Characterization of Juquitiba Virus in Oligoryzomys fornesi from Brazilian Cerrado

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
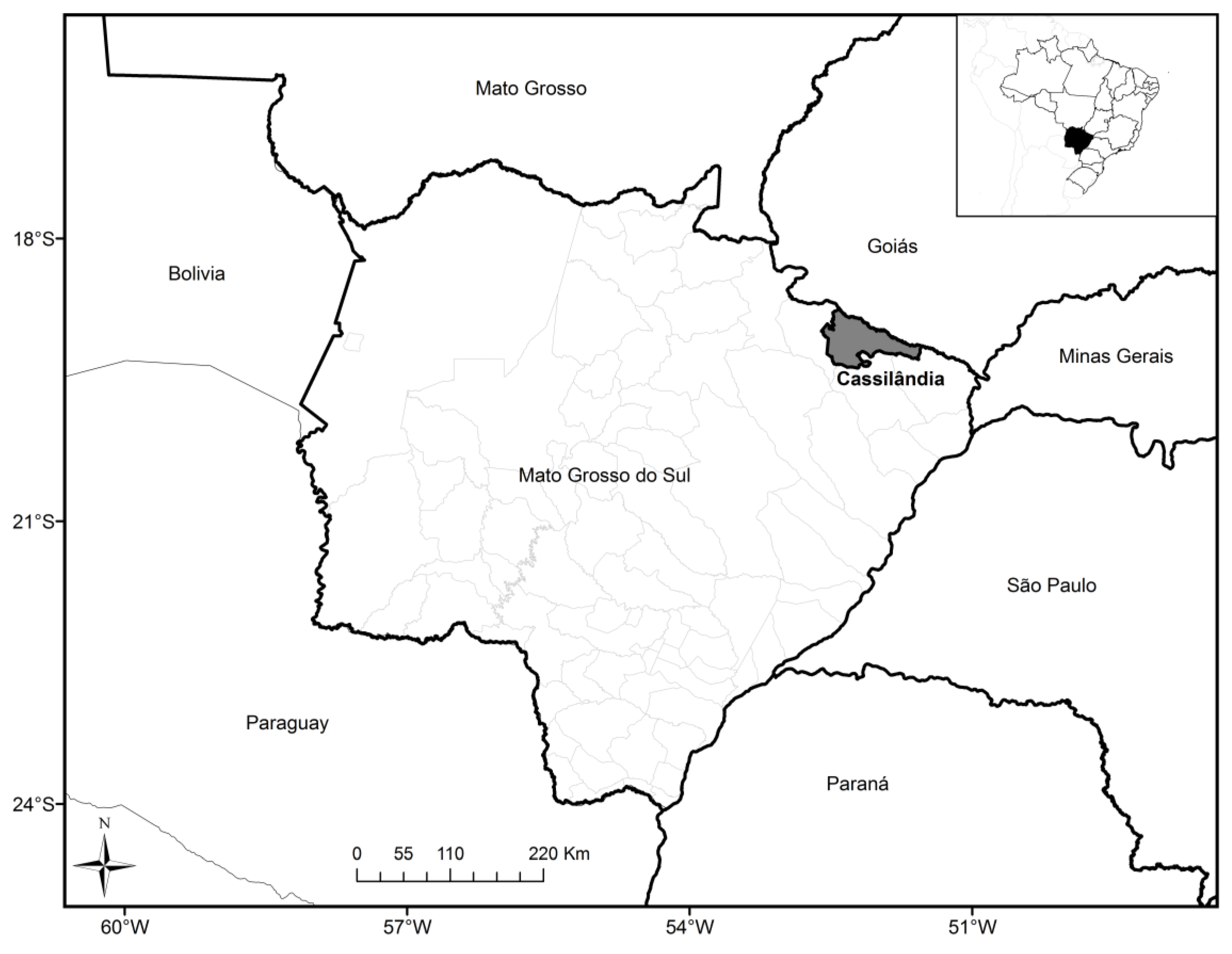
2. Experimental
Ethics Statement

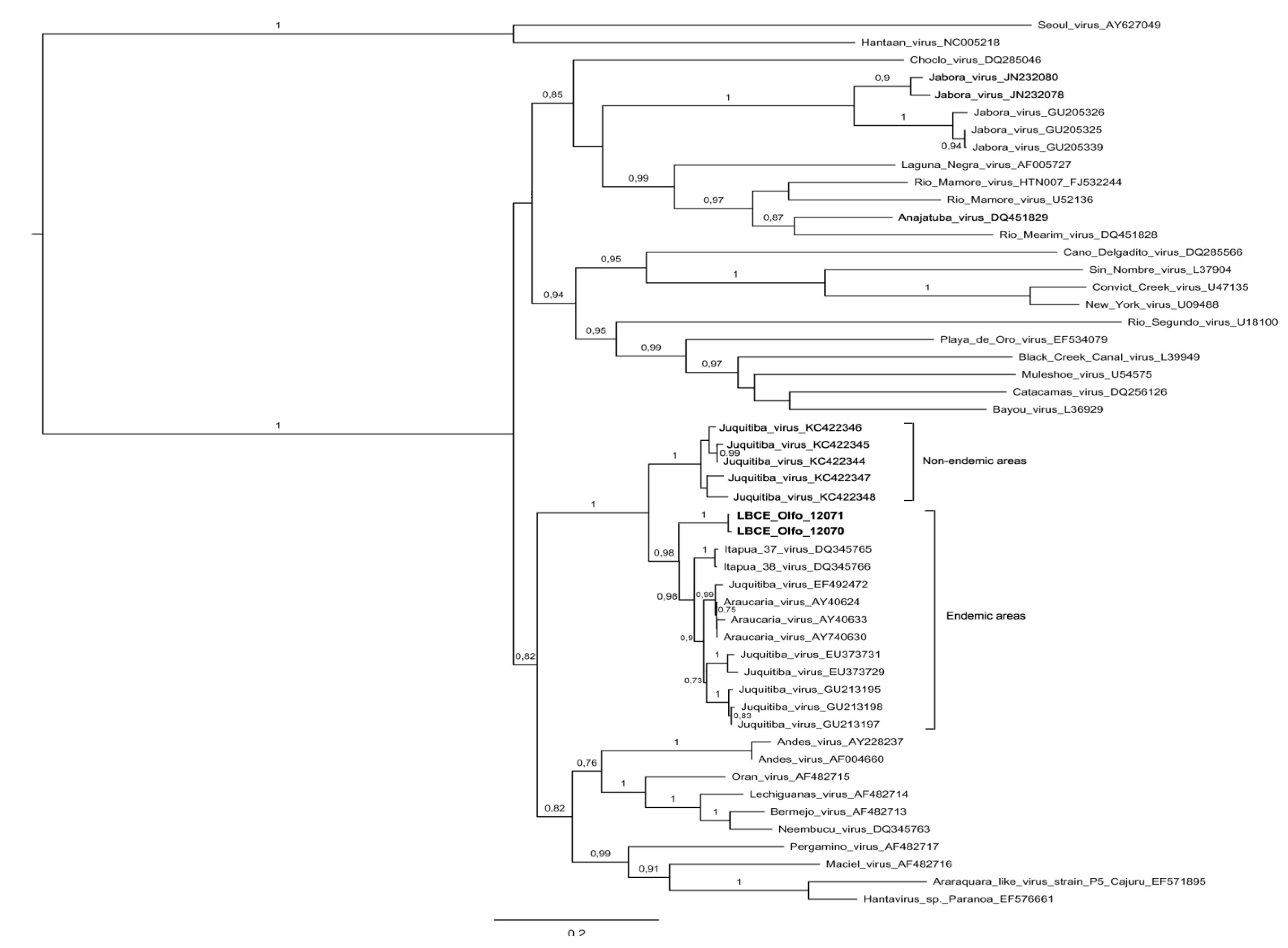
3. Results and Discussion


4. Conclusions
Acknowledgements
Author Contributions
References and Notes
- Jonsson, C.B.; Figueiredo, L.T.M.; Vapalahti, O.A. Global perspective on hantavirus ecology, epidemiology, and disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef]
- Watson, D.C.; Sargianou, M.; Papa, A.; Chra, P.; Starakis, I.; Panos, G. Epidemiology of Hantavirus infections in humans: A comprehensive, global overview. Crit. Rev. Microbiol. 2013, 7828, 1–12. [Google Scholar]
- Hjelle, B.; Torres-Pérez, F. Hantaviruses in the americas and their role as emerging pathogens. Viruses 2010, 2, 2559–2586. [Google Scholar] [CrossRef]
- Mills, J.N. Biodiversity loss and emerging infectious disease: An example from the rodent-borne hemorrhagic fevers. Biodiversity 2006, 7, 9–17. [Google Scholar] [CrossRef]
- Musser, G.G.; Carleton, M.D. Superfamily muroidea. In Mammal Species of the World: A Taxonomic and Geographic Reference; Wilson, D.E., Reeder, D.M., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2005; pp. 849–1531. [Google Scholar]
- Childs, J.E.; Ksiazek, T.G.; Spiropoulou, C.F.; Krebs, J.W.; Morzunov, S.; Maupin, G.O.; Gage, K.L.; Rollin, P.E.; Sarisky, J.; Enscore, R.E. Serologic and genetic identification of Peromyscus maniculatus as the primary rodent reservoir for a new hantavirus in the southwestern United States. J. Infect. Dis. 1994, 169, 1271–1280. [Google Scholar] [CrossRef]
- Delfraro, A.; Tomé, L.; D’Elía, G.; Clara, M.; Achával, F.; Russi, J.C.; Rodonz, J.R.A. Juquitiba-like hantavirus from 2 nonrelated rodent species, Uruguay. Emerg. Infect. Dis. 2008, 14, 1447–1451. [Google Scholar] [CrossRef]
