scholar] [CrossRef]
- Steininger, C.; Puchhammer-Stockl, E.; Popow-Kraupp, T. Cytomegalovirus disease in the era of highly active antiretroviral therapy (HAART). J. Clin. Virol. 2006, 37, 1–9. [Google Scholar] [CrossRef]
- Husain, S.; Pietrangeli, C.E.; Zeevi, A. Delayed onset CMV disease in solid organ transplant recipients. Transpl. Immunol. 2009, 21, 1–9. [Google Scholar] [CrossRef]
- Limaye, A.P. Antiviral resistance in cytomegalovirus: An emerging problem in organ transplant recipients. Semin. Respir. Infect. 2002, 17, 265–273. [Google Scholar] [CrossRef]
- Karltorp, E.; Hellstrom, S.; Lewensohn-Fuchs, I.; Carlsson-Hansen, E.; Carlsson, P.I.; Engman, M.L. Congenital cytomegalovirus infection—A common cause of hearing loss of unknown aetiology. Acta Paediatr. 2012, 101, e357–e362. [Google Scholar] [CrossRef]
- Leung, A.K.; Sauve, R.S.; Davies, H.D. Congenital cytomegalovirus infection. J. Natl. Med. Assoc. 2003, 95, 213–218. [Google Scholar]
- Cannon, M.J.; Davis, K.F. Washing our hands of the congenital cytomegalovirus disease epidemic. BMC Publ. Health 2005, 5, 70. [Google Scholar] [CrossRef] [Green Version]
- Lurain, N.S.; Chou, S. Antiviral drug resistance of human cytomegalovirus. Clin. Microbiol. Rev. 2010, 23, 689–712. [Google Scholar] [CrossRef]
- Kalejta, R.F. Functions of human cytomegalovirus tegument proteins prior to immediate early gene expression. Curr. Top. Microbiol. Immunol. 2008, 325, 101–115. [Google Scholar]
- Kalejta, R.F. Tegument proteins of human cytomegalovirus. Microbiol. Mol. Biol. Rev. 2008, 72, 249–265. [Google Scholar] [CrossRef]
- Penkert, R.R.; Kalejta, R.F. Tale of a tegument transactivator: The past, present and future of human CMV pp71. Future Virol. 2012, 7, 855–869. [Google Scholar] [CrossRef]
- Compton, T.; Feire, A. Early events in human cytomegalovirus infection. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Arvin, A., Campadelli-Fiume, G., Mocarski, E., Moore, P.S., Roizman, B., Whitley, R., Yamanishi, K., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Varnum, S.M.; Streblow, D.N.; Monroe, M.E.; Smith, P.; Auberry, K.J.; Pasa-Tolic, L.; Wang, D.; Camp, D.G., 2nd; Rodland, K.; Wiley, S.; et al. Identification of proteins in human cytomegalovirus (HCMV) particles: The HCMV proteome. J. Virol. 2004, 78, 10960–10966. [Google Scholar] [CrossRef]
- Terhune, S.S.; Schroer, J.; Shenk, T. RNAs are packaged into human cytomegalovirus virions in proportion to their intracellular concentration. J. Virol. 2004, 78, 10390–10398. [Google Scholar] [CrossRef]
- Stinski, M.F.; Meier, J.L. Immediate-early viral gene regulation and function. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Arvin, A., Campadelli-Fiume, G., Mocarski, E., Moore, P.S., Roizman, B., Whitley, R., Yamanishi, K., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- White, E.A.; Spector, D.H. Early viral gene expression and function. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Arvin, A., Campadelli-Fiume, G., Mocarski, E., Moore, P.S., Roizman, B., Whitley, R., Yamanishi, K., Eds.; Cambridge University Press: Cambridge, UK, Cambridge, 2007. [Google Scholar]
- Anders, D.G.; Kerry, J.A.; Pari, G.S. DNA synthesis and late viral gene expression. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Arvin, A., Campadelli-Fiume, G., Mocarski, E., Moore, P.S., Roizman, B., Whitley, R., Yamanishi, K., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Tandon, R.; Mocarski, E.S. Viral and host control of cytomegalovirus maturation. Trends Microbiol. 2012, 20, 392–401. [Google Scholar] [CrossRef]
