
Archaeal Viruses Multiply: Temporal Screening in a Solar Saltern

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Growth Conditions
2.3. Isolation of Prokaryotes and Their Taxonomic Definition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample a | Nbr | Strain | 16S rRNA Gene Partial Sequence GenBank Acc. No. and Length (bp) | Reference |
|---|---|---|---|---|
| SSII | 1 | Halorubrum sp. SS6-1 | KJ917631 (1315) | This study |
| 2 | Halorubrum sp. SS6-2 | KJ917632 (1343) | This study | |
| 3 | Halolamina sp. SS6-3 | KJ917633 (996) | This study | |
| 4 | Halobacterium sp. SS6-4 | KJ917634 (1343) | This study | |
| 5 | Halobacterium sp. SS6-5 | KJ917635 (1354) | This study | |
| 6 | Halobellus sp. SS6-7 | KJ917636 (1331) | This study | |
| 7 | Haloarcula sp. SS7-2 | KJ917637 (1345) | This study | |
| 8 | Halorubrum sp. SS7-4 | JN971009 (1333) | [23] | |
| 9 | Haloarcula sp. SS8-1 | KJ917638 (1342) | This study | |
| 10 | Halorubrum sp. SS8-2 | KJ917639 (1338) | This study | |
| 11 | Haloarcula sp. SS8-3 | KJ917640 (1341) | This study | |
| 12 | Haloarcula sp. SS8-4 | KJ917641 (1335) | This study | |
| 13 | Haloarcula sp. SS8-5 | KJ917642 (1357) | This study | |
| 14 | Halorubrum sp. SS8-7 | KJ917643 (1292) | This study | |
| 15 | Haloterrigena sp. SS9-2 | KJ917644 (1323) | This study | |
| 16 | Halogeometricum sp. SS9-3 | KJ917645 (1323) | This study | |
| 17 | Halogranum sp. SS9-5 | KJ917646 (1328) | This study | |
| 18 | Haloferax sp. SS9-6 | KJ917647 (1326) | This study | |
| 19 | Halorubrum sp. SS9-12 | KJ917648 (1345) | This study | |
| SSIII | 20 | Halorubrum sp. SS10-3 | KJ917649 (1345) | This study |
| 21 | Haloarcula sp. SS10-4 | KJ917650 (1317) | This study | |
| 22 | Natrinema sp. SS10-5 | KJ917651 (1348) | This study | |
| 23 | Haloferax sp. SS10-6 | KJ917652 (1338) | This study | |
| 24 | Haloferax sp. SS10-7 | KJ917653 (1334) | This study | |
| 25 | Halorubrum sp. SS10-9 | KJ917654 (1342) | This study | |
| 26 | Halogranum sp. SS11-3 | KJ917655 (1342) | This study | |
| 27 | Halogranum sp. SS13-4 | KJ917656 (1352) | This study | |
| 28 | Halogranum sp. SS13-5 | KJ917657 (1353) | This study | |
| 29 | Halogranum sp. SS13-6 | KJ917658 (1329) | This study | |
| 30 | Haloterrigena sp. SS13-7 | KJ917659 (1344) | This study | |
| 31 | Haloterrigena sp. SS13-8 | KJ917660 (1343) | This study | |
| 32 | Haloterrigena sp. SS13-10 | KJ917661 (1348) | This study | |
| 33 | Halogranum sp. SS13-11 | KJ917662 (1349) | This study | |
| 34 | Halorubrum sp. SS13-12 | KJ917663 (1321) | This study | |
| 35 | Halorubrum sp. SS13-13 | KJ917664 (1306) | This study | |
| 36 | Haloarcula sp. SS13-14 | KJ917665 (1336) | This study | |
| CC | 37 | Halorubrum sp. SS1-3 | JN196470 (1330) | [22] |
| 38 | Halorubrum sp. SS5-4 | JN196482 (1401) | [22] | |
| 39 | Halorubrum sp. SP3-3 | JN196487 (1414) | [22] | |
| 40 | Haloarcula hispanica ATCC 33960 | U68541 | [34] | |