- De Oliveira, R.C.; Guterres, A.; Schrago, C.G.; Fernandes, J.; Teixeira, B.R.; Zeccer, S.; Bonvicino, C.R.; D’Andrea, P.S.; de Lemos, E.R.S. Detection of the first incidence of Akodon paranaensis naturally infected with the Jabora virus strain (Hantavirus) in Brazil. Mem. Inst. Oswaldo Cruz 2012, 107, 424–428. [Google Scholar] [CrossRef]
- Raboni, S.M.; Hoffmann, F.G.; Oliveira, R.C.; Teixeira, B.R.; Bonvicino, C.R.; Stella, V.; Carstensen, S.; Bordignon, J.; D’Andrea, P.S.; Lemos, E.R.S.; et al. Phylogenetic characterization of hantaviruses from wild rodents and hantavirus pulmonary syndrome cases in the state of Paraná (southern Brazil). J. Gen. Virol. 2009, 90, 2166–2171. [Google Scholar] [CrossRef]
- Rosa, E.S.T.; Mills, J.N.; Padula, P.J.; Elkhoury, M.R.; Ksiazek, T.G.; Mendes, W.S.; Santos, E.D.; Araújo, G.C.B.; Martinez, V.P.; Rosa, J.F.S.T.; et al. Newly recognized hantaviruses associated with hantavirus pulmonary syndrome in northern Brazil: Partial genetic characterization of viruses and serologic implication of likely reservoirs. Vector Borne Zoonotic Dis. 2005, 5, 11–19. [Google Scholar] [CrossRef]
- Guterres, A.; de Oliveira, R.C.; Fernandes, J.; D’Andrea, P.S.; Bonvicino, C.R.; Bragagnolo, C.; Guimarães, G.D.; Almada, G.L.; Machado, R.R.; Lavocat, M.; et al. Phylogenetic analysis of the S segment from Juquitiba hantavirus: Identification of two distinct lineages in Oligoryzomys nigripes. Infect. Genet. Evol. 2013, 18, 262–268. [Google Scholar] [CrossRef]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Anisimova, M.; Gascuel, O. Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef]
- Chu, Y.K.; Goodin, D.; Owen, R.D.; Koch, D.; Jonsson, C.B. Sympatry of 2 hantavirus strains, paraguay, 2003–2007. Emerg. Infect. Dis. 2009, 15, 1977–1980. [Google Scholar] [CrossRef]
- Chu, Y.-K.; Owen, R.D.; Jonsson, C.B. Phylogenetic exploration of hantaviruses in Paraguay reveals reassortment and host switching in South America. Virol. J. 2011, 8. [Google Scholar] [CrossRef]