- Yu, X.; Shah, S.; Lee, M.; Dai, W.; Lo, P.; Britt, W.; Zhu, H.; Liu, F.; Zhou, Z.H. Biochemical and structural characterization of the capsid-bound tegument proteins of human cytomegalovirus. J. Struct. Biol. 2011, 174, 451–460. [Google Scholar] [CrossRef]
- Das, S.; Pellett, P.E. Spatial relationships between markers for secretory and endosomal machinery in human cytomegalovirus-infected cells versus those in uninfected cells. J. Virol. 2011, 85, 5864–5879. [Google Scholar] [CrossRef]
- Das, S.; Vasanji, A.; Pellett, P.E. Three-dimensional structure of the human cytomegalovirus cytoplasmic virion assembly complex includes a reoriented secretory apparatus. J. Virol. 2007, 81, 11861–11869. [Google Scholar] [CrossRef]
- Alwine, J.C. The human cytomegalovirus assembly compartment: A masterpiece of viral manipulation of cellular processes that facilitates assembly and egress. PLoS Pathog. 2012, 8, e1002878. [Google Scholar] [CrossRef]
- Winkler, M.; Stamminger, T. A specific subform of the human cytomegalovirus transactivator protein pUL69 is contained within the tegument of virus particles. J. Virol. 1996, 70, 8984–8987. [Google Scholar]
- Jones, T.R.; Lee, S.W. An acidic cluster of human cytomegalovirus UL99 tegument protein is required for trafficking and function. J. Virol. 2004, 78, 1488–1502. [Google Scholar] [CrossRef]
- Gallina, A.; Simoncini, L.; Garbelli, S.; Percivalle, E.; Pedrali-Noy, G.; Lee, K.S.; Erikson, R.L.; Plachter, B.; Gerna, G.; Milanesi, G. Polo-like kinase 1 as a target for human cytomegalovirus pp65 lower matrix protein. J. Virol. 1999, 73, 1468–1478. [Google Scholar]
- Van Zeijl, M.; Fairhurst, J.; Baum, E.Z.; Sun, L.; Jones, T.R. The human cytomegalovirus UL97 protein is phosphorylated and a component of virions. Virology 1997, 231, 72–80. [Google Scholar] [CrossRef]
- Michelson, S.; Turowski, P.; Picard, L.; Goris, J.; Landini, M.P.; Topilko, A.; Hemmings, B.; Bessia, C.; Garcia, A.; Virelizier, J.L. Human cytomegalovirus carries serine/threonine protein phosphatases PP1 and a host-cell derived PP2a. J. Virol. 1996, 70, 1415–1423. [Google Scholar]
- Sampaio, K.L.; Cavignac, Y.; Stierhof, Y.D.; Sinzger, C. Human cytomegalovirus labeled with green fluorescent protein for live analysis of intracellular particle movements. J. Virol. 2005, 79, 2754–2767. [Google Scholar] [CrossRef]
- Tandon, R.; Mocarski, E.S. Cytomegalovirus pUL96 is critical for the stability of pp150-associated nucleocapsids. J. Virol. 2011, 85, 7129–7141. [Google Scholar] [CrossRef]
- Meyer, H.H.; Ripalti, A.; Landini, M.P.; Radsak, K.; Kern, H.F.; Hensel, G.M. Human cytomegalovirus late-phase maturation is blocked by stably expressed UL32 antisense mrna in astrocytoma cells. J. Gen. Virol. 1997, 78, 2621–2631. [Google Scholar]
- Tandon, R.; Mocarski, E.S. Control of cytoplasmic maturation events by cytomegalovirus tegument protein pp150. J. Virol. 2008, 82, 9433–9444. [Google Scholar] [CrossRef]
- Moorman, N.J.; Sharon-Friling, R.; Shenk, T.; Cristea, I.M. A targeted spatial-temporal proteomics approach implicates multiple cellular trafficking pathways in human cytomegalovirus virion maturation. Mol. Cell. Proteomics 2010, 9, 851–860. [Google Scholar] [CrossRef]
- Trus, B.L.; Gibson, W.; Cheng, N.; Steven, A.C. Capsid structure of simian cytomegalovirus from cryoelectron microscopy: Evidence for tegument attachment sites. J. Virol. 1999, 73, 2181–2192. [Google Scholar]
- Phillips, S.L.; Bresnahan, W.A. Identification of binary interactions between human cytomegalovirus virion proteins. J. Virol. 2011, 85, 440–447. [Google Scholar] [CrossRef]
- To, A.; Bai, Y.; Shen, A.; Gong, H.; Umamoto, S.; Lu, S.; Liu, F. Yeast two hybrid analyses reveal novel binary interactions between human cytomegalovirus-encoded virion proteins. PLoS One 2011, 6, e17796. [Google Scholar]
