HIV-1 Env-Specific Memory and Germinal Center B Cells in C57BL/6 Mice

Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Serological Assessment of Env-Specific Responses in BALB/c and C57BL/6 Mice


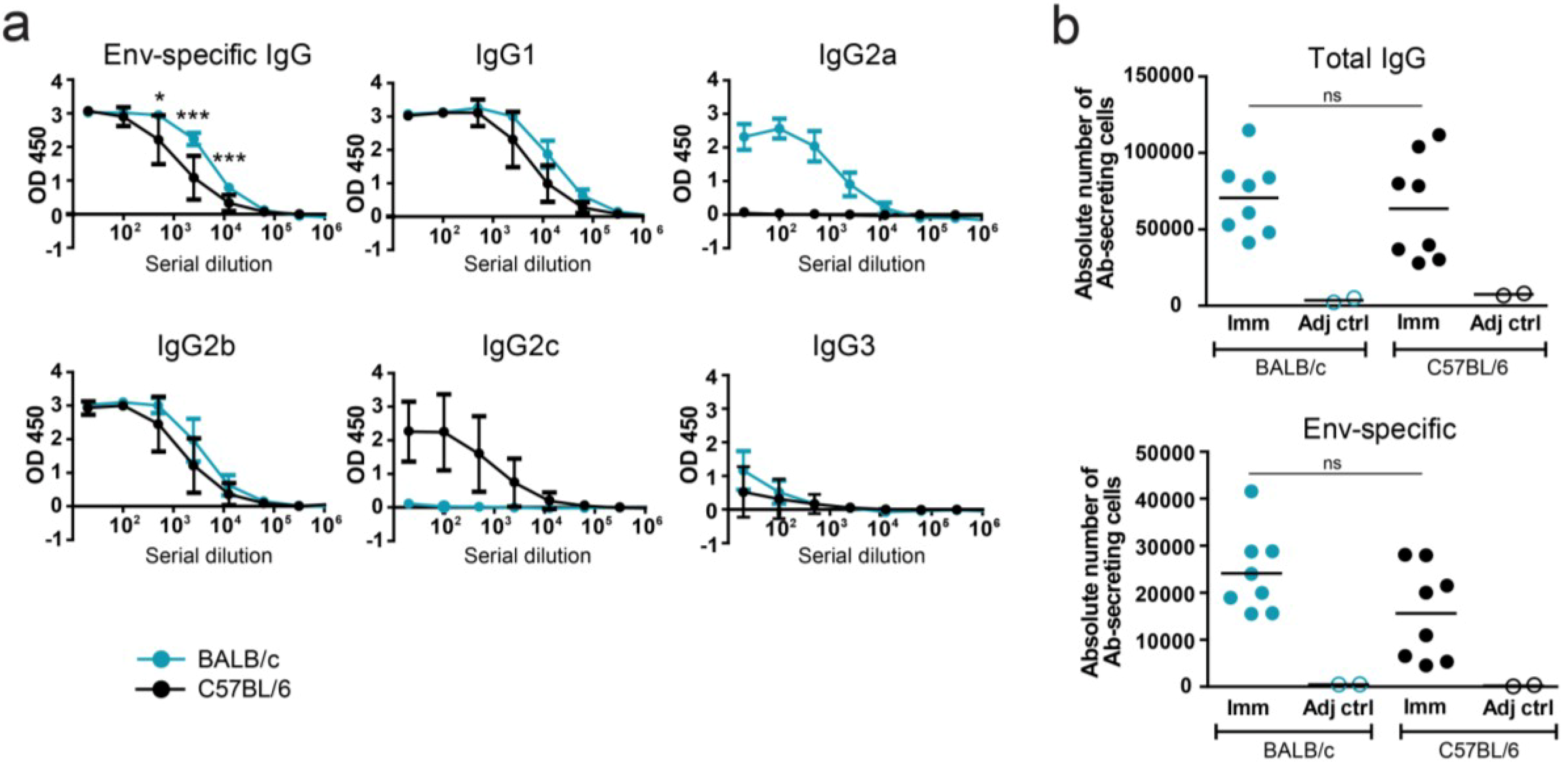{kind=link}
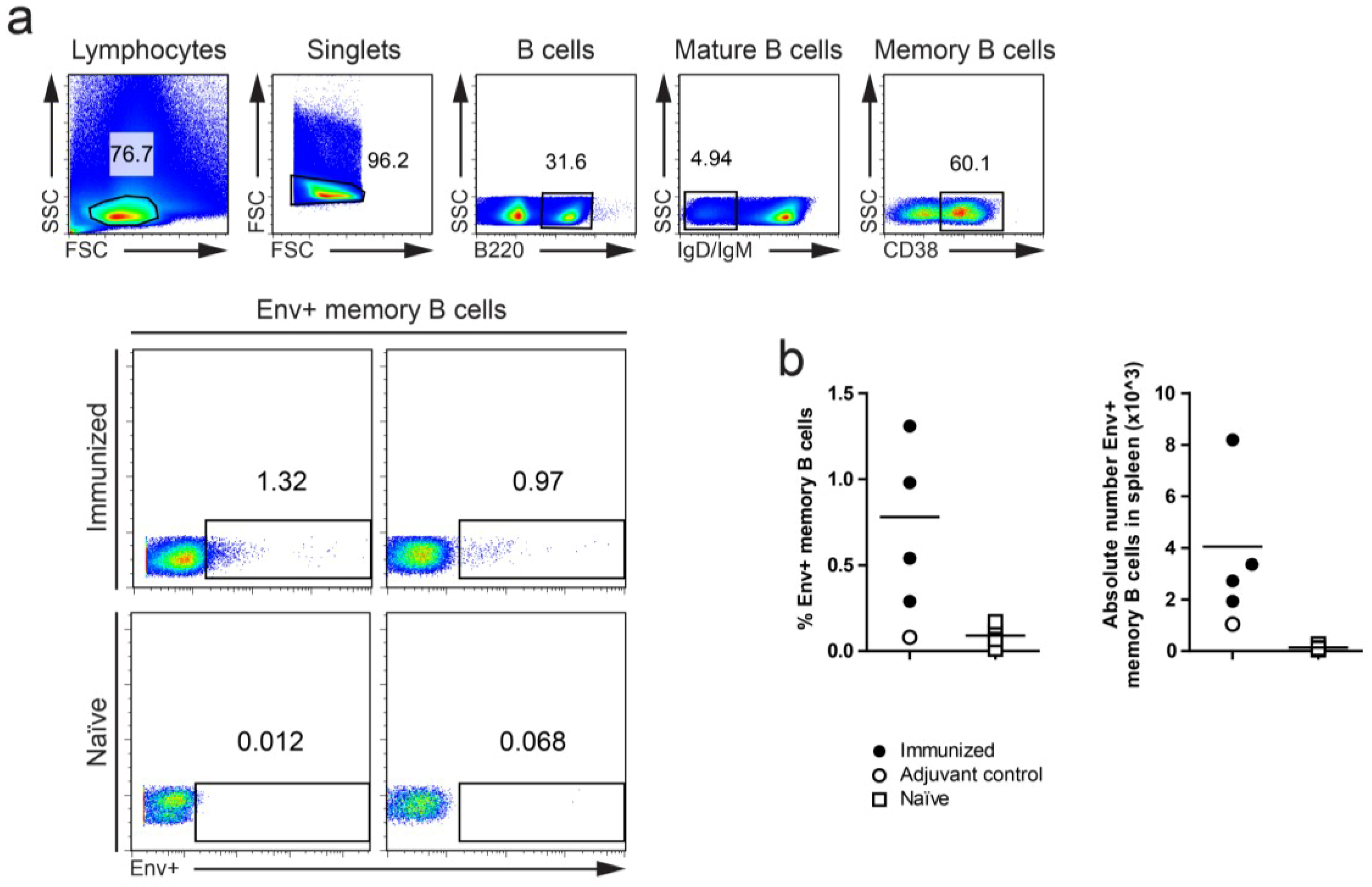{kind=link}
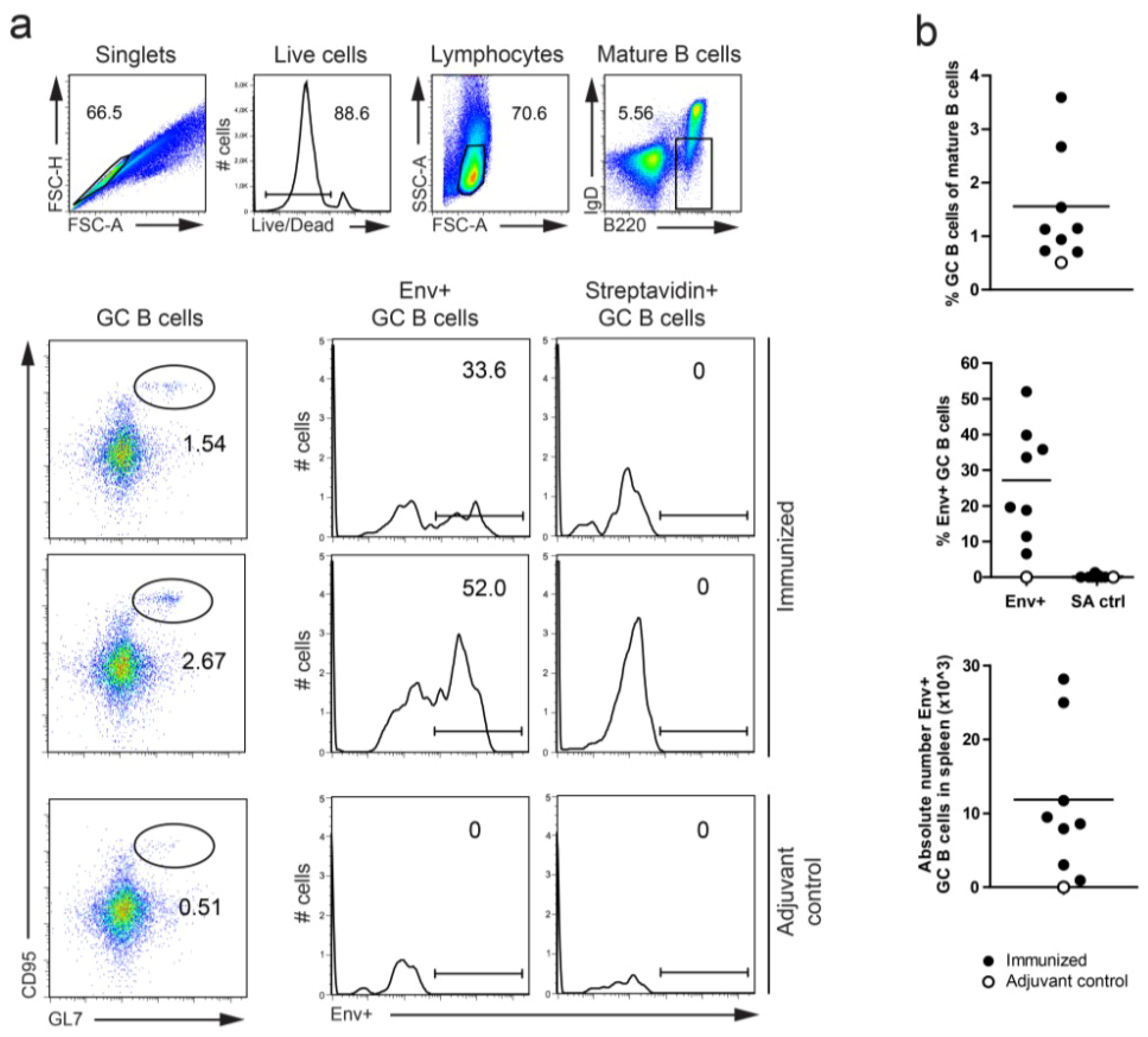{kind=link}
| Group | #ID | HXBc2 | MN | SF162 | BaL | 6535 | Neg Ctrl SIVmac |
|---|---|---|---|---|---|---|---|
| 1 | 97 | 15 | 20 | 49 | 14 | 18 | |
| 2 | 1151 | 768 | 47 | 10 | 13 | 10 | |
| Immunized | 3 | <5 | 58 | 38 | 30 | 5 | <5 |
| 4 | 17 | 24 | 21 | 28 | 19 | 7 | |
| 5 | 71 | 172 | 56 | 49 | 12 | 8 | |
| 6 | 27 | 32 | 16 | 30 | 5 | 9 | |
| Adjuvant | 1 | <5 | <5 | <5 | 8 | <5 | <5 |
| control | 2 | <5 | <5 | 5 | 13 | 5 | 16 |
3.2. Definition of Env-Specific Memory B Cells and GC B Cells in C57BL/6 Mice


4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Zhang, L.; Kovalev, G.I.; Su, L. HIV-1 infection and pathogenesis in a novel humanized mouse model. Blood 2007, 109, 2978–2981. [Google Scholar]
- Sun, Z.; Denton, P.W.; Estes, J.D.; Othieno, F.A.; Wei, B.L.; Wege, A.K.; Melkus, M.W.; Padgett-Thomas, A.; Zupancic, M.; Haase, A.T.; et al. Intrarectal transmission, systemic infection, and CD4+ T cell depletion in humanized mice infected with HIV-1. J. Exp. Med. 2007, 204, 705–714. [Google Scholar]
