
Ageratum enation virus—A Begomovirus of Weeds with the Potential to Infect Crops

Abstract
:1. Introduction
2. Materials and Methods
2.1. DNA Extraction, PCR Amplification and Cloning
2.2. Sequencing and Sequence Analysis
2.3. Constructs for Infectivity
2.4. Agrobacterium-Mediated Inoculation
3. Results
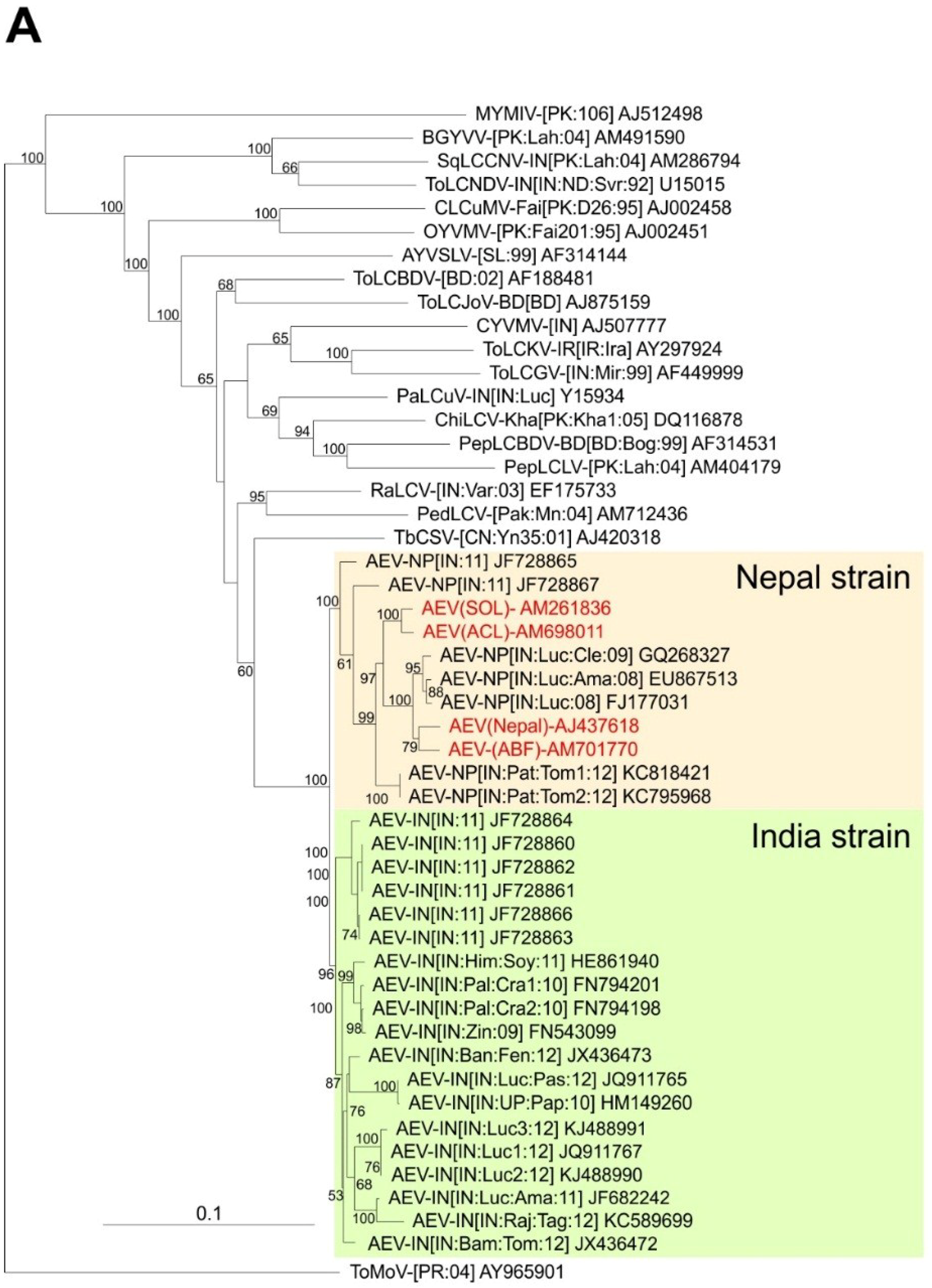
3.1. Cloning, Sequencing and Sequence Analysis of Begomoviruses


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Origin (year) | Plant Species (Symptoms *) | Component | Accession No. | Size (bp) | ORF | Start/Stop Codon (Nucleotide Coordinates) | Predicted Size (no. of Amino Acids) | Predicted Molecular Weight (kDa) |
|---|---|---|---|---|---|---|---|---|---|
| SOL | Lahore, Pakistan (2005) | Sonchus oleraceus (YV) | begomovirus | AM261836 | 2750 | V2 | 135/482 | 115 | 13.3 |
| CP | 295/1065 | 256 | 29.7 | ||||||
| Rep | 2599/1514 | 361 | 40.7 | ||||||
| TrAP | 1611/1180 | 143 | 16.2 | ||||||
| REn | 1466/1062 | 134 | 15.9 | ||||||
| C4 | 2442/2185 | 85 | 9.3 | ||||||
| betasatellite | AM412239 | 1368 | βC1 | 605/189 | 138 | 16.1 | |||
| ACL | Lahore, Pakistan (2006) | Ageratum conyzoides (YV) | begomovirus | AM698011 | 2749 | V2 | 134/481 | 115 | 13.3 |
| CP | 294/1064 | 256 | 29.6 | ||||||
| Rep | 2598/1513 | 361 | 40.8 | ||||||
| TrAP | 1610/1206 | 134 | 15.0 | ||||||
| REn | 1465/1061 | 134 | 15.9 | ||||||
| C4 | 2441/2184 | 85 | 9.4 | ||||||
| betasatellite | AM698010 | 1355 | βC1 | 596/180 | 138 | 16.1 | |||
| ACN | Nepal (2001) | Ageratum conyzoides (LC, EN) | begomovirus | AJ437618 | 2746 | V2 | 134/481 | 115 | 13.3 |
| CP | 294/1064 | 256 | 29.5 | ||||||
| Rep | 2597/1512 | 361 | 40.6 | ||||||
| TrAP | 1610/1206 | 134 | 15.1 | ||||||
| REn | 1465/1061 | 134 | 15.9 | ||||||
| C4 | 2441/2184 | 85 | 9.2 | ||||||
| betasatellite | - | - | - | - | - | - | |||
| ABF | Faisalabad, Pakistan (2006) | Brassica rapa var. rapa (YV) | begomovirus | AM701770 | 2747 | V2 | 134/481 | 115 | 13.3 |
| CP | 294/1064 | 256 | 29.6 | ||||||
| Rep | 2598/1513 | 361 | 40.8 | ||||||
| TrAP | 1610/1206 | 134 | 15.2 | ||||||
| REn | 1465/1061 | 134 | 15.8 | ||||||
| C4 | 2441/2184 | 85 | 9.4 | ||||||
| betasatellite | AM701771 | 1359 | βC1 | 596/180 | 138 | 16.1 |
3.2. Comparison of the Sequences Obtained Here with Sequences Available in the Databases


