
Is the New Variant RHDV Replacing Genogroup 1 in Portuguese Wild Rabbit Populations?


 ,
, 

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
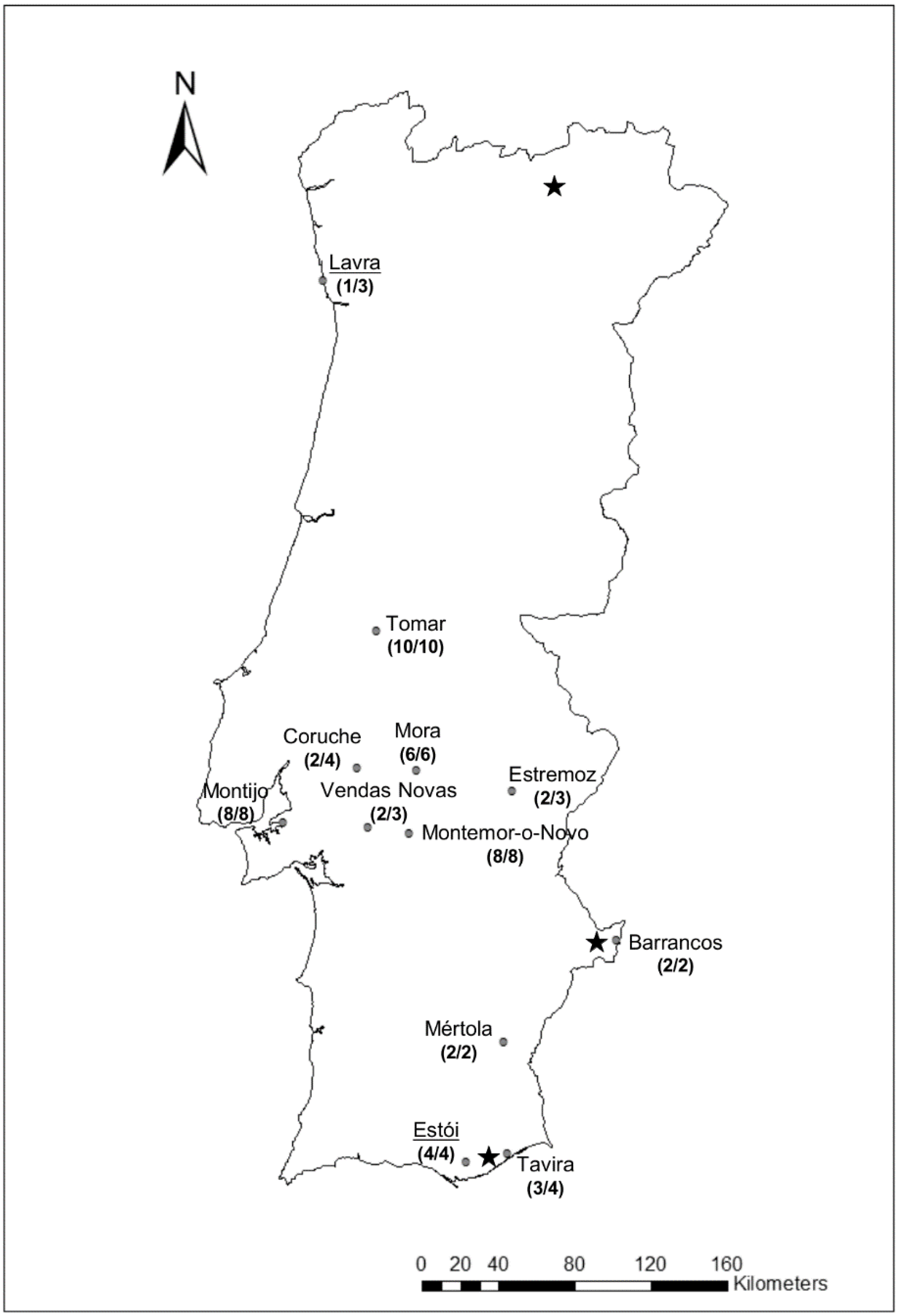
2. Materials and Methods

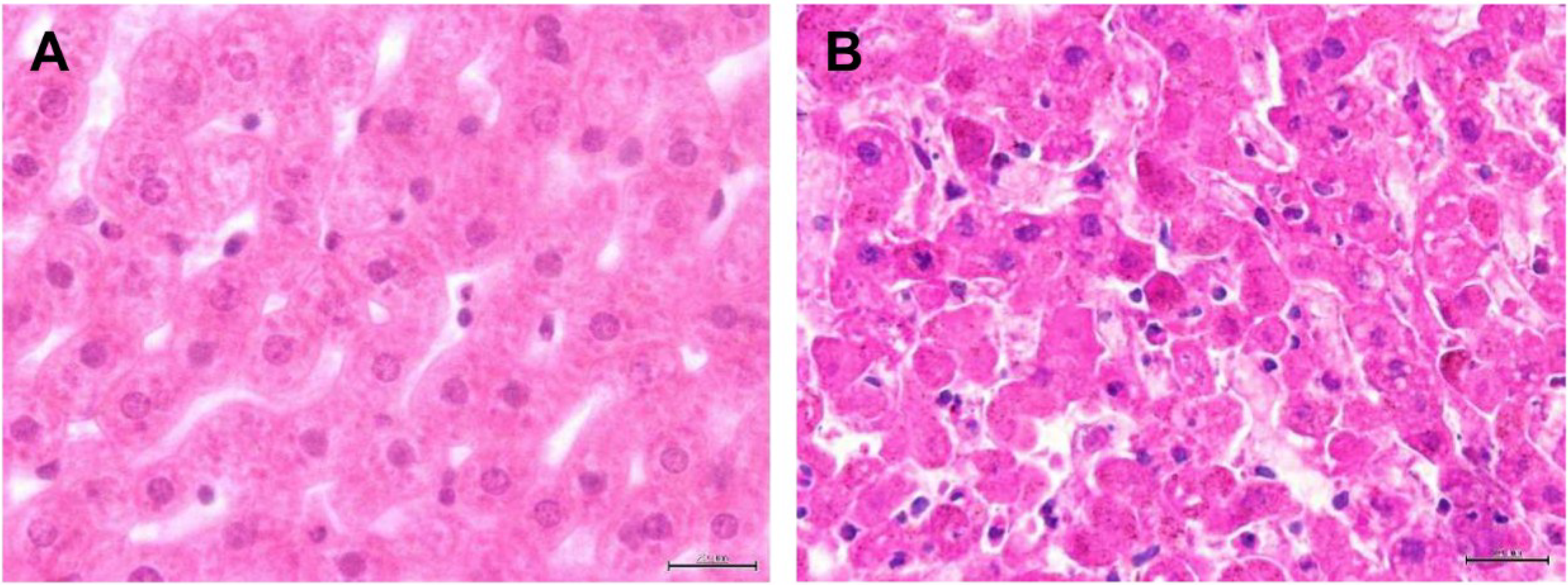
3. Results and Discussion


Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Liu, S.; Xue, H.; Pu, B.; Qian, N. A new viral disease in rabbit. Anim. Husb. Vet. Med. 1984, 16, 253–255. [Google Scholar]
- Marcato, P.S.; Benazzi, C.; Vecchi, G.; Galeotti, M.; Salda, L.D.; Sarli, G.; Lucidi, P. Clinical and pathological features of viral haemorrhagic disease of rabbits and the European brown hare syndrome. Rev. Sci. Tech. 1991, 10, 371–392. [Google Scholar] [PubMed]
- Abrantes, J.; van der Loo, W.; Le Pendu, J.; Esteves, P.J. Rabbit haemorrhagic disease (RHD) and rabbit haemorhhagic disease virus (RHDV): A review. Vet. Res. 2012, 43, 12. [Google Scholar] [CrossRef] [PubMed]
- Cancellotti, F.M.; Renzi, M. Epidemiology and current situation of viral haemorrhagic disease of rabbits and the European brown hare syndrome in Italy. Rev. Sci. Tech. Off. Int. Epiz. 1991, 10, 409–422. [Google Scholar]
- Mutze, G.; Kovaliski, J.; Butler, K.; Capucci, L.; McPhee, S. The effect of rabbit population control programmes on the impact of rabbit haemorrhagic disease in south-eastern Australia. J. Appl. Ecol. 2010, 47, 1137–1146. [Google Scholar] [CrossRef]
- Le Gall, G.; Arnauld, C.; Boilletot, E.; Morisse, J.P.; Rasschaert, D. Molecular epidemiology of rabbit haemorrhagic disease virus outbreaks in France during 1988 to 1995. J. Gen. Virol. 1998, 79, 11–16. [Google Scholar] [PubMed]
- Le Gall-Reculé, G.; Zwingelstein, F.; Laurent, S.; de Boisséson, C.; Portejoie, Y.; Rasschaert, D. Phylogenetic analysis of rabbit haemorrhagic disease virus in France between 1993 and 2000, and the characterisation of RHDV antigenic variants. Arch. Virol. 2003, 148, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; Kitchen, A.; Holmes, E.C. Origin and phylodynamics of rabbit hemorrhagic disease virus. J. Virol. 2009, 83, 12129–12138. [Google Scholar] [CrossRef] [PubMed]