- Baenziger, S.; Tussiwand, R.; Schlaepfer, E.; Mazzucchelli, L.; Heikenwalder, M.; Kurrer, M.O.; Behnke, S.; Frey, J.; Oxenius, A.; Joller, H.; et al. Disseminated and sustained HIV infection in CD34+ cord blood cell-transplanted Rag2-/-gamma c-/- mice. Proc. Natl. Acad. Sci. USA 2006, 103, 15951–15956. [Google Scholar]
- Akkina, R.; Berges, B.K.; Palmer, B.E.; Remling, L.; Neff, C.P.; Kuruvilla, J.; Connick, E.; Folkvord, J.; Gagliardi, K.; Kassu, A.; et al. Humanized Rag1-/- gammac-/- mice support multilineage hematopoiesis and are susceptible to HIV-1 infection via systemic and vaginal routes. PLoS One 2011, 6, e20169. [Google Scholar] [CrossRef]
- Legrand, N.; Ploss, A.; Balling, R.; Becker, P.D.; Borsotti, C.; Brezillon, N.; Debarry, J.; de Jong, Y.; Deng, H.; Di Santo, J.P.; et al. Humanized mice for modeling human infectious disease: challenges, progress, and outlook. Cell Host Microbe 2009, 6, 5–9. [Google Scholar]
- Li, M.; Gao, F.; Mascola, J.R.; Stamatatos, L.; Polonis, V.R.; Koutsoukos, M.; Voss, G.; Goepfert, P.; Gilbert, P.; Greene, K.M.; et al. Human immunodeficiency virus type 1 env clones from acute and early subtype B infections for standardized assessments of vaccine-elicited neutralizing antibodies. J. Virol. 2005, 79, 10108–10125. [Google Scholar] [CrossRef]
- Sundling, C.; Forsell, M.N.; O'Dell, S.; Feng, Y.; Chakrabarti, B.; Rao, S.S.; Lore, K.; Mascola, J.R.; Wyatt, R.T.; Douagi, I.; et al. Soluble HIV-1 Env trimers in adjuvant elicit potent and diverse functional B cell responses in primates. J. Exp. Med. 2010, 207, 2003–2017. [Google Scholar] [CrossRef]