- Patrone, M.; Percivalle, E.; Secchi, M.; Fiorina, L.; Pedrali-Noy, G.; Zoppe, M.; Baldanti, F.; Hahn, G.; Koszinowski, U.H.; Milanesi, G.; et al. The human cytomegalovirus UL45 gene product is a late, virion-associated protein and influences virus growth at low multiplicities of infection. J. Gen. Virol. 2003, 84, 3359–3370. [Google Scholar] [CrossRef]
- Dunn, W.; Chou, C.; Li, H.; Hai, R.; Patterson, D.; Stolc, V.; Zhu, H.; Liu, F. Functional profiling of a human cytomegalovirus genome. Proc. Natl. Acad. Sci. USA 2003, 100, 14223–14228. [Google Scholar] [CrossRef]
- Mettenleiter, T.C.; Klupp, B.G.; Granzow, H. Herpesvirus assembly: An update. Virus Res. 2009, 143, 222–234. [Google Scholar] [CrossRef]
- Gallina, A.; Percivalle, E.; Simoncini, L.; Revello, M.G.; Gerna, G.; Milanesi, G. Human cytomegalovirus pp65 lower matrix phosphoprotein harbours two transplantable nuclear localization signals. J. Gen. Virol. 1996, 77, 1151–1157. [Google Scholar] [CrossRef]
- Schmolke, S.; Drescher, P.; Jahn, G.; Plachter, B. Nuclear targeting of the tegument protein pp65 (UL83) of human cytomegalovirus: An unusual bipartite nuclear localization signal functions with other portions of the protein to mediate its efficient nuclear transport. J. Virol. 1995, 69, 1071–1078. [Google Scholar]
- Shen, W.; Westgard, E.; Huang, L.; Ward, M.D.; Osborn, J.L.; Chau, N.H.; Collins, L.; Marcum, B.; Koach, M.A.; Bibbs, J.; et al. Nuclear trafficking of the human cytomegalovirus pp71 (ppUL82) tegument protein. Virology 2008, 376, 42–52. [Google Scholar] [CrossRef]
- Munger, J.; Yu, D.; Shenk, T. UL26-deficient human cytomegalovirus produces virions with hypophosphorylated pp28 tegument protein that is unstable within newly infected cells. J. Virol. 2006, 80, 3541–3548. [Google Scholar] [CrossRef]
- Sanchez, V.; Sztul, E.; Britt, W.J. Human cytomegalovirus pp28 (UL99) localizes to a cytoplasmic compartment which overlaps the endoplasmic reticulum-golgi-intermediate compartment. J. Virol. 2000, 74, 3842–3851. [Google Scholar] [CrossRef]
- Womack, A.; Shenk, T. Human cytomegalovirus tegument protein pUL71 is required for efficient virion egress. mBio 2010, 1, e00282-10. [Google Scholar]
- Zini, N.; Santi, S.; Riccio, M.; Landini, M.P.; Battista, M.C.; Maraldi, N.M. pUL25 immunolocalization in human cytomegalovirus-infected and gene-transfected cells. Arch. Virol. 2000, 145, 795–803. [Google Scholar] [CrossRef]
- Frankenberg, N.; Lischka, P.; Pepperl-Klindworth, S.; Stamminger, T.; Plachter, B. Nucleocytoplasmic shuttling and CRM1-dependent MHC class I peptide presentation of human cytomegalovirus pp65. Med. Microbiol. Immunol. 2012, 201, 567–579. [Google Scholar] [CrossRef]
- Lischka, P.; Rosorius, O.; Trommer, E.; Stamminger, T. A novel transferable nuclear export signal mediates CRM1-independent nucleocytoplasmic shuttling of the human cytomegalovirus transactivator protein pUL69. EMBO J. 2001, 20, 7271–7283. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Z.; Zhao, X.; Wei, H.; Deng, J.; Cui, Z.; Zhang, X.E. Human cytomegalovirus UL94 is a nucleocytoplasmic shuttling protein containing two NLSs and one NES. Virus Res. 2012, 166, 31–42. [Google Scholar] [CrossRef]
- Sanchez, V.; Mahr, J.A.; Orazio, N.I.; Spector, D.H. Nuclear export of the human cytomegalovirus tegument protein pp65 requires cyclin-dependent kinase activity and the CRM1 exporter. J. Virol. 2007, 81, 11730–11736. [Google Scholar] [CrossRef]
- Goldberg, M.D.; Honigman, A.; Weinstein, J.; Chou, S.; Taraboulos, A.; Rouvinski, A.; Shinder, V.; Wolf, D.G. Human cytomegalovirus UL97 kinase and nonkinase functions mediate viral cytoplasmic secondary envelopment. J. Virol. 2011, 85, 3375–3384. [Google Scholar] [CrossRef]