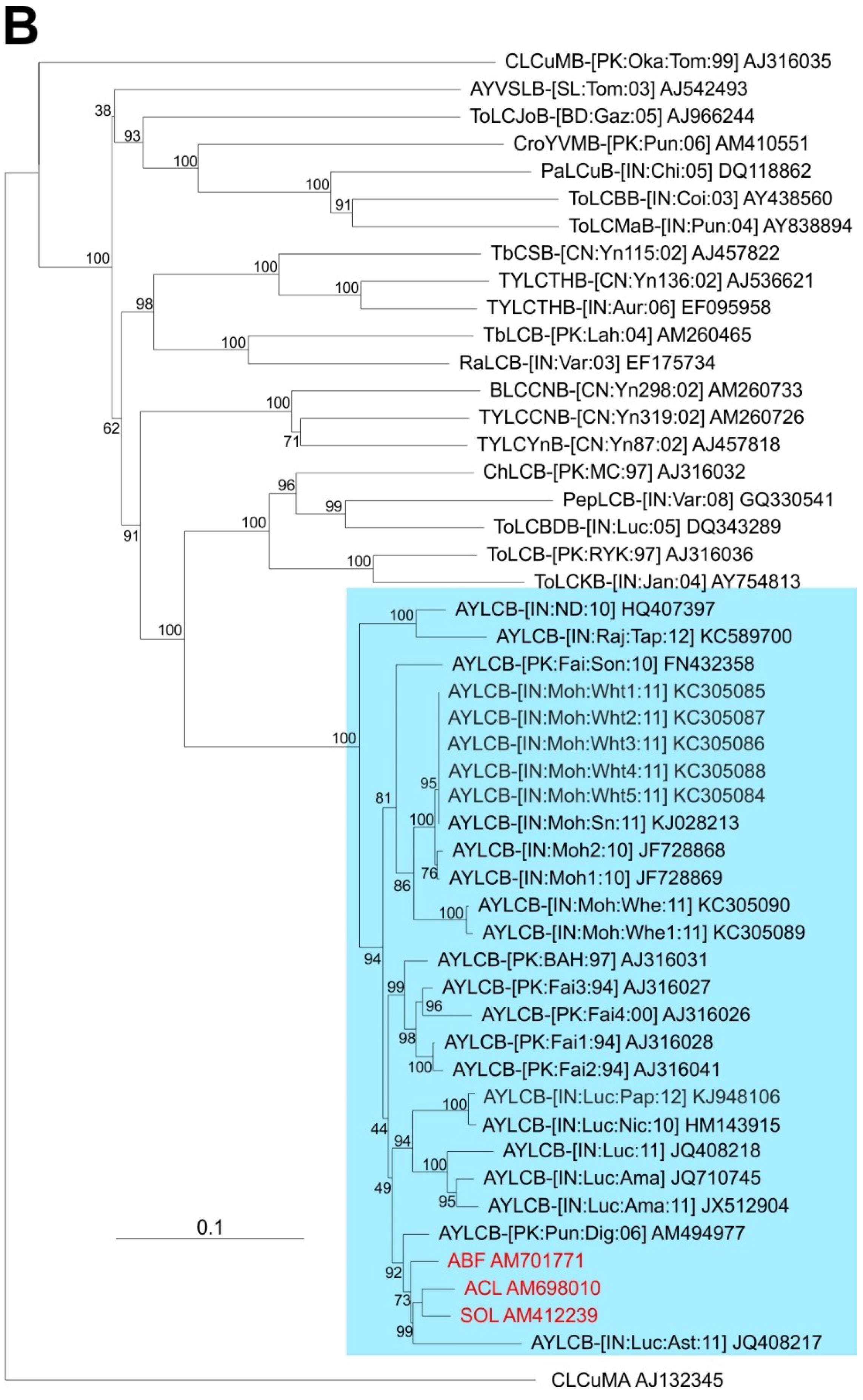
3.3. Cloning, Sequencing and Sequence Analysis of Betasatellites
3.4. Analysis of the Infectivity of AEV and Betasatellite Clones
| Inoculum | Plant Species | Infectivity (Plants Infected/Plants Inoculated) | Latent Period # (days) | PCR Detection | Symptoms * | |
|---|---|---|---|---|---|---|
| Virus | Betasatellite | |||||
| AEVSOL and AYLCBSOL | N. benthamiana | 60/60 | 12–18 | + | + | D, LC, C |
| N. tabacum | 20/20 | 12–18 | + | + | VY, D | |
| S. lycopersicon | 20/20 | 15–21 | + | + | C, N | |
| A. conyzoides | 2/2 | 15–18 | + | + | CR, LC | |
| S. oleraceus | 0/5 | - | - | - | - | |
| AEVSOL | N. benthamiana | 22/30 | 12–18 | + | - | - |
| AYLCBSOL | N. benthamiana | 00/30 | - | - | - | - |
| pGreen0029 | N. benthamiana | 00/20 | - | - | - | - |
| AEVACL and AYLCBACL | N. benthamiana | 10/10 | 12–18 | + | + | R, LC, C |
| S. lycopersicon | 10/10 | 15–21 | + | + | SG, C, R, N | |
| A. conyzoides | 2/2 | 15–18 | + | + | CR, LC | |
| S. oleraceus | 0/5 | - | - | - | - | |
| none | N. benthamiana | 00/20 | - | - | - | - |
| pGreen0029 | N. benthamiana | 00/20 | - | - | - | - |


4. Discussion
Supplementary Files
Supplementary File 1Acknowledgements
Author Contributions
Conflicts of Interest
References
- Lefeuvre, P.; Martin, D.P.; Harkins, G.; Lemey, P.; Gray, A.J.A.; Meredith, S.; Lakay, F.; Monjane, A.; Lett, J.M.; Varsani, A.; et al. The spread of Tomato yellow leaf curl virus from the Middle East to the world. PLoS Pathog. 2010, 6, e1001164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wren, J.D.; Roossinck, M.J.; Nelson, R.S.; Scheets, K.; Palmer, M.W.; Melcher, U. Plant virus biodiversity and ecology. PLoS Biol. 2006, 4, e80. [Google Scholar] [CrossRef] [PubMed]
- Mubin, M.; Akhtar, S.; Amin, I.; Briddon, R.W.; Mansoor, S. Xanthiumstrumarium: A weed host of components of begomovirus-betasatellite complexes affecting crops. Virus Genes 2012, 44, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Ilyas, M.; Qazi, J.; Mansoor, S.; Briddon, R.W. Molecular characterisation and infectivity of a “Legumovirus” (genus Begomovirus: Family Geminiviridae) infecting the leguminous weed Rhynchosia minima in Pakistan. Virus Res. 2009, 145, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.K.; Fauquet, C.M.; Briddon, R.W.; Zerbini, M.; Moriones, E.; Navas-Castillo, J. Virus Taxonomy—Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Lefkowitz, E., Adams, M.J., Carstens, E.B., Eds.; Associated Press: London, UK, 2012; pp. 351–373. [Google Scholar]