- Sundling, C.; Li, Y.; Huynh, N.; Poulsen, C.; Wilson, R.; O'Dell, S.; Feng, Y.; Mascola, J.R.; Wyatt, R.T.; Karlsson Hedestam, G.B. High-resolution definition of vaccine-elicited B cell responses against the HIV primary receptor binding site. Sci. Transl. Med. 2012, 4, 142ra96. [Google Scholar]
- Sundling, C.; Martinez, P.; Soldemo, M.; Spangberg, M.; Bengtsson, K.L.; Stertman, L.; Forsell, M.N.; Karlsson Hedestam, G.B. Immunization of macaques with soluble HIV type 1 and influenza virus envelope glycoproteins results in a similarly rapid contraction of peripheral B-cell responses after boosting. J. Infect. Dis. 2013, 207, 426–431. [Google Scholar] [CrossRef]
- Sundling, C.; Phad, G.; Douagi, I.; Navis, M.; Karlsson Hedestam, G.B. Isolation of antibody V(D)J sequences from single cell sorted rhesus macaque B cells. J. Immunol. Meth. 2012, 386, 85–93. [Google Scholar] [CrossRef]
- Sundling, C.; Zhang, Z.; Phad, G.E.; Sheng, Z.; Wang, Y.; Mascola, J.R.; Li, Y.; Wyatt, R.T.; Shapiro, L.; Karlsson Hedestam, G.B. Single-Cell and Deep Sequencing of IgG-Switched Macaque B Cells Reveal a Diverse Ig Repertoire following Immunization. J. Immunol. 2014, 192, 3637–3644. [Google Scholar] [CrossRef]
- Lee, S.K.; Rigby, R.J.; Zotos, D.; Tsai, L.M.; Kawamoto, S.; Marshall, J.L.; Ramiscal, R.R.; Chan, T.D.; Gatto, D.; Brink, R.; et al. B cell priming for extrafollicular antibody responses requires Bcl-6 expression by T cells. J. Exp. Med. 2011, 208, 1377–1388. [Google Scholar]