- Kinnear, M.; Linde, C.C. Capsid gene divergence in rabbit hemorrhagic disease virus. J. Gen. Virol. 2010, 91, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Capucci, L.; Fusi, P.; Lavazza, A.; Pacciarini, M.L.; Rossi, C. Detection and preliminary characterization of a new rabbit calicivirus related to rabbit hemorrhagic disease virus but nonpathogenic. J. Virol. 1996, 70, 8614–8623. [Google Scholar] [PubMed]
- Strive, T.; Wright, J.D.; Robinson, A.J. Identification and partial characterisation of a new lagovirus in Australian wild rabbits. Virology 2009, 384, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Bergin, I.L.; Wise, A.G.; Bolin, S.R.; Mullaney, T.P.; Kiupel, M.; Maes, R.K. Novel calicivirus identified in rabbits, Michigan, USA. Emerg. Infect. Dis. 2009, 15, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Abrantes, J.; Esteves, P.J. Not-so-novel Michigan rabbit calicivirus. Emerg. Infect. Dis. 2010, 16, 1331–1332. [Google Scholar] [CrossRef] [PubMed]
- Le Gall-Reculé, G.; Zwingelstein, F.; Fages, M.-P.; Bertagnoli, S.; Gelfi, J.; Aubineau, J.; Roobrouck, A.; Botti, G.; Lavazza, A.; Marchandeau, S. Characterisation of a non-pathogenic and non-protective infectious rabbit lagovirus related to RHDV. Virology 2011, 410, 395–402. [Google Scholar] [Green Version]
- Le Gall, G.; Huguet, S.; Vende, P.; Vautherot, J.-F.; Rasschaert, D. European brown hare syndrome virus: molecular cloning and sequencing of the genome. J. Gen. Virol. 1996, 77, 1693–1697. [Google Scholar]
- Le Gall-Reculé, G.; Zwingelstein, F.; Boucher, S.; Le Normand, B.; Plassiart, G.; Portejoie, Y.; Decors, A.; Bertagnoli, S.; Guérin, J.L.; Marchandeau, S. Detection of a new variant of rabbit haemorrhagic disease virus in France. Vet. Rec. 2011, 168, 137–138. [Google Scholar]
- Dalton, K.P.; Nicieza, I.; Balseiro, A.; Muguerza, M.A.; Rosell, J.M.; Casais, R.; Álvarez, Á.L.; Parra, F. Variant rabbit hemorrhagic disease virus in young rabbits, Spain. Emerg. Infect. Dis. 2012, 18, 2009–2012. [Google Scholar] [CrossRef]
- Abrantes, J.; Lopes, A.M.; Dalton, K.P.; Melo, P.; Correia, J.J.; Ramada, M.; Alves, P.C.; Parra, F.; Esteves, P.J. New variant of rabbit hemorrhagic disease virus, Portugal, 2012–2013. Emerg. Infect. Dis. 2013, 19, 1900–1902. [Google Scholar] [CrossRef] [PubMed]