- Varsani, A.; Navas-Castillo, J.; Moriones, E.; Hernández-Zepeda, C.; Idris, A.; Brown, J.; Murilo Zerbini, F.; Martin, D. Establishment of three new genera in the family Geminiviridae: Becurtovirus, Eragrovirus and Turncurtovirus. Arch. Virol. 2014, 159, 2193–2203. [Google Scholar] [CrossRef] [PubMed]
- Rojas, M.R.; Hagen, C.; Lucas, W.J.; Gilbertson, R.L. Exploiting chinks in the plant’s armor: Evolution and emergence of geminiviruses. Ann. Rev. Phytopathol. 2005, 43, 361–394. [Google Scholar] [CrossRef]
- Briddon, R.W.; Brown, J.K.; Moriones, E.; Stanley, J.; Zerbini, M.; Zhou, X.; Fauquet, C.M. Recommendations for the classification and nomenclature of the DNA-_ satellites of begomoviruses. Arch Virol. 2008, 153, 763–781. [Google Scholar] [CrossRef] [PubMed]
- Briddon, R.W.; Stanley, J. Sub-viral agents associated with plant-infecting single-stranded DNA viruses. Virology 2006, 344, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Saunders, K.; Briddon, R.W.; Stanley, J. Replication promiscuity of DNA-β satellites associated with monopartite begomoviruses; deletion mutagenesis of the Ageratum yellow vein virus DNA- β satellite localises sequences involved in replication. J. Gen. Virol. 2008, 89, 3165–3172. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Ikegami, M. RNA-silencing suppressors of geminiviruses. J. Gen. Plant Pathol. 2008, 74, 189–202. [Google Scholar] [CrossRef]
- Settlage, S.B.; See, R.G.; Hanley-Bowdoin, L. Geminivirus C3 protein: Replication enhancement and protein interactions. J. Virol. 2005, 79, 9885–9895. [Google Scholar] [CrossRef] [PubMed]
- Vanitharani, R.; Chellappan, P.; Pita, J.S.; Fauquet, C.M. Differential roles of AC2 and AC4 of cassava geminiviruses in mediating synergism and suppression of posttranscriptional gene silencing. J. Virol. 2004, 78, 9487–9498. [Google Scholar] [CrossRef] [PubMed]