- Paus, D.; Phan, T.G.; Chan, T.D.; Gardam, S.; Basten, A.; Brink, R. Antigen recognition strength regulates the choice between extrafollicular plasma cell and germinal center B cell differentiation. J. Exp. Med. 2006, 203, 1081–1091. [Google Scholar] [CrossRef]
- Adams, C.L.; Macleod, M.K.; James Milner-White, E.; Aitken, R.; Garside, P.; Stott, D.I. Complete analysis of the B-cell response to a protein antigen, from in vivo germinal centre formation to 3-D modelling of affinity maturation. Immunology 2003, 108, 274–287. [Google Scholar] [CrossRef]
- Zhang, Y.; Meyer-Hermann, M.; George, L.A.; Figge, M.T.; Khan, M.; Goodall, M.; Young, S.P.; Reynolds, A.; Falciani, F.; Waisman, A.; et al. Germinal center B cells govern their own fate via antibody feedback. J. Exp. Med. 2013, 210, 457–464. [Google Scholar]
- Lu, Y.F.; Singh, M.; Cerny, J. Canonical germinal center B cells may not dominate the memory response to antigenic challenge. Int. Immunol. 2001, 13, 643–655. [Google Scholar] [CrossRef]
- Doyle-Cooper, C.; Hudson, K.E.; Cooper, A.B.; Ota, T.; Skog, P.; Dawson, P.E.; Zwick, M.B.; Schief, W.R.; Burton, D.R.; Nemazee, D. Immune tolerance negatively regulates B cells in knock-in mice expressing broadly neutralizing HIV antibody 4E10. J. Immunol. 2013, 191, 3186–3191. [Google Scholar] [CrossRef]
