
Genome Variability and Gene Content in Chordopoxviruses: Dependence on Microsatellites

Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome Sequences and Assessment of Gene Content

{kind=link}
{kind=link}