| 41 | “Haloarcula californiae” ATCC 33799 | AB477984 | [35] | |
| 42 | Haloarcula japonica TR1 ATCC 49778 | NR_028234 | [36] | |
| 43 | Haloarcula marismortui ATCC 43049 | X61688 | [37,38] | |
| 44 | Haloarcula quadrata ATCC 700850 | AB010964 | [39] | |
| 45 | “Haloarcula sinaiiensis” ATCC 33800 | D14129 | [35] | |
| 46 | Haloarcula vallismortis ATCC 29715 | AB355982 | [40,41] | |
| 47 | Halorubrum sodomense DSM 33755 | D13379 | [42] |
2.4. Isolation of Viruses and Their Characterization
2.5. Virus-Host Interactions Test
| Samplea | Virus Isolate No. | Virus | Nomenclature | Plaque Morphology | Stock Titer (pfu ml−1) | Chloroform Sensitivity | Morphotype | Isolation Host | Reference for the Virus | |
|---|---|---|---|---|---|---|---|---|---|---|
| SSII | SS9 | 1 | HRTV-13 | Halorubrum tailed virus 13 | Clear | 3.6 × 109 | NS | Myovirus | Halorubrum sp. SS8-2 | This study |
| SS9 | 2 | HRTV-14 | Halorubrum tailed virus 14 | Clear | 3.0 × 1010 | NS | Myovirus | Halorubrum sp. SS6-2 | This study | |
| SS6 | 3 | HRTV-15 | Halorubrum tailed virus 15 | Clear | 1.4 × 1010 | NS | Myovirus | Halorubrum sp. SS6-2 | This study | |
| SS7 | 4 | HRTV-16 | Halorubrum tailed virus 16 | Clear | 4.9 × 1010 | NS | Myovirus | Halorubrum sp. SS6-2 | This study | |
| SS6 | 5 | HRTV-17 | Halorubrum tailed virus 17 | Clear | 1.0 × 109 | NS | Myovirus | Halorubrum sp. SS9-12 | This study | |
| SSIII | SS10 | 6 | HSTV-4 | Halorubrum sodomense tailed virus 4 | Clear | 4.5 × 107 | NS | Myovirus | Halorubrum sodomense | This study |
| SS10 | 7 | HRTV-18 | Halorubrum tailed virus 18 | Clear | 1.4 × 109 | NS | Myovirus | Halorubrum sp. SS10-3 | This study | |
| SS13 | 8 | HRTV-19 | Halorubrum tailed virus 19 | Clear | 2.7 × 108 | NS | Myovirus | Halorubrum sp. SS10-3 | This study | |
| SS10 | 9 | HRTV-20 | Halorubrum tailed virus 20 | Turbid | 5.5 × 109 | NS | Myovirus | Halorubrum sp. SS10-9 | This study | |
| SS10 | 10 | HRTV-21 | Halorubrum tailed virus 21 | Turbid | 1.4 × 107 | NS | Myovirus | Halorubrum sp. SS10-9 | This study | |
| SS10 | 11 | HRTV-22 | Halorubrum tailed virus 22 | Clear | 5.7 × 109 | NS | Myovirus | Halorubrum sp. SS10-9 | This study | |
| SS13 | 12 | HRTV-23 | Halorubrum tailed virus 23 | Turbid | 5.8 × 109 | NS | Myovirus | Halorubrum sp. SS10-9 | This study | |
| SS13 | 13 | HRTV-24 | Halorubrum tailed virus 24 | Turbid | 2.6 × 108 | NS | Myovirus | Halorubrum sp. SS10-9 | This study | |
| SS10 | 14 | HRTV-25 | Halorubrum tailed virus 25 | Turbid | 1.2 × 1011 | NS | Myovirus | Halorubrum sp. SS13-12 | This study | |
| SS10 | 15 | HCTV-6 | “Haloarcula californiae” tailed virus 6 | Clear | 1.3 × 109 | NS | Myovirus | “Haloarcula californiae” | This study | |
| SS10 | 16 | HCTV-7 | “Haloarcula californiae” tailed virus 7 | Turbid | 1.3 × 1010 | NS | Myovirus | “Haloarcula californiae” | This study | |