- Prichard, M.N.; Britt, W.J.; Daily, S.L.; Hartline, C.B.; Kern, E.R. Human cytomegalovirus UL97 kinase is required for the normal intranuclear distribution of pp65 and virion morphogenesis. J. Virol. 2005, 79, 15494–15502. [Google Scholar] [CrossRef]
- Tandon, R.; AuCoin, D.P.; Mocarski, E.S. Human cytomegalovirus exploits ESCRT machinery in the process of virion maturation. J. Virol. 2009, 83, 10797–10807. [Google Scholar] [CrossRef]
- Yu, D.; Silva, M.C.; Shenk, T. Functional map of human cytomegalovirus AD169 defined by global mutational analysis. Proc. Natl. Acad. Sci. USA 2003, 100, 12396–12401. [Google Scholar] [CrossRef]
- Schierling, K.; Buser, C.; Mertens, T.; Winkler, M. Human cytomegalovirus tegument protein ppUL35 is important for viral replication and particle formation. J. Virol. 2005, 79, 3084–3096. [Google Scholar] [CrossRef]
- Bresnahan, W.A.; Hultman, G.E.; Shenk, T. Replication of wild-type and mutant human cytomegalovirus in life-extended human diploid fibroblasts. J. Virol. 2000, 74, 10816–10818. [Google Scholar] [CrossRef]
- Bresnahan, W.A.; Shenk, T.E. UL82 virion protein activates expression of immediate early viral genes in human cytomegalovirus-infected cells. Proc. Natl. Acad. Sci. USA 2000, 97, 14506–14511. [Google Scholar] [CrossRef]
- Schmolke, S.; Kern, H.F.; Drescher, P.; Jahn, G.; Plachter, B. The dominant phosphoprotein pp65 (UL83) of human cytomegalovirus is dispensable for growth in cell culture. J. Virol. 1995, 69, 5959–5968. [Google Scholar]
- Chevillotte, M.; Landwehr, S.; Linta, L.; Frascaroli, G.; Luske, A.; Buser, C.; Mertens, T.; von Einem, J. Major tegument protein pp65 of human cytomegalovirus is required for the incorporation of pUL69 and pUL97 into the virus particle and for viral growth in macrophages. J. Virol. 2009, 83, 2480–2490. [Google Scholar] [CrossRef]
- Cristea, I.M.; Moorman, N.J.; Terhune, S.S.; Cuevas, C.D.; O'Keefe, E.S.; Rout, M.P.; Chait, B.T.; Shenk, T. Human cytomegalovirus pUL83 stimulates activity of the viral immediate-early promoter through its interaction with the cellular IFI16 protein. J. Virol. 2010, 84, 7803–7814. [Google Scholar] [CrossRef]
- Phillips, S.L.; Bresnahan, W.A. The human cytomegalovirus (HCMV) tegument protein UL94 is essential for secondary envelopment of HCMV virions. J. Virol. 2012, 86, 2523–2532. [Google Scholar] [CrossRef]
- Prichard, M.N.; Gao, N.; Jairath, S.; Mulamba, G.; Krosky, P.; Coen, D.M.; Parker, B.O.; Pari, G.S. A recombinant human cytomegalovirus with a large deletion in UL97 has a severe replication deficiency. J. Virol. 1999, 73, 5663–5670. [Google Scholar]
- Blankenship, C.A.; Shenk, T. Mutant human cytomegalovirus lacking the immediate-early TRS1 coding region exhibits a late defect. J. Virol. 2002, 76, 12290–12299. [Google Scholar] [CrossRef]
- Silva, M.C.; Yu, Q.C.; Enquist, L.; Shenk, T. Human cytomegalovirus UL99-encoded pp28 is required for the cytoplasmic envelopment of tegument-associated capsids. J. Virol. 2003, 77, 10594–10605. [Google Scholar] [CrossRef]
- Baxter, M.K.; Gibson, W. Cytomegalovirus basic phosphoprotein (pUL32) binds to capsids in vitro through its amino one-third. J. Virol. 2001, 75, 6865–6873. [Google Scholar] [CrossRef]
- Dai, X.; Yu, X.; Gong, H.; Jiang, X.; Abenes, G.; Liu, H.; Shivakoti, S.; Britt, W.J.; Zhu, H.; Liu, F.; et al. The smallest capsid protein mediates binding of the essential tegument protein pp150 to stabilize DNA-containing capsids in human cytomegalovirus. PLoS Pathog. 2013, 9, e1003525. [Google Scholar] [CrossRef]