- Weksler, M.; Bonvicino, C.R. Taxonomy of pigmy rice rats genus oligoryzomys banGS, 1900 (Rodentia, Sigmodontinae ) of the Brazilian cerrado. Arq. do Mus. Nac. 2005, 63, 113–130. [Google Scholar]
- De Oliveira, R.C.; Padula, P.J.; Gomes, R.; Martinez, V.P.; Bellomo, C.; Bonvicino, C.R.; Freire e Lima, D.I.; Bragagnolo, C.; Caldas, A.C.S.; D’Andrea, P.S.; et al. Genetic characterization of hantaviruses associated with sigmodontine rodents in an endemic area for hantavirus pulmonary syndrome in southern Brazil. Vector Borne Zoonotic Dis. 2011, 11, 301–314. [Google Scholar] [CrossRef]
- Lin, X.-D.; Wang, W.; Guo, W.-P.; Zhang, X.-H.; Xing, J.-G.; Chen, S.-Z.; Li, M.-H.; Chen, Y.; Xu, J.; Plyusnin, A.; et al. Cross-species transmission in the speciation of the currently known murinae-associated hantaviruses. J. Virol. 2012, 86, 11171–11182. [Google Scholar] [CrossRef]
- Guo, W.-P.; Lin, X.-D.; Wang, W.; Tian, J.-H.; Cong, M.-L.; Zhang, H.-L.; Wang, M.-R.; Zhou, R.-H.; Wang, J.-B.; Li, M.-H.; et al. Phylogeny and origins of hantaviruses harbored by bats, insectivores, and rodents. PLoS Pathog. 2013, 9, e1003159. [Google Scholar] [CrossRef]
- Yadav, P.D.; Vincent, M.J.; Nichol, S.T. Thottapalayam virus is genetically distant to the rodent-borne hantaviruses, consistent with its isolation from the Asian house shrew (Suncus murinus). Virol. J. 2007, 4. [Google Scholar] [CrossRef]
- Lloyd-Smith, J.O.; George, D.; Pepin, K.M.; Pitzer, V.E.; Pulliam, J.R.C.; Dobson, A.P.; Hudson, P.J.; Grenfell, B.T. Epidemic dynamics at the human-animal interface. Science 2009, 326, 1362–1367. [Google Scholar] [CrossRef]
- Allen, L.J.S.; Wesley, C.L.; Owen, R.D.; Goodin, D.G.; Koch, D.; Jonsson, C.B.; Chu, Y.-K.; Shawn Hutchinson, J.M.; Paige, R.L. A habitat-based model for the spread of hantavirus between reservoir and spillover species. J. Theor. Biol. 2009, 260, 510–522. [Google Scholar] [CrossRef]
- Johnson, A.M.; Bowen, M.D.; Ksiazek, T.G.; Williams, R.J.; Bryan, R.T.; Mills, J.N.; Peters, C.J.; Nichol, S.T. Laguna Negra virus associated with HPS in western Paraguay and Bolivia. Virology 1997, 238, 115–127. [Google Scholar] [CrossRef]