| SS10 | 17 | HCTV-8 | “Haloarcula californiae” tailed virus 8 | Turbid | 5.0 × 1010 | NS | Myovirus | “Haloarcula californiae” | This study | |
| SS10 | 18 | HCTV-9 | “Haloarcula californiae” tailed virus 9 | Turbid | 2.6 × 1010 | NS | Myovirus | “Haloarcula californiae” | This study | |
| SS10 | 19 | HCTV-10 | “Haloarcula californiae” tailed virus 10 | Turbid | 1.5 × 1010 | NS | Myovirus | “Haloarcula californiae” | This study | |
| SS10 | 20 | HCTV-11 | “Haloarcula californiae” tailed virus 11 | Turbid | 4.3 × 1010 | NS | Myovirus | “Haloarcula californiae” | This study | |
| SS11 | 21 | HCTV-12 | “Haloarcula californiae” tailed virus 12 | Turbid | 8.0 × 1010 | NS | Myovirus | “Haloarcula californiae” | This study | |
| SS12 | 22 | HCTV-13 | “Haloarcula californiae” tailed virus 13 | Clear | 3.0 × 109 | NS | Myovirus | “Haloarcula californiae” | This study | |
| SS12 | 23 | HCTV-14 | “Haloarcula californiae” tailed virus 14 | Turbid | 4.5 × 1010 | NS | Myovirus | “Haloarcula californiae” | This study | |
| SS12 | 24 | HCTV-15 | “Haloarcula californiae” tailed virus 15 | Clear | 7.0 × 1010 | NS | Myovirus | “Haloarcula californiae” | This study | |
| SS10 | 25 | HJTV-3 | Haloarcula japonica tailed virus 3 | Clear | 7.1 × 108 | NS | Myovirus | Haloarcula japonica | This study | |
| SS10 | 26 | HRTV-26 | Halorubrum tailed virus 26 | Turbid | 1.7 × 108 | NS | Myovirus | Halorubrum sp. SS13-13 | This study | |
| SS13 | 27 | HRTV-27 | Halorubrum tailed virus 27 | Clear | 5.0 × 106 | NS | Myovirus | Halorubrum sp. SS13-13 | This study | |
| SSII | SS6 | 28 | HRTV-28 | Halorubrum tailed virus 28 | Turbid | 6.0 × 1010 | NS | Siphovirus | Halorubrum sp. SS8-7 | This study |
| SS7 | 29 | HRTV-29 | Halorubrum tailed virus 29 | Turbid | 4.5 × 1012 | NS | Siphovirus | Halorubrum sp. SS7-4 | This study | |
| SS7 | 30 | HATV-3 | Haloarcula tailed virus 3 | Turbid | 1.0 × 1011 | NS | Siphovirus | Haloarcula sp. SS8-5 | This study | |
| SSIII | SS10 | 31 | HCTV-16 | “Haloarcula californiae” tailed virus 25 | Turbid | 1.5 × 1011 | NS | Siphovirus | “Haloarcula californiae” | This study |
| SSII | SS8 | 32 | HRPV-6 | Halorubrum pleomorphic virus 6 | Turbid | 1.1 × 1012 | NS | Pleomorphic | Halorubrum sp. SS7-4 | [23] |
| SS6 | 33 | HRPV-7 | Halorubrum pleomorphic virus 7 | Turbid | 8.7 × 1010 | −1 log | Pleomorphic | Halorubrum sp. SS5-4 | This study | |
| SS8 | 34 | HRPV-8 | Halorubrum pleomorphic virus 8 | Turbid | 4.9 × 109 | NS | Pleomorphic | Halorubrum sp. SP3-3 | This study | |
| SSIII | SS13 | 35 | HAPV-2 | Haloarcula pleomorphic virus 2 | Turbid | 1.6 × 109 | −3 log | Pleomorphic | Haloarcula sp. SS13-14 | This study |
| SSIII | SS13 | 36 | HCIV-1 | “Haloarcula californiae” icosahedral virus 1 | Clear | 5.8 × 1010 | −2 log | Icosahedral | “Haloarcula californiae” | This study |
3. Results
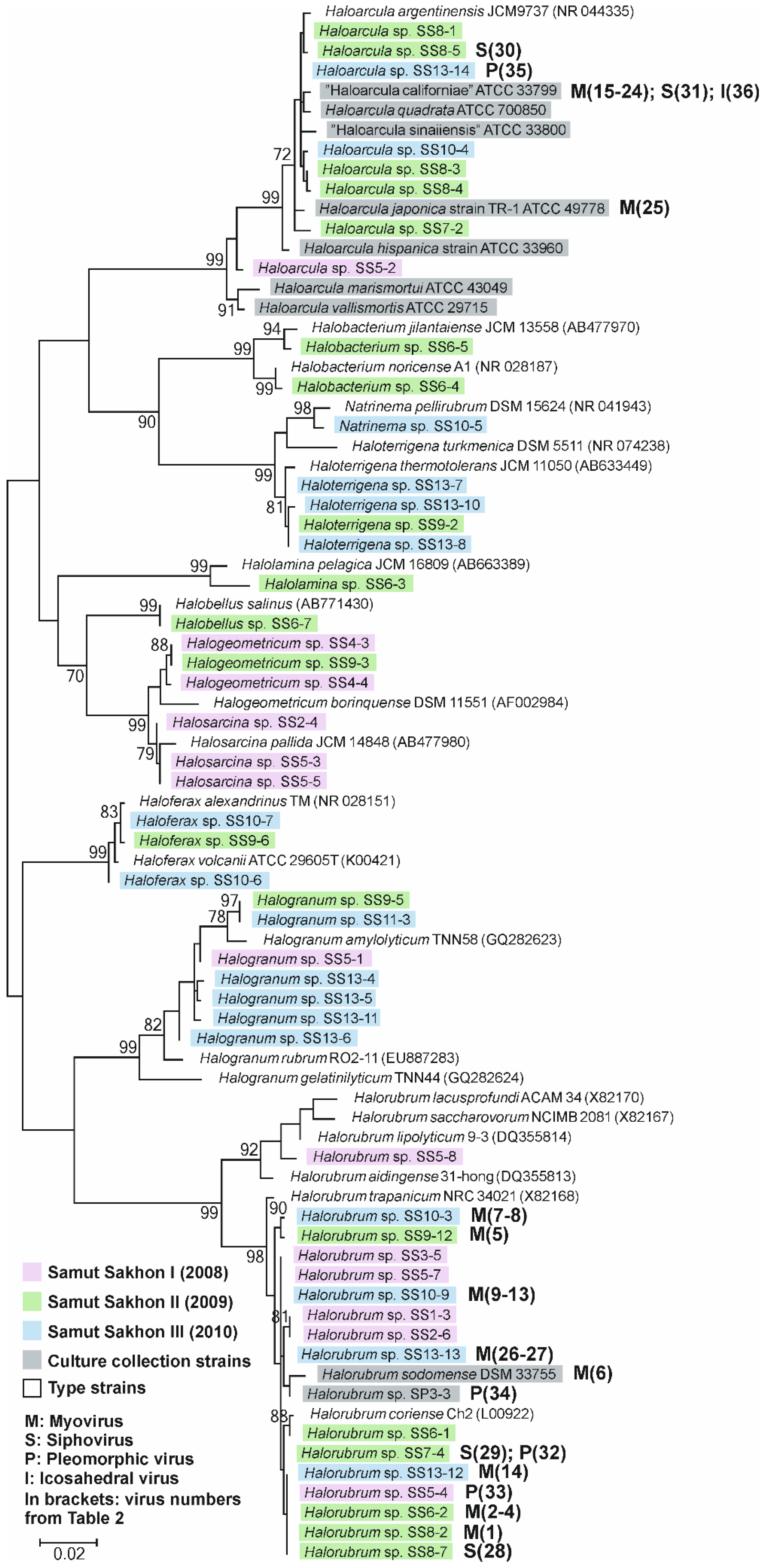
3.1. All Isolated Archaeal Strains Belonged to Ten Genera of the Halobacteriaceae Family