- Bogdanow, B.; Weisbach, H.; von Einem, J.; Straschewski, S.; Voigt, S.; Winkler, M.; Hagemeier, C.; Wiebusch, L. Human cytomegalovirus tegument protein pp150 acts as a cyclin A2-CDK-dependent sensor of the host cell cycle and differentiation state. Proc. Natl. Acad. Sci. USA 2013, 110, 17510–17515. [Google Scholar] [CrossRef]
- Hensel, G.; Meyer, H.; Gartner, S.; Brand, G.; Kern, H.F. Nuclear localization of the human cytomegalovirus tegument protein pp150 (ppUL32). J. Gen. Virol. 1995, 76, 1591–1601. [Google Scholar] [CrossRef]
- Sanchez, V.; Greis, K.D.; Sztul, E.; Britt, W.J. Accumulation of virion tegument and envelope proteins in a stable cytoplasmic compartment during human cytomegalovirus replication: Characterization of a potential site of virus assembly. J. Virol. 2000, 74, 975–986. [Google Scholar] [CrossRef]
- Indran, S.V.; Ballestas, M.E.; Britt, W.J. Bicaudal D1-dependent trafficking of human cytomegalovirus tegument protein pp150 in virus-infected cells. J. Virol. 2010, 84, 3162–3177. [Google Scholar] [CrossRef]
- Dienstbier, M.; Li, X. Bicaudal-D and its role in cargo sorting by microtubule-based motors. Biochem. Soc. Trans. 2009, 37, 1066–1071. [Google Scholar] [CrossRef]
- Short, B.; Haas, A.; Barr, F.A. Golgins and GTPases, giving identity and structure to the golgi apparatus. Biochim. Biophys. Acta 2005, 1744, 383–395. [Google Scholar]
- Indran, S.V.; Britt, W.J. A role for the small GTPase Rab6 in assembly of human cytomegalovirus. J. Virol. 2011, 85, 5213–5219. [Google Scholar] [CrossRef]
- Gibson, W. Structure and assembly of the virion. Intervirology 1996, 39, 389–400. [Google Scholar]
- Brock, I.; Kruger, M.; Mertens, T.; von Einem, J. Nuclear targeting of human cytomegalovirus large tegument protein pUL48 is essential for viral growth. J. Virol. 2013, 87, 6005–6019. [Google Scholar] [CrossRef]
- Bechtel, J.T.; Shenk, T. Human cytomegalovirus UL47 tegument protein functions after entry and before immediate-early gene expression. J. Virol. 2002, 76, 1043–1050. [Google Scholar] [CrossRef]
- Kim, E.T.; Oh, S.E.; Lee, Y.O.; Gibson, W.; Ahn, J.H. Cleavage specificity of the UL48 deubiquitinating protease activity of human cytomegalovirus and the growth of an active-site mutant virus in cultured cells. J. Virol. 2009, 83, 12046–12056. [Google Scholar] [CrossRef]
- Wang, J.; Loveland, A.N.; Kattenhorn, L.M.; Ploegh, H.L.; Gibson, W. High-molecular-weight protein (pUL48) of human cytomegalovirus is a competent deubiquitinating protease: Mutant viruses altered in its active-site cysteine or histidine are viable. J. Virol. 2006, 80, 6003–6012. [Google Scholar] [CrossRef]
- Ogawa-Goto, K.; Irie, S.; Omori, A.; Miura, Y.; Katano, H.; Hasegawa, H.; Kurata, T.; Sata, T.; Arao, Y. An endoplasmic reticulum protein, p180, is highly expressed in human cytomegalovirus-permissive cells and interacts with the tegument protein encoded by UL48. J. Virol. 2002, 76, 2350–2362. [Google Scholar] [CrossRef]
- Desai, P.J. A null mutation in the UL36 gene of herpes simplex virus type 1 results in accumulation of unenveloped DNA-filled capsids in the cytoplasm of infected cells. J. Virol. 2000, 74, 11608–11618. [Google Scholar] [CrossRef]
- Sandbaumhuter, M.; Dohner, K.; Schipke, J.; Binz, A.; Pohlmann, A.; Sodeik, B.; Bauerfeind, R. Cytosolic herpes simplex virus capsids not only require binding inner tegument protein pUL36 but also pUL37 for active transport prior to secondary envelopment. Cell. Microbiol. 2013, 15, 248–269. [Google Scholar] [CrossRef]
- Schipke, J.; Pohlmann, A.; Diestel, R.; Binz, A.; Rudolph, K.; Nagel, C.H.; Bauerfeind, R.; Sodeik, B. The C terminus of the large tegument protein pUL36 contains multiple capsid binding sites that function differently during assembly and cell entry of herpes simplex virus. J. Virol. 2012, 86, 3682–3700. [Google Scholar] [CrossRef]