- Verkoczy, L.; Chen, Y.; Zhang, J.; Bouton-Verville, H.; Newman, A.; Lockwood, B.; Scearce, R.M.; Montefiori, D.C.; Dennison, S.M.; Xia, S.M.; et al. Induction of HIV-1 broad neutralizing antibodies in 2F5 knock-in mice: selection against membrane proximal external region-associated autoreactivity limits T-dependent responses. J. Immunol. 2013, 191, 2538–2550. [Google Scholar] [CrossRef]
- Klein, F.; Diskin, R.; Scheid, J.F.; Gaebler, C.; Mouquet, H.; Georgiev, I.S.; Pancera, M.; Zhou, T.; Incesu, R.B.; Fu, B.Z.; et al. Somatic mutations of the immunoglobulin framework are generally required for broad and potent HIV-1 neutralization. Cell 2013, 153, 126–138. [Google Scholar] [CrossRef]
- Dosenovic, P.; Chakrabarti, B.; Soldemo, M.; Douagi, I.; Forsell, M.N.; Li, Y.; Phogat, A.; Paulie, S.; Hoxie, J.; Wyatt, R.T.; et al. Selective expansion of HIV-1 envelope glycoprotein-specific B cell subsets recognizing distinct structural elements following immunization. J. Immunol. 2009, 183, 3373–3382. [Google Scholar]
- Forsell, M.N.; Soldemo, M.; Dosenovic, P.; Wyatt, R.T.; Karlsson, M.C.; Karlsson Hedestam, G.B. Independent expansion of epitope-specific plasma cell responses upon HIV-1 envelope glycoprotein immunization. J. Immunol. 2013, 191, 44–51. [Google Scholar] [CrossRef]
- Yang, X.; Lee, J.; Mahony, E.M.; Kwong, P.D.; Wyatt, R.; Sodroski, J. Highly stable trimers formed by human immunodeficiency virus type 1 envelope glycoproteins fused with the trimeric motif of T4 bacteriophage fibritin. J. Virol. 2002, 76, 4634–4642. [Google Scholar]
- Flowjo, version vX.0.6; Tree Star, Inc.: Ashland, OR, USA, 2013.