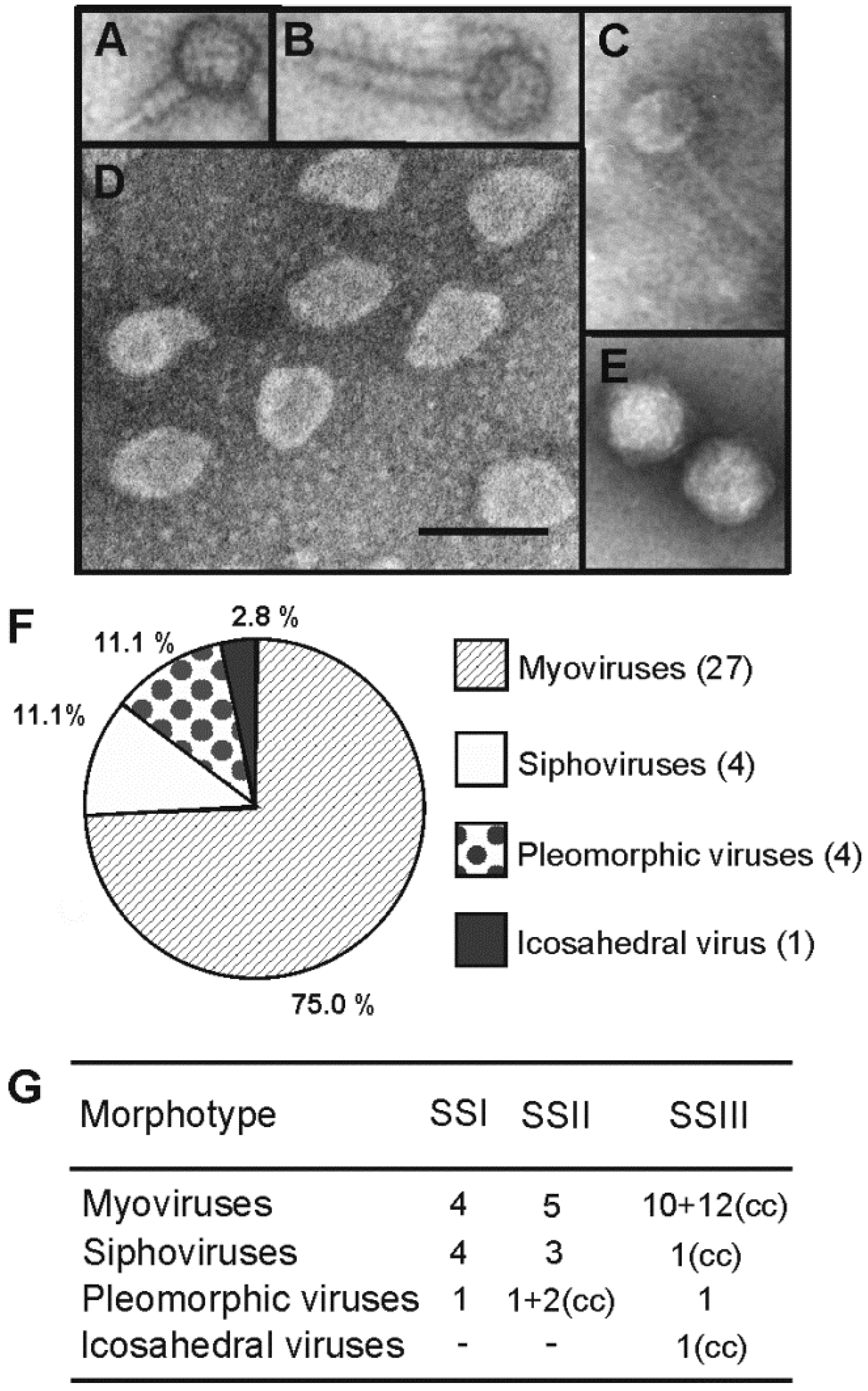
3.2. Thirty Six Euryarchaeal Virus Isolates Were Assigned to Known Virus Morphotypes


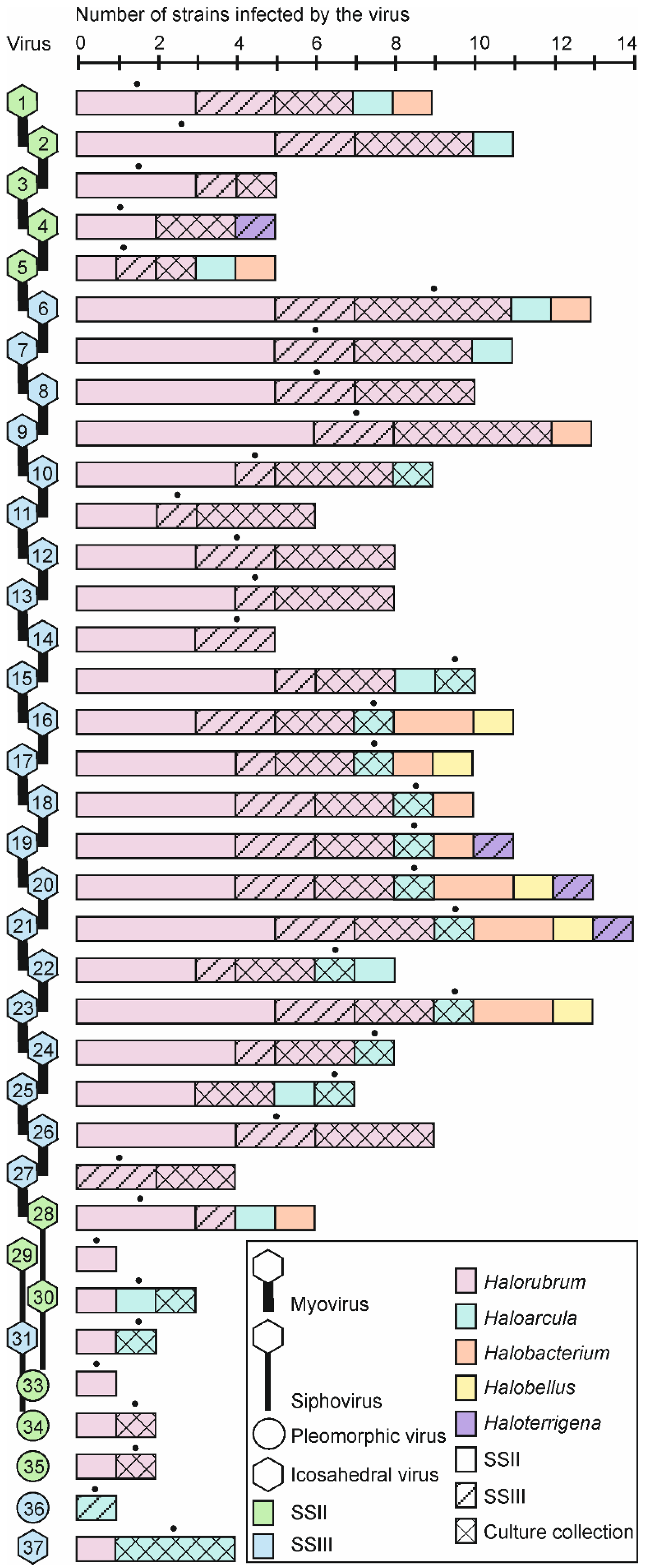
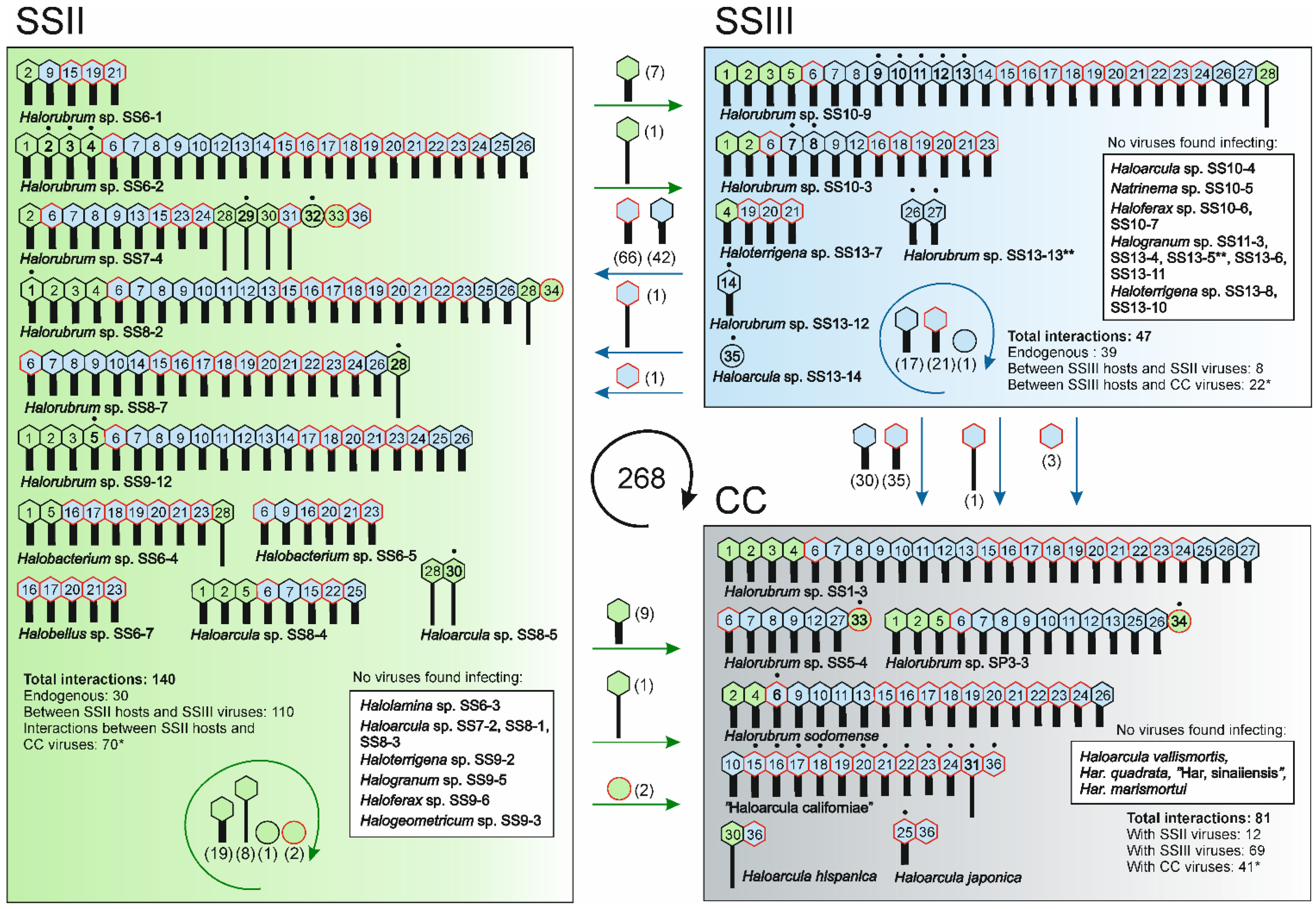
3.3. Multiple Virus-Host Interactions, in which Myoviruses Were the Most Promiscuous, Were Observed
3.4. Broad Host Ranges Covering Archaea from Different Genera Were Characteristic to Many Myovirus Isolates

3.5. Halorubrum Strains Were the Most Susceptible to Viruses
3.6. Virus-Host Interactions Were Observed Within and Across Samples

| Hosts | Viruses | ||||||
|---|---|---|---|---|---|---|---|
| SSII (2009) | SSIII (2010) | ||||||
| Myoviruses | Sipho-, Pleomorphic, icosahedral Viruses | Sipho-, Pleomorphic, Icosahedral Viruses (cc a) | Myoviruses | Myoviruses (cc) | Sipho-, Pleomorphic, Icosahedral Viruses | Sipho-, Pleomorphic, Icosahedral Viruses (cc) | |
| SSII | 19 (3.8)b | 9 (2.3) | 2 (0.5) | 42 (4.2) | 66 (6.0) | - | 2 (1.0) |
| SSIII | 7 (1.4) | 1 (0.3) | - | 17 (1.7) | 21 (1.9) | 1 (1.0) | - |
| CC | 9 (1.8) | 1 (0.3) | 2 (0.5) | 30 (3.0) | 35 (3.1) | - | 4 (0.5) |
4. Discussion
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pina, M.; Bize, A.; Forterre, P.; Prangishvili, D. The archeoviruses. FEMS Microbiol. Rev. 2011, 35, 1035–1054. [Google Scholar] [CrossRef] [PubMed]
- Roine, E.; Oksanen, H.M. Viruses from the hypersaline environment. In Halophiles and Hypersaline Environments: Current Research and Future Trends; Ventosa, A., Oren, A., Ma, Y., Eds.; Springer: Heidelberg, Germany, 2011; pp. 153–172. [Google Scholar]
- Emerson, J.B.; Thomas, B.C.; Andrade, K.; Allen, E.E.; Heidelberg, K.B.; Banfield, J.F. Dynamic viral populations in hypersaline systems as revealed by metagenomic assembly. Appl. Environ. Microbiol. 2012, 78, 6309–6320. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.; Yarza, P.; Parro, V.; Meseguer, I.; Rosselló-Móra, R.; Antón, J. Culture-independent approaches for studying viruses from hypersaline environments. Appl. Environ. Microbiol. 2012, 78, 1635–1643. [Google Scholar] [CrossRef] [PubMed]
- Prangishvili, D. The wonderful world of archaeal viruses. Annu. Rev. Microbiol. 2013, 67, 565–585. [Google Scholar] [CrossRef] [PubMed]
- Pietilä, M.K.; Demina, T.A.; Atanasova, N.S.; Oksanen, H.M.; Bamford, D.H. Archaeal viruses and bacteriophages: Comparisons and contrasts. Trends Microbiol. 2014, 22, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Senčilo, A.; Roine, E. A glimpse of the genomic diversity of haloarchaeal tailed viruses. Front. Microbiol. 2014, 5, e84. [Google Scholar]
- Porter, K.; Kukkaro, P.; Bamford, J.K.; Bath, C.; Kivelä, H.M.; Dyall-Smith, M.L.; Bamford, D.H. SH1: A novel, spherical halovirus isolated from an Australian hypersaline lake. Virology 2005, 335, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Bath, C.; Cukalac, T.; Porter, K.; Dyall-Smith, M.L. His1 and His2 are distantly related, spindle-shaped haloviruses belonging to the novel virus group. Salterprovirus. Virol. 2006, 350, 228–239. [Google Scholar] [CrossRef]