- Krake, L.R.; Rezaian, M.A.; Dry, I.B. Expression of the tomato leaf curl geminivirus C4 gene produces viruslike symptoms in transgenic plants. Mol. Plant-Microbe Interact. 1998, 11, 413–417. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Settlage, S.B.; Robertson, D. Reprogramming plant gene expression: A prerequisite to geminivirus DNA replication. Mol. Plant Pathol. 2004, 5, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Orozco, B.M.; Kong, L.J.; Batts, L.A.; Elledge, S.; Hanley-Bowdoin, L. The multifunctional character of a geminivirus replication protein is reflected by its complex oligomerization properties. J. Biol. Chem. 2000, 275, 6114–6122. [Google Scholar] [CrossRef]
- Argüello-Astorga, G.R.; Guevara-González, L.R.; Herrera-Estrella, L.R.; Rivera-Bustamante, R.F. Geminivirus replication origins have a group-specific organization of iterative elements: A model for replication. Virology 1994, 203, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Argüello-Astorga, G.R.; Ruiz-Medrano, R. An iteron-related domain is associated to motif 1 in the replication proteins of geminiviruses: Identification of potential interacting amino acid-base pairs by a comparative approach. Arch. Virol. 2001, 146, 1465–1485. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Mansoor, S.; Khan, S.H.; Hussain, M.; Zafar, Y.; Pinner, M.S.; Briddon, R.W.; Stanley, J.; Markham, P.G. Association of a begomovirus and nanovirus-like molecule with Ageratum yellow vein disease in Pakistan. Plant Dis. 2000, 84, 101. [Google Scholar] [CrossRef]
- Tahir, M.; Haider, M.S.; Briddon, R.W. Complete nucleotide sequences of a distinct bipartite begomovirus, Bitter gourd yellow vein virus, infecting Momordica charantia. Arch. Virol. 2010, 155, 1901–1905. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.S.; Mansoor, S.; Briddon, R.W. Complete nucleotide sequences of cotton leaf curl Rajasthan virus and its associated DNA β molecule infecting tomato. Arch. Virol. 2007, 152, 2131–2134. [Google Scholar] [CrossRef] [PubMed]
- Briddon, R.W.; Markham, P.G. Universal primers for the PCR amplification of dicot-infecting geminiviruses. Mol. Biotechnol. 1994, 1, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Briddon, R.W.; Bull, S.E.; Mansoor, S.; Amin, I.; Markham, P.G. Universal primers for the PCR-mediated amplification of DNA β; a molecule associated with some monopartite begomoviruses. Mol. Biotechnol. 2002, 20, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Page, R.D.M. TREEVIEW: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar] [PubMed]