- Wu, L.; Oficjalska, K.; Lambert, M.; Fennell, B.J.; Darmanin-Sheehan, A.; Ni Shuilleabhain, D.; Autin, B.; Cummins, E.; Tchistiakova, L.; Bloom, L.; et al. Fundamental characteristics of the immunoglobulin VH repertoire of chickens in comparison with those of humans, mice, and camelids. J. Immunol. 2012, 188, 322–333. [Google Scholar] [CrossRef]
- Lee, E.C.; Liang, Q.; Ali, H.; Bayliss, L.; Beasley, A.; Bloomfield-Gerdes, T.; Bonoli, L.; Brown, R.; Campbell, J.; Carpenter, A.; et al. Complete humanization of the mouse immunoglobulin loci enables efficient therapeutic antibody discovery. Nat. Biotechnol. 2014, 32, 356–363. [Google Scholar]
- Dosenovic, P.; Soldemo, M.; Scholz, J.L.; O'Dell, S.; Grasset, E.K.; Pelletier, N.; Karlsson, M.C.; Mascola, J.R.; Wyatt, R.T.; Cancro, M.P.; et al. BLyS-mediated modulation of naive B cell subsets impacts HIV Env-induced antibody responses. J. Immunol. 2012, 188, 6018–6026. [Google Scholar] [CrossRef]
- Forsell, M.N.; Li, Y.; Sundback, M.; Svehla, K.; Liljestrom, P.; Mascola, J.R.; Wyatt, R.; Hedestam, G.B. Biochemical and immunogenic characterization of soluble human immunodeficiency virus type 1 envelope glycoprotein trimers expressed by semliki forest virus. J. Virol. 2005, 79, 10902–10914. [Google Scholar] [CrossRef]
- Martin, R.M.; Brady, J.L.; Lew, A.M. The need for IgG2c specific antiserum when isotyping antibodies from C57BL/6 and NOD mice. J. Immunol. Meth. 1998, 212, 187–192. [Google Scholar]
- Jouvin-Marche, E.; Morgado, M.G.; Leguern, C.; Voegtle, D.; Bonhomme, F.; Cazenave, P.A. The mouse Igh-1a and Igh-1b H chain constant regions are derived from two distinct isotypic genes. Immunogenetics 1989, 29, 92–97. [Google Scholar]
- Morgado, M.G.; Cam, P.; Gris-Liebe, C.; Cazenave, P.A.; Jouvin-Marche, E. Further evidence that BALB/c and C57BL/6 gamma 2a genes originate from two distinct isotypes. EMBO J. 1989, 8, 3245–3251. [Google Scholar]
- Tiller, T.; Busse, C.E.; Wardemann, H. Cloning and expression of murine Ig genes from single B cells. J. Immunol. Meth. 2009, 350, 183–193. [Google Scholar]
- Plotkin, S.A. Complex correlates of protection after vaccination. Clin. Infect. Dis. 2013, 56, 1458–1465. [Google Scholar]
- Tran, K.; Poulsen, C.; Guenaga, J.; de Val, N.; Wilson, R.; Sundling, C.; Li, Y.; Stanfield, R.L.; Wilson, I.A.; Ward, A.B.; et al. Vaccine-elicited primate antibodies use a distinct approach to the HIV-1 primary receptor binding site informing vaccine redesign. Proc. Natl. Acad. Sci. USA 2014, 111, E738–E747. [Google Scholar] [CrossRef]
- Wolniak, K.L.; Noelle, R.J.; Waldschmidt, T.J. Characterization of (4-hydroxy-3-nitrophenyl)acetyl (NP)-specific germinal center B cells and antigen-binding B220- cells after primary NP challenge in mice. J. Immunol. 2006, 177, 2072–2079. [Google Scholar] [CrossRef]