- Hensel, G.M.; Meyer, H.H.; Buchmann, I.; Pommerehne, D.; Schmolke, S.; Plachter, B.; Radsak, K.; Kern, H.F. Intracellular localization and expression of the human cytomegalovirus matrix phosphoprotein pp71 (ppUL82): Evidence for its translocation into the nucleus. J. Gen. Virol. 1996, 77, 3087–3097. [Google Scholar] [CrossRef]
- Revello, M.G.; Percivalle, E.; Di Matteo, A.; Morini, F.; Gerna, G. Nuclear expression of the lower matrix protein of human cytomegalovirus in peripheral blood leukocytes of immunocompromised viraemic patients. J. Gen. Virol. 1992, 73, 437–442. [Google Scholar] [CrossRef]
- Lorz, K.; Hofmann, H.; Berndt, A.; Tavalai, N.; Mueller, R.; Schlotzer-Schrehardt, U.; Stamminger, T. Deletion of open reading frame UL26 from the human cytomegalovirus genome results in reduced viral growth, which involves impaired stability of viral particles. J. Virol. 2006, 80, 5423–5434. [Google Scholar] [CrossRef]
- Liljeroos, L.; Butcher, S.J. Matrix proteins as centralized organizers of negative-sense RNA virions. Front. Biosci. 2013, 18, 696–715. [Google Scholar] [CrossRef]
- Liu, Y.; Biegalke, B.J. The human cytomegalovirus UL35 gene encodes two proteins with different functions. J. Virol. 2002, 76, 2460–2468. [Google Scholar] [CrossRef]
- Schierling, K.; Stamminger, T.; Mertens, T.; Winkler, M. Human cytomegalovirus tegument proteins ppUL82 (pp71) and ppUL35 interact and cooperatively activate the major immediate-early enhancer. J. Virol. 2004, 78, 9512–9523. [Google Scholar] [CrossRef]
- Salsman, J.; Wang, X.; Frappier, L. Nuclear body formation and PML body remodeling by the human cytomegalovirus protein UL35. Virology 2011, 414, 119–129. [Google Scholar] [CrossRef]
- Tavalai, N.; Stamminger, T. Interplay between herpesvirus infection and host defense by PML nuclear bodies. Viruses 2009, 1, 1240–1264. [Google Scholar] [CrossRef]
- Salsman, J.; Jagannathan, M.; Paladino, P.; Chan, P.K.; Dellaire, G.; Raught, B.; Frappier, L. Proteomic profiling of the human cytomegalovirus UL35 gene products reveals a role for UL35 in the DNA repair response. J. Virol. 2012, 86, 806–820. [Google Scholar]
- Craighead, J.E.; Kanich, R.E.; Almeida, J.D. Nonviral microbodies with viral antigenicity produced in cytomegalovirus-infected cells. J. Virol. 1972, 10, 766–775. [Google Scholar]
- Weiland, K.L.; Oien, N.L.; Homa, F.; Wathen, M.W. Functional analysis of human cytomegalovirus polymerase accessory protein. Virus Res. 1994, 34, 191–206. [Google Scholar] [CrossRef]
- Chee, M.; Rudolph, S.A.; Plachter, B.; Barrell, B.; Jahn, G. Identification of the major capsid protein gene of human cytomegalovirus. J. Virol. 1989, 63, 1345–1353. [Google Scholar]
- He, Z.; He, Y.S.; Kim, Y.; Chu, L.; Ohmstede, C.; Biron, K.K.; Coen, D.M. The human cytomegalovirus UL97 protein is a protein kinase that autophosphorylates on serines and threonines. J. Virol. 1997, 71, 405–411. [Google Scholar]
- Milbradt, J.; Auerochs, S.; Marschall, M. Cytomegaloviral proteins pUL50 and pUL53 are associated with the nuclear lamina and interact with cellular protein kinase C. J. Gen. Virol. 2007, 88, 2642–2650. [Google Scholar] [CrossRef]
- Borst, E.M.; Wagner, K.; Binz, A.; Sodeik, B.; Messerle, M. The essential human cytomegalovirus gene UL52 is required for cleavage-packaging of the viral genome. J. Virol. 2008, 82, 2065–2078. [Google Scholar]
- Bogner, E.; Radsak, K.; Stinski, M.F. The gene product of human cytomegalovirus open reading frame UL56 binds the pac motif and has specific nuclease activity. J. Virol. 1998, 72, 2259–2264. [Google Scholar]