- Pietilä, M.K.; Roine, E.; Paulin, L.; Kalkkinen, N.; Bamford, D.H. An ssDNA virus infecting archaea: A new lineage of viruses with a membrane envelope. Mol. Microbiol. 2009, 72, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Guixa-Boixareu, N.; Calderón-Paz, J.; Heldal, M.; Bratbak, G.; Pedrós-Alió, C. Viral lysis and bacterivory as prokaryotic loss factors along a salinity gradient. Aquat. Microb. Ecol. 1996, 11, 215–227. [Google Scholar] [CrossRef]
- Oren, A.; Bratbak, G.; Heldal, M. Occurrence of virus-like particles in the Dead Sea. Extremophiles 1997, 1, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Steward, G.; Jellison, R.; Chu, W.; Choi, S. Abundance, distribution, and diversity of viruses in alkaline, hypersaline Mono Lake, California. Microb. Ecol. 2004, 47, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Pietilä, M.K.; Atanasova, N.S.; Oksanen, H.M.; Bamford, D.H. Modified coat protein forms the flexible spindle-shaped virion of haloarchaeal virus His1. Environ. Microbiol. 2013, 15, 1674–1686. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.; Pietilä, M.K.; Fu, C.J.; Schmid, M.F.; Bamford, D.H.; Chiu, W. Lemon-shaped virus His1 with uniform tail but variable capsid structure. Proc. Natl. Acad. Sci. USA 2015, 112, 2449–2454. [Google Scholar] [CrossRef] [PubMed]
- Pietilä, M.K.; Laurinmäki, P.; Russell, D.A.; Ko, C.C.; Jacobs-Sera, D.; Hendrix, R.W.; Bamford, D.H.; Butcher, S.J. Structure of the archaeal head-tailed virus HSTV-1 completes the HK97 fold story. Proc. Natl. Acad. Sci. USA 2013, 110, 10604–10609. [Google Scholar] [CrossRef] [PubMed]
- Benson, S.D.; Bamford, J.K.; Bamford, D.H.; Burnett, R.M. Viral evolution revealed by bacteriophage PRD1 and human adenovirus coat protein structures. Cell 1999, 98, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Bamford, D.H. Do viruses form lineages across different domains of life? Res. Microbiol. 2003, 154, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Khayat, R.; Tang, L.; Larson, E.T.; Lawrence, C.M.; Young, M.; Johnson, J.E. Structure of an archaeal virus capsid protein reveals a common ancestry to eukaryotic and bacterial viruses. Proc. Natl. Acad. Sci. USA 2005, 102, 18944–18949. [Google Scholar] [CrossRef] [PubMed]
- Abrescia, N.G.; Bamford, D.H.; Grimes, J.M.; Stuart, D.I. Structure unifies the viral universe. Annu. Rev. Biochem. 2012, 81, 795–822. [Google Scholar] [CrossRef] [PubMed]
- Jaakkola, S.T.; Penttinen, R.K.; Vilen, S.T.; Jalasvuori, M.; Rönnholm, G.; Bamford, J.K.; Bamford, D.H.; Oksanen, H.M. Closely related archaeal Haloarcula hispanica icosahedral viruses HHIV-2 and SH1 have nonhomologous genes encoding host recognition functions. J. Virol. 2012, 86, 4734–4742. [Google Scholar] [CrossRef] [PubMed]