- Fauquet, C.M.; Briddon, R.W.; Brown, J.K.; Moriones, E.; Stanley, J.; Zerbini, M.; Zhou, X. Geminivirus strain demarcation and nomenclature. Arch. Virol. 2008, 153, 783–821. [Google Scholar] [CrossRef] [PubMed]
- Hellens, R.P.; Edwards, E.A.; Leyland, N.R.; Bean, S.; Mullineaux, P.M. pGreen: A versatile and flexible binary Ti vector for Agrobacterium-mediated plant transformation. Plant Mol. Biol. 2000, 42, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Briddon, R.W.; Watts, J.; Markham, P.G.; Stanley, J. The coat protein of beet curly top virus is essential for infectivity. Virology 1989, 172, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Fauquet, C.M.; Bisaro, D.M.; Briddon, R.W.; Brown, J.K.; Harrison, B.D.; Rybicki, E.P.; Stenger, D.C.; Stanley, J. Revision of taxonomic criteria for species demarcation in the family Geminiviridae, and an updated list of begomovirus species. Arch. Virol. 2003, 148, 405–421. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Gunapati, S.; Singh, S.P.; Gadre, R.; Sharma, N.C.; Tuli, R. Molecular characterization and pathogenicity of a carrot (Daucus carota) infecting begomovirus and associated betasatellite from India. Virus Res. 2013, 178, 478–485. [Google Scholar] [CrossRef]
- Kumar, Y.; Hallan, V.; Zaidi, A.A. Chilli leaf curl Palampur virus is a distinct begomovirus species associated with a betasatellite. Plant Pathol. 2011, 60, 1040–1047. [Google Scholar] [CrossRef]
- Kumar, Y.; Bhardwaj, P.; Hallan, V.; Zaidi, A. Detection and characterization of Ageratum enation virus and a nanovirus-like satellite DNA1 from zinnia causing leaf curl symptoms in India. J. Gen. Plant Pathol. 2010, 76, 395–398. [Google Scholar] [CrossRef]
- Raj, S.; Snehi, S.; Khan, M.; Tiwari, A.; Rao, G. Detection of Ageratum enation virus from cat’s whiskers (Cleome gynandra L.) with leaf curl symptoms in India. J. Gen. Plant. Pathol. 2010, 76, 292–294. [Google Scholar] [CrossRef]