- Baldick, C.J., Jr.; Shenk, T. Proteins associated with purified human cytomegalovirus particles. J. Virol. 1996, 70, 6097–6105. [Google Scholar]
- Lee, S.H.; Kalejta, R.F.; Kerry, J.; Semmes, O.J.; O’Connor, C.M.; Khan, Z.; Garcia, B.A.; Shenk, T.; Murphy, E. BclAF1 restriction factor is neutralized by proteasomal degradation and microRNA repression during human cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2012, 109, 9575–9580. [Google Scholar] [CrossRef]
- Becke, S.; Fabre-Mersseman, V.; Aue, S.; Auerochs, S.; Sedmak, T.; Wolfrum, U.; Strand, D.; Marschall, M.; Plachter, B.; Reyda, S. Modification of the major tegument protein pp65 of human cytomegalovirus inhibits virus growth and leads to the enhancement of a protein complex with pUL69 and pUL97 in infected cells. J. Gen. Virol. 2010, 91, 2531–2541. [Google Scholar] [CrossRef]
- Kamil, J.P.; Coen, D.M. Human cytomegalovirus protein kinase UL97 forms a complex with the tegument phosphoprotein pp65. J. Virol. 2007, 81, 10659–10668. [Google Scholar] [CrossRef]
- Hayashi, M.L.; Blankenship, C.; Shenk, T. Human cytomegalovirus UL69 protein is required for efficient accumulation of infected cells in the G1 phase of the cell cycle. Proc. Natl. Acad. Sci. USA 2000, 97, 2692–2696. [Google Scholar] [CrossRef]
- Toth, Z.; Stamminger, T. The human cytomegalovirus regulatory protein UL69 and its effect on mrna export. Front. Biosci. 2008, 13, 2939–2949. [Google Scholar] [CrossRef]
- Cui, Z.; Zhang, K.; Zhang, Z.; Liu, Y.; Zhou, Y.; Wei, H.; Zhang, X.E. Visualization of the dynamic multimerization of human cytomegalovirus pp65 in punctuate nuclear foci. Virology 2009, 392, 169–177. [Google Scholar] [CrossRef]
- Wing, B.A.; Lee, G.C.; Huang, E.S. The human cytomegalovirus UL94 open reading frame encodes a conserved herpesvirus capsid/tegument-associated virion protein that is expressed with true late kinetics. J. Virol. 1996, 70, 3339–3345. [Google Scholar]
- Phillips, S.L.; Cygnar, D.; Thomas, A.; Bresnahan, W.A. Interaction between the human cytomegalovirus tegument proteins UL94 and UL99 is essential for virus replication. J. Virol. 2012, 86, 9995–10005. [Google Scholar] [CrossRef]
- Seo, J.Y.; Britt, W.J. Sequence requirements for localization of human cytomegalovirus tegument protein pp28 to the virus assembly compartment and for assembly of infectious virus. J. Virol. 2006, 80, 5611–5626. [Google Scholar] [CrossRef]
- Littler, E.; Stuart, A.D.; Chee, M.S. Human cytomegalovirus UL97 open reading frame encodes a protein that phosphorylates the antiviral nucleoside analogue ganciclovir. Nature 1992, 358, 160–162. [Google Scholar] [CrossRef]
- Prichard, M.N. Function of human cytomegalovirus UL97 kinase in viral infection and its inhibition by maribavir. Rev. Med. Virol. 2009, 19, 215–229. [Google Scholar] [CrossRef]
- Sharma, M.; Kamil, J.P.; Coughlin, M.; Reim, N.I.; Coen, D.M. Human cytomegalovirus UL50 and UL53 recruit viral protein kinase UL97, not protein kinase C, for disruption of nuclear lamina and nuclear egress in infected cells. J. Virol. 2014, 88, 249–262. [Google Scholar] [CrossRef]
- Marschall, M.; Marzi, A.; aus dem Siepen, P.; Jochmann, R.; Kalmer, M.; Auerochs, S.; Lischka, P.; Leis, M.; Stamminger, T. Cellular p32 recruits cytomegalovirus kinase pUL97 to redistribute the nuclear lamina. J. Biol. Chem. 2005, 280, 33357–33367. [Google Scholar] [CrossRef]
- Azzeh, M.; Honigman, A.; Taraboulos, A.; Rouvinski, A.; Wolf, D.G. Structural changes in human cytomegalovirus cytoplasmic assembly sites in the absence of UL97 kinase activity. Virology 2006, 354, 69–79. [Google Scholar] [CrossRef]