- Levis, S.; Garcia, J.; Pini, N.; Calderón, G.; Ramírez, J.; Bravo, D.; St Jeor, S.; Ripoll, C.; Bego, M.; Lozano, E.; et al. Hantavirus pulmonary syndrome in northwestern Argentina: Circulation of Laguna Negra virus associated with Calomys callosus. Am. J. Trop. Med. Hyg. 2004, 71, 658–663. [Google Scholar]
- Travassos Da Rosa, E.S.; Medeiros, D.B.A.; Nunes, M.R.T.; Simith, D.B.; Pereira, A.D.S.; Elkhoury, M.R.; Santos, E.D.; Lavocat, M.; Marques, A.A.; Via, A.V.G.; et al. Molecular epidemiology of Laguna Negra virus, Mato Grosso State, Brazil. Emerg. Infect. Dis. 2012, 18, 982–985. [Google Scholar] [CrossRef]
- D’Elía, G.; Luna, L.; González, E.M.; Patterson, B.D. On the Sigmodontinae radiation (Rodentia, Cricetidae): An appraisal of the phylogenetic position of Rhagomys. Mol. Phylogenet. Evol. 2006, 38, 558–564. [Google Scholar] [CrossRef]
- Parada, A.; Pardiñas, U.F.J.; Salazar-Bravo, J.; D’Elía, G.; Palma, R.E. Dating an impressive Neotropical radiation: Molecular time estimates for the Sigmodontinae (Rodentia) provide insights into its historical biogeography. Mol. Phylogenet. Evol. 2013, 66, 960–968. [Google Scholar] [CrossRef]
- Miranda, G.B.; Oliveira, L.F.B.; Andrades-Miranda, J.; Langguth, A.; Callegari-Jacques, S.M.; Mattevi, M.S. Phylogenetic and phylogeographic patterns in sigmodontine rodents of the genus oligoryzomys. J. Hered. 2009, 100, 309–321. [Google Scholar] [CrossRef]
- Raboni, S.M.; Delfraro, A.; de Borba, L.; Teixeira, B.R.; Stella, V.; de Araujo, M.R.; Carstensen, S.; Rubio, G.; Maron, A.; Lemos, E.R.; et al. Hantavirus infection prevalence in wild rodents and human anti-hantavirus serological profiles from different geographic areas of South Brazil. Am. J. Trop. Med. Hyg. 2012, 87, 371–378. [Google Scholar] [CrossRef]
- Ramsden, C.; Melo, F.L.; Figueiredo, L.M.; Holmes, E.C.; Zanotto, P.M. A high rates of molecular evolution in hantaviruses. Mol. Biol. Evol. 2008, 25, 1488–1492. [Google Scholar] [CrossRef]