- Atanasova, N.S.; Roine, E.; Oren, A.; Bamford, D.H.; Oksanen, H.M. Global network of specific virus-host interactions in hypersaline environments. Environ. Microbiol. 2012, 14, 426–440. [Google Scholar] [CrossRef] [PubMed]
- Pietilä, M.K.; Atanasova, N.S.; Manole, V.; Liljeroos, L.; Butcher, S.J.; Oksanen, H.M.; Bamford, D.H. Virion architecture unifies globally distributed pleolipoviruses infecting halophilic archaea. J. Virol. 2012, 86, 5067–5079. [Google Scholar] [CrossRef] [PubMed]
- Senčilo, A.; Paulin, L.; Kellner, S.; Helm, M.; Roine, E. Related haloarchaeal pleomorphic viruses contain different genome types. Nucleic Acids Res. 2012, 40, 5523–5534. [Google Scholar] [CrossRef] [PubMed]
- Dyall-Smith, M.; Tang, S.L.; Bath, C. Haloarchaeal viruses: How diverse are they? Res. Microbiol. 2003, 154, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Sandaa, R.; Skjoldal, E.F.; Bratbak, G. Virioplankton community structure along a salinity gradient in a solar saltern. Extremophiles 2003, 7, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Brito, B.; Li, L.; Wegley, L.; Furlan, M.; Angly, F.; Breitbart, M.; Buchanan, J.; Desnues, C.; Dinsdale, E.; Edwards, R. Viral and microbial community dynamics in four aquatic environments. ISME J. 2010, 4, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Emerson, J.B.; Thomas, B.C.; Andrade, K.; Heidelberg, K.B.; Banfield, J.F. New approaches indicate constant viral diversity despite shifts in assemblage structure in an Australian hypersaline lake. Appl. Environ. Microbiol. 2013, 79, 6755–6764. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Valera, F.; Martin-Cuadrado, A.; Rodriguez-Brito, B.; Pašić, L.; Thingstad, T.F.; Rohwer, F.; Mira, A. Explaining microbial population genomics through phage predation. Nat. Rev. Microbiol. 2009, 7, 828–836. [Google Scholar] [CrossRef] [PubMed]
- Ventosa, A.; Fernández, A.B.; León, M.J.; Sánchez-Porro, C.; Rodriguez-Valera, F. The Santa Pola saltern as a model for studying the microbiota of hypersaline environments. Extremophiles 2014, 18, 811–824. [Google Scholar] [CrossRef]
- Nuttall, S.D.; Dyall-Smith, M.L. HF1 and HF2: Novel bacteriophages of halophilic archaea. Virology 1993, 197, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Dyall-Smith, M. The Halohandbook. 2009. Available online: http://www.haloarchaea.com/resources/halohandbook/ (accessed on 27 January 2015).
- Olkkonen, V.M.; Bamford, D.H. Quantitation of the adsorption and penetration stages of bacteriophage φ6 infection. Virology 1989, 171, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Juez, G.; Rodriguez-Valera, F.; Ventosa, A.; Kushner, D.J. Haloarcula hispanica spec. nov. and Haloferax gibbonsii spec, nov., two new species of extremely halophilic archaebacteria. Syst. Appl. Microbiol. 1986, 8, 75–79. [Google Scholar] [CrossRef]