- Le Gall-Reculé, G.; Lavazza, A.; Marchandeau, S.; Bertagnoli, S.; Zwingelstein, F.; Cavadini, P.; Martinelli, N.; Lombardi, G.; Guerin, J.-L.; Lemaitre, E.; et al. Emergence of a new lagovirus related to Rabbit Haemorrhagic Disease Virus. Vet. Res. 2013, 44, 81. [Google Scholar]
- Westcott, D.; Frossard, J.-P.; Everest, D.; Dastjerdi, A.; Duff, J.P.; Steinbach, F.; Choudhury, B. Incursion of RHDV2-like variant in Great Britain. Vet. Rec. 2014, 174, 333. [Google Scholar] [CrossRef] [PubMed]
- Baily, J.L.; Dagleish, M.P.; Graham, M.; Maley, M.; Rocchi, M.S. RHDV variant 2 presence detected in Scotland. Vet. Rec. 2014, 174, 411. [Google Scholar] [CrossRef] [PubMed]
- Müller, A.; Freitas, J.; Silva, E.; Le Gall-Reculé, G.; Zwingelstein, F.; Abrantes, J.; Esteves, P.J.; Alves, P.C.; van der Loo, W.; Kolodziejek, J.; et al. Evolution of rabbit haemorrhagic disease virus (RHDV) in the European rabbit (Oryctolagus cuniculus) from the Iberian Peninsula. Vet. Microbiol. 2009, 135, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Alda, F.; Gaitero, T.; Suárez, M.; Merchán, T.; Rocha, G.; Doadrio, I. Evolutionary history and molecular epidemiology of rabbit haemorrhagic disease virus in the Iberian Peninsula and Western Europe. BMC Evol. Biol. 2010, 10, 347–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrantes, J.; Lopes, A.M.; Esteves, P.J. Complete genomic sequences of rabbit hemorrhagic disease virus G1 strains isolated in the European rabbit original range. J. Virol. 2012, 86, 13886. [Google Scholar] [CrossRef] [PubMed]
- Abrantes, J.; Lopes, A.M.; Dalton, K.P.; Parra, F.; Esteves, P.J. Detection of RHDVa on the Iberian Peninsula: Isolation of an RHDVa strain from a Spanish rabbitry. Arch. Virol. 2014, 159, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Ferrand, N.; Branco, M. The evolutionary history of the European rabbit (Oryctolagus cuniculus): Major patterns of population differentiation and geographic expansion inferred from protein polymorphism. In Phylogeography of Southern European Refugia; Weiss, S., Ferrand, N., Eds.; Springer: Dordrecht, the Netherlands, 2007; p. 207. [Google Scholar]
- Dalton, K.P.; Abrantes, J.; Lopes, A.M.; Nicieza, I.; Álvarez, Á.L.; Esteves, P.J.; Parra, F. Complete genome sequence of two variant RHDVb isolates detected on the Iberian Peninsula. Arch. Virol. accepted.