- Oliveira, R.C.; Teixeira, B.R.; Mello, F.C.; Pereira, P.; Duarte, S.; Bonaldo, M.C.; Bonvicino, C.R.; D’Andrea, P.S.; Lemos, E.R.S. Genetic characterization of a Juquitiba-like viral lineage in Oligoryzomys nigripes in Rio de Janeiro, Brazil. Acta Trop. 2009, 112, 212–218. [Google Scholar] [CrossRef]
- Padula, P.; Martinez, V.P.; Bellomo, C.; Maidana, S.; San Juan, J.; Tagliaferri, P.; Bargardi, S.; Vazquez, C.; Colucci, N.; Estévez, J.; et al. Pathogenic hantaviruses, northeastern Argentina and eastern Paraguay. Emerg. Infect. Dis. 2007, 13, 1211–1214. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Guterres, A.; De Oliveira, R.C.; Fernandes, J.; Strecht, L.; Casado, F.; Gomes de Oliveira, F.C.; D'Andrea, P.S.; Bonvicino, C.R.; Schrago, C.G.; Sampaio de Lemos, E.R. Characterization of Juquitiba Virus in Oligoryzomys fornesi from Brazilian Cerrado. Viruses 2014, 6, 1473-1482. https://doi.org/10.3390/v6041473
Guterres A, De Oliveira RC, Fernandes J, Strecht L, Casado F, Gomes de Oliveira FC, D'Andrea PS, Bonvicino CR, Schrago CG, Sampaio de Lemos ER. Characterization of Juquitiba Virus in Oligoryzomys fornesi from Brazilian Cerrado. Viruses. 2014; 6(4):1473-1482. https://doi.org/10.3390/v6041473
Chicago/Turabian StyleGuterres, Alexandro, Renata Carvalho De Oliveira, Jorlan Fernandes, Liana Strecht, Flavia Casado, Flavio Cesar Gomes de Oliveira, Paulo Sérgio D'Andrea, Cibele Rodrigues Bonvicino, Carlos Guerra Schrago, and Elba Regina Sampaio de Lemos. 2014. "Characterization of Juquitiba Virus in Oligoryzomys fornesi from Brazilian Cerrado" Viruses 6, no. 4: 1473-1482. https://doi.org/10.3390/v6041473
APA StyleGuterres, A., De Oliveira, R. C., Fernandes, J., Strecht, L., Casado, F., Gomes de Oliveira, F. C., D'Andrea, P. S., Bonvicino, C. R., Schrago, C. G., & Sampaio de Lemos, E. R. (2014). Characterization of Juquitiba Virus in Oligoryzomys fornesi from Brazilian Cerrado. Viruses, 6(4), 1473-1482. https://doi.org/10.3390/v6041473