- Briddon, R.W.; Bull, S.E.; Amin, I.; Idris, A.M.; Mansoor, S.; Bedford, I.D.; Dhawan, P.; Rishi, N.; Siwatch, S.S.; Abdel-Salam, A.M.; et al. Diversity of DNA β: A satellite molecule associated with some monopartite begomoviruses. Virology 2003, 312, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Marwal, A.; Sahu, A.; Choudhary, D.; Gaur, R.K. Complete nucleotide sequence of a begomovirus associated with satellites molecules infecting a new host Tagetes patula in India. Virus Genes 2013, 47, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Mubin, M.; Shahid, M.S.; Tahir, M.N.; Briddon, R.W.; Mansoor, S. Characterization of begomovirus components from a weed suggests that begomoviruses may associate with multiple distinct DNA satellites. Virus Genes 2010, 40, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Nawaz-ul-Rehman, M.S.; Mansoor, S.; Briddon, R.W.; Fauquet, C.M. Maintenance of an Old World betasatellite by a New World helper begomovirus and possible rapid adaptation of the betasatellite. J. Virol. 2009, 83, 9347–9355. [Google Scholar] [CrossRef] [PubMed]
- Qazi, J.; Amin, I.; Mansoor, S.; Iqbal, J.; Briddon, R.W. Contribution of the satellite encoded gene βC1 to cotton leaf curl disease symptoms. Virus Res. 2007, 128, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Amin, I.; Patil, B.L.; Briddon, R.W.; Mansoor, S.; Fauquet, C.M. A common set of developmental miRNAs are up regulated in Nicotiana benthamiana by diverse begomoviruses. Virol. J. 2011, 8, e143. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tahir, M.; Amin, I.; Haider, M.S.; Mansoor, S.; Briddon, R.W. Ageratum enation virus—A Begomovirus of Weeds with the Potential to Infect Crops. Viruses 2015, 7, 647-665. https://doi.org/10.3390/v7020647
Tahir M, Amin I, Haider MS, Mansoor S, Briddon RW. Ageratum enation virus—A Begomovirus of Weeds with the Potential to Infect Crops. Viruses. 2015; 7(2):647-665. https://doi.org/10.3390/v7020647
Chicago/Turabian StyleTahir, Muhammad, Imran Amin, Muhammad Saleem Haider, Shahid Mansoor, and Rob W. Briddon. 2015. "Ageratum enation virus—A Begomovirus of Weeds with the Potential to Infect Crops" Viruses 7, no. 2: 647-665. https://doi.org/10.3390/v7020647
APA StyleTahir, M., Amin, I., Haider, M. S., Mansoor, S., & Briddon, R. W. (2015). Ageratum enation virus—A Begomovirus of Weeds with the Potential to Infect Crops. Viruses, 7(2), 647-665. https://doi.org/10.3390/v7020647