- Lopes, A.M.; Marques, S.; Silva, E.; Magalhães, M.J.; Pinheiro, A.; Alves, P.C.; Le Pendu, J.; Esteves, P.J.; Thompson, G.; Abrantes, J. Detection of RHDV strains in the Iberian hare (Lepus granatensis): Earliest evidence of rabbit lagovirus cross-species infection. Vet. Res. 2014, 45, 94. [Google Scholar] [PubMed]
- Esteves, P.J.; Lopes, A.M.; Magalhães, M.J.; Pinheiro, A.; Gonçalves, D.; Abrantes, J. Rabbit hemorrhagic disease virus detected in Pico, Azores, Portugal, revealed a unique endemic strain with more than 17 years of independent evolution. Viruses 2014, 6, 2698–2707. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Delibes-Mateos, M.; Ferreira, C.; Carro, F.; Escudero, M.A.; Gortázar, C. Ecosystem effects of variant rabbit hemorrhagic disease virus, Iberian Peninsula. Emerg. Infect. Dis. 2014, 20, 2166–2168. [Google Scholar] [CrossRef] [PubMed]
- Calvete, C.; Sarto, P.; Calvo, A.J.; Monroy, F.; Calvo, J.H. Could the new rabbit haemorrhagic disease virus variant (RHDVb) be fully replacing classical RHD strains in the Iberian Peninsula? World Rabbit Sci. 2014, 22, 91. [Google Scholar] [CrossRef]
- Dalton, K.P.; Nicieza, I.; Abrantes, J.; Esteves, P.J.; Parra, F. Spread of new variant RHDV in domestic rabbits on the Iberian Peninsula. Vet. Microbiol. 2014, 169, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Elsworth, P.; Cooke, B.D.; Kovaliski, J.; Sinclair, R.; Holmes, E.C.; Strive, T. Increased virulence of rabbit haemorrhagic disease virus associated with genetic resistance in wild Australian rabbits (Oryctolagus cuniculus). Virology 2014, 464–465, 415–423. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopes, A.M.; Correia, J.; Abrantes, J.; Melo, P.; Ramada, M.; Magalhães, M.J.; Alves, P.C.; Esteves, P.J. Is the New Variant RHDV Replacing Genogroup 1 in Portuguese Wild Rabbit Populations? Viruses 2015, 7, 27-36. https://doi.org/10.3390/v7010027
Lopes AM, Correia J, Abrantes J, Melo P, Ramada M, Magalhães MJ, Alves PC, Esteves PJ. Is the New Variant RHDV Replacing Genogroup 1 in Portuguese Wild Rabbit Populations? Viruses. 2015; 7(1):27-36. https://doi.org/10.3390/v7010027
Chicago/Turabian StyleLopes, Ana M., Jorge Correia, Joana Abrantes, Pedro Melo, Margarida Ramada, Maria J. Magalhães, Paulo C. Alves, and Pedro J. Esteves. 2015. "Is the New Variant RHDV Replacing Genogroup 1 in Portuguese Wild Rabbit Populations?" Viruses 7, no. 1: 27-36. https://doi.org/10.3390/v7010027
APA StyleLopes, A. M., Correia, J., Abrantes, J., Melo, P., Ramada, M., Magalhães, M. J., Alves, P. C., & Esteves, P. J. (2015). Is the New Variant RHDV Replacing Genogroup 1 in Portuguese Wild Rabbit Populations? Viruses, 7(1), 27-36. https://doi.org/10.3390/v7010027