- Rankin, A.L.; MacLeod, H.; Keegan, S.; Andreyeva, T.; Lowe, L.; Bloom, L.; Collins, M.; Nickerson-Nutter, C.; Young, D.; Guay, H. IL-21 receptor is critical for the development of memory B cell responses. J. Immunol. 2011, 186, 667–674. [Google Scholar] [CrossRef]
- McHeyzer-Williams, L.J.; Cool, M.; McHeyzer-Williams, M.G. Antigen-specific B cell memory: Expression and replenishment of a novel b220(-) memory b cell compartment. J. Exp. Med. 2000, 191, 1149–1166. [Google Scholar] [CrossRef]
- Liu, H.; Moynihan, K.D.; Zheng, Y.; Szeto, G.L.; Li, A.V.; Huang, B.; Van Egeren, D.S.; Park, C.; Irvine, D.J. Structure-based programming of lymph-node targeting in molecular vaccines. Nature 2014, 507, 519–522. [Google Scholar]
- Kanekiyo, M.; Wei, C.J.; Yassine, H.M.; McTamney, P.M.; Boyington, J.C.; Whittle, J.R.; Rao, S.S.; Kong, W.P.; Wang, L.; Nabel, G.J. Self-assembling influenza nanoparticle vaccines elicit broadly neutralizing H1N1 antibodies. Nature 2013, 499, 102–106. [Google Scholar]
- Nie, X.; Basu, S.; Cerny, J. Immunization with immune complex alters the repertoire of antigen-reactive B cells in the germinal centers. Eur J. Immunol. 1997, 27, 3517–3525. [Google Scholar] [CrossRef]
- Isik, G.; van Montfort, T.; Boot, M.; Cobos Jimenez, V.; Kootstra, N.A.; Sanders, R.W. Chimeric HIV-1 envelope glycoproteins with potent intrinsic granulocyte-macrophage colony-stimulating factor (GM-CSF) activity. PLoS One 2013, 8, e60126. [Google Scholar]
- Phogat, S.; Svehla, K.; Tang, M.; Spadaccini, A.; Muller, J.; Mascola, J.; Berkower, I.; Wyatt, R. Analysis of the human immunodeficiency virus type 1 gp41 membrane proximal external region arrayed on hepatitis B surface antigen particles. Virology 2008, 373, 72–84. [Google Scholar] [CrossRef]
- Koch, M.; Frazier, J.; Sodroski, J.; Wyatt, R. Characterization of antibody responses to purified HIV-1 gp120 glycoproteins fused with the molecular adjuvant C3d. Virology 2005, 340, 277–284. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Soldemo, M.; Pedersen, G.K.; Karlsson Hedestam, G.B. HIV-1 Env-Specific Memory and Germinal Center B Cells in C57BL/6 Mice. Viruses 2014, 6, 3400-3414. https://doi.org/10.3390/v6093400
Soldemo M, Pedersen GK, Karlsson Hedestam GB. HIV-1 Env-Specific Memory and Germinal Center B Cells in C57BL/6 Mice. Viruses. 2014; 6(9):3400-3414. https://doi.org/10.3390/v6093400
Chicago/Turabian StyleSoldemo, Martina, Gabriel K. Pedersen, and Gunilla B. Karlsson Hedestam. 2014. "HIV-1 Env-Specific Memory and Germinal Center B Cells in C57BL/6 Mice" Viruses 6, no. 9: 3400-3414. https://doi.org/10.3390/v6093400
APA StyleSoldemo, M., Pedersen, G. K., & Karlsson Hedestam, G. B. (2014). HIV-1 Env-Specific Memory and Germinal Center B Cells in C57BL/6 Mice. Viruses, 6(9), 3400-3414. https://doi.org/10.3390/v6093400