- Chee, M.S.; Bankier, A.T.; Beck, S.; Bohni, R.; Brown, C.M.; Cerny, R.; Horsnell, T.; Hutchison, C.A., 3rd; Kouzarides, T.; Martignetti, J.A.; et al. Analysis of the protein-coding content of the sequence of human cytomegalovirus strain AD169. Curr. Top. Microbiol. Immunol. 1990, 154, 125–169. [Google Scholar]
- Adamo, J.E.; Schroer, J.; Shenk, T. Human cytomegalovirus TRS1 protein is required for efficient assembly of DNA-containing capsids. J. Virol. 2004, 78, 10221–10229. [Google Scholar] [CrossRef]
- Marshall, E.E.; Geballe, A.P. Multifaceted evasion of the interferon response by cytomegalovirus. J. Interferon Cytokine Res. 2009, 29, 609–619. [Google Scholar] [CrossRef]
- Hakki, M.; Marshall, E.E.; de Niro, K.L.; Geballe, A.P. Binding and nuclear relocalization of protein kinase R by human cytomegalovirus TRS1. J. Virol. 2006, 80, 11817–11826. [Google Scholar] [CrossRef]
- Strang, B.L.; Geballe, A.P.; Coen, D.M. Association of human cytomegalovirus proteins IRS1 and TRS1 with the viral DNA polymerase accessory subunit UL44. J. Gen. Virol. 2010, 91, 2167–2175. [Google Scholar] [CrossRef]
- Strang, B.L.; Bender, B.J.; Sharma, M.; Pesola, J.M.; Sanders, R.L.; Spector, D.H.; Coen, D.M. A mutation deleting sequences encoding the amino terminus of human cytomegalovirus UL84 impairs interaction with UL44 and capsid localization. J. Virol. 2012, 86, 11066–11077. [Google Scholar] [CrossRef]
- Schauflinger, M.; Fischer, D.; Schreiber, A.; Chevillotte, M.; Walther, P.; Mertens, T.; von Einem, J. The tegument protein UL71 of human cytomegalovirus is involved in late envelopment and affects multivesicular bodies. J. Virol. 2011, 85, 3821–3832. [Google Scholar] [CrossRef]
- Meissner, C.S.; Suffner, S.; Schauflinger, M.; von Einem, J.; Bogner, E. A leucine zipper motif of a tegument protein triggers final envelopment of human cytomegalovirus. J. Virol. 2012, 86, 3370–3382. [Google Scholar] [CrossRef]
- Borst, E.M.; Kleine-Albers, J.; Gabaev, I.; Babic, M.; Wagner, K.; Binz, A.; Degenhardt, I.; Kalesse, M.; Jonjic, S.; Bauerfeind, R.; et al. The human cytomegalovirus UL51 protein is essential for viral genome cleavage-packaging and interacts with the terminase subunits pUL56 and pul89. J. Virol. 2013, 87, 1720–1732. [Google Scholar] [CrossRef]
- Britt, W.J.; Jarvis, M.; Seo, J.Y.; Drummond, D.; Nelson, J. Rapid genetic engineering of human cytomegalovirus by using a lambda phage linear recombination system: Demonstration that pp28 (UL99) is essential for production of infectious virus. J. Virol. 2004, 78, 539–543. [Google Scholar] [CrossRef]
- Seo, J.Y.; Britt, W.J. Cytoplasmic envelopment of human cytomegalovirus requires the postlocalization function of tegument protein pp28 within the assembly compartment. J. Virol. 2007, 81, 6536–6547. [Google Scholar] [CrossRef]
- Seo, J.Y.; Britt, W.J. Multimerization of tegument protein pp28 within the assembly compartment is required for cytoplasmic envelopment of human cytomegalovirus. J. Virol. 2008, 82, 6272–6287. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Smith, R.M.; Kosuri, S.; Kerry, J.A. Role of Human Cytomegalovirus Tegument Proteins in Virion Assembly. Viruses 2014, 6, 582-605. https://doi.org/10.3390/v6020582
Smith RM, Kosuri S, Kerry JA. Role of Human Cytomegalovirus Tegument Proteins in Virion Assembly. Viruses. 2014; 6(2):582-605. https://doi.org/10.3390/v6020582
Chicago/Turabian StyleSmith, Rebecca Marie, Srivenkat Kosuri, and Julie Anne Kerry. 2014. "Role of Human Cytomegalovirus Tegument Proteins in Virion Assembly" Viruses 6, no. 2: 582-605. https://doi.org/10.3390/v6020582
APA StyleSmith, R. M., Kosuri, S., & Kerry, J. A. (2014). Role of Human Cytomegalovirus Tegument Proteins in Virion Assembly. Viruses, 6(2), 582-605. https://doi.org/10.3390/v6020582