- Javor, B.; Requadt, C.; Stoeckenius, W. Box-shaped halophilic bacteria. J. Bacteriol. 1982, 151, 1532–1542. [Google Scholar] [PubMed]
- Takashina, T.; Hamamoto, T.; Otozai, K.; Grant, W.D.; Horikoshi, K. Haloarcula japonica sp. nov., a new triangular halophilic archaebacterium. Syst. Appl. Microbiol. 1990, 13, 177–181. [Google Scholar] [CrossRef]
- Oren, A.; Ginzburg, M.; Ginzburg, B.Z.; Hochstein, L.I.; Volcani, B.E. Haloarcula marismortui (Volcani) sp. nov., nom. rev., an extremely halophilic bacterium from the Dead Sea. Int. J. Syst. Bacteriol. 1990, 40, 209–210. [Google Scholar] [CrossRef] [PubMed]
- Mylvaganam, S.; Dennis, P.P. Sequence heterogeneity between the two genes encoding 16S rRNA from the halophilic archaebacterium Haloarcula marismortui. Genetics 1992, 130, 399–410. [Google Scholar] [PubMed]
- Oren, A.; Ventosa, A.; Gutierrez, M.C.; Kamekura, M. Haloarcula quadrata sp. nov., a square, motile archaeon isolated from a brine pool in Sinai (Egypt). Int. J. Syst. Bacteriol. 1999, 49, 1149–1155. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, C.; Gutierrez, C.; Ramirez, C. Halobacterium vallismortis sp. nov. An amylolytic and carbohydrate-metabolizing, extremely halophilic bacterium. Can. J. Microbiol. 1978, 24, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Torreblanca, M.; Rodriguez-Valera, F.; Juez, G.; Ventosa, A.; Kamekura, M.; Kates, M. Classification of non-alkaliphilic halobacteria based on numerical taxonomy and polar lipid composition, and description of Haloarcula gen. nov. and Haloferax gen. nov. Syst. Appl. Microbiol. 1986, 8, 89–99. [Google Scholar] [CrossRef]
- Oren, A. Halobacterium sodomense sp. nov., a Dead Sea halobacterium with an extremely high magnesium requirement. Int. J. Syst. Bacteriol. 1983, 33, 381–386. [Google Scholar] [CrossRef]
- Arahal, D.R.; Dewhirst, F.E.; Paster, B.J.; Volcani, B.E.; Ventosa, A. Phylogenetic analyses of some extremely halophilic archaea isolated from Dead Sea water, determined on the basis of their 16S rRNA sequences. Appl. Environ. Microbiol. 1996, 62, 3779–3786. [Google Scholar] [PubMed]
- Edwards, U.; Rogall, T.; Blocker, H.; Emde, M.; Bottger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Schägger, H.; von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E.; Goebel, B. Taxonomic note: A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Bacteriol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
- Schwarzer, D.; Buettner, F.F.; Browning, C.; Nazarov, S.; Rabsch, W.; Bethe, A.; Oberbeck, A.; Bowman, V.D.; Stummeyer, K.; Mühlenhoff, M.; et al. A multivalent adsorption apparatus explains the broad host range of phage phi92: A comprehensive genomic and structural analysis. J. Virol. 2012, 86, 10384–10398. [Google Scholar] [CrossRef] [PubMed]
- Stummeyer, K.; Schwarzer, D.; Claus, H.; Vogel, U.; Gerardy-Schahn, R.; Mühlenhoff, M. Evolution of bacteriophages infecting encapsulated bacteria: Lessons from Escherichia coli K1-specific phages. Mol. Microbiol. 2006, 60, 1123–1135. [Google Scholar] [CrossRef] [PubMed]
- Sime-Ngando, T.; Lucas, S.; Robin, A.; Tucker, K.P.; Colombet, J.; Bettarel, Y.; Desmond, E.; Gribaldo, S.; Forterre, P.; Breitbart, M. Diversity of virus-host systems in hypersaline Lake Retba, Senegal. Environ. Microbiol. 2011, 13, 1956–1972. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Emerson, J.B.; Andrade, K.; Thomas, B.C.; Norman, A.; Allen, E.E.; Heidelberg, K.B.; Banfield, J.F. Virus-host and CRISPR dynamics in archaea-dominated Hypersaline Lake Tyrrell, Victoria, Australia. Archaea 2013, 13, e370871. [Google Scholar]
- Dyall-Smith, M.L.; Pfeiffer, F.; Klee, K.; Palm, P.; Gross, K.; Schuster, S.C.; Rampp, M.; Oesterhelt, D. Haloquadratum walsbyi: Limited diversity in a global pond. PLOS ONE 2011, 6, e20968. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, R.M.; Marceau, C.D.; Marceau, J.O.; Morris, S.; Clore, A.J.; Stedman, K.M. Differential virus host-ranges of the Fuselloviridae of hyperthermophilic Archaea: implications for evolution in extreme environments. Front. Microbiol. 2012, 3, e295. [Google Scholar] [CrossRef]
- Atanasova, N.S.; Pietilä, M.K.; Oksanen, H.M. Diverse antimicrobial interactions of halophilic archaea and bacteria extend over geographical distances and cross the domain barrier. Microbiol. Open 2013, 2, 811–825. [Google Scholar]
- Senčilo, A.; Jacobs-Sera, D.; Russell, D.A.; Ko, C.; Bowman, C.A.; Atanasova, N.S.; Österlund, E.; Oksanen, H.M.; Bamford, D.H.; Hatfull, G.F. Snapshot of haloarchaeal tailed virus genomes. RNA Biol. 2013, 10, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Bamford, D.H.; Burnett, R.M.; Stuart, D.I. Evolution of viral structure. Theor. Popul. Biol. 2002, 61, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Krupovič, M.; Bamford, D.H. Order to the viral universe. J. Virol. 2010, 84, 12476–12479. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atanasova, N.S.; Demina, T.A.; Buivydas, A.; Bamford, D.H.; Oksanen, H.M. Archaeal Viruses Multiply: Temporal Screening in a Solar Saltern. Viruses 2015, 7, 1902-1926. https://doi.org/10.3390/v7041902
Atanasova NS, Demina TA, Buivydas A, Bamford DH, Oksanen HM. Archaeal Viruses Multiply: Temporal Screening in a Solar Saltern. Viruses. 2015; 7(4):1902-1926. https://doi.org/10.3390/v7041902
Chicago/Turabian StyleAtanasova, Nina S., Tatiana A. Demina, Andrius Buivydas, Dennis H. Bamford, and Hanna M. Oksanen. 2015. "Archaeal Viruses Multiply: Temporal Screening in a Solar Saltern" Viruses 7, no. 4: 1902-1926. https://doi.org/10.3390/v7041902
APA StyleAtanasova, N. S., Demina, T. A., Buivydas, A., Bamford, D. H., & Oksanen, H. M. (2015). Archaeal Viruses Multiply: Temporal Screening in a Solar Saltern. Viruses, 7(4), 1902-1926. https://doi.org/10.3390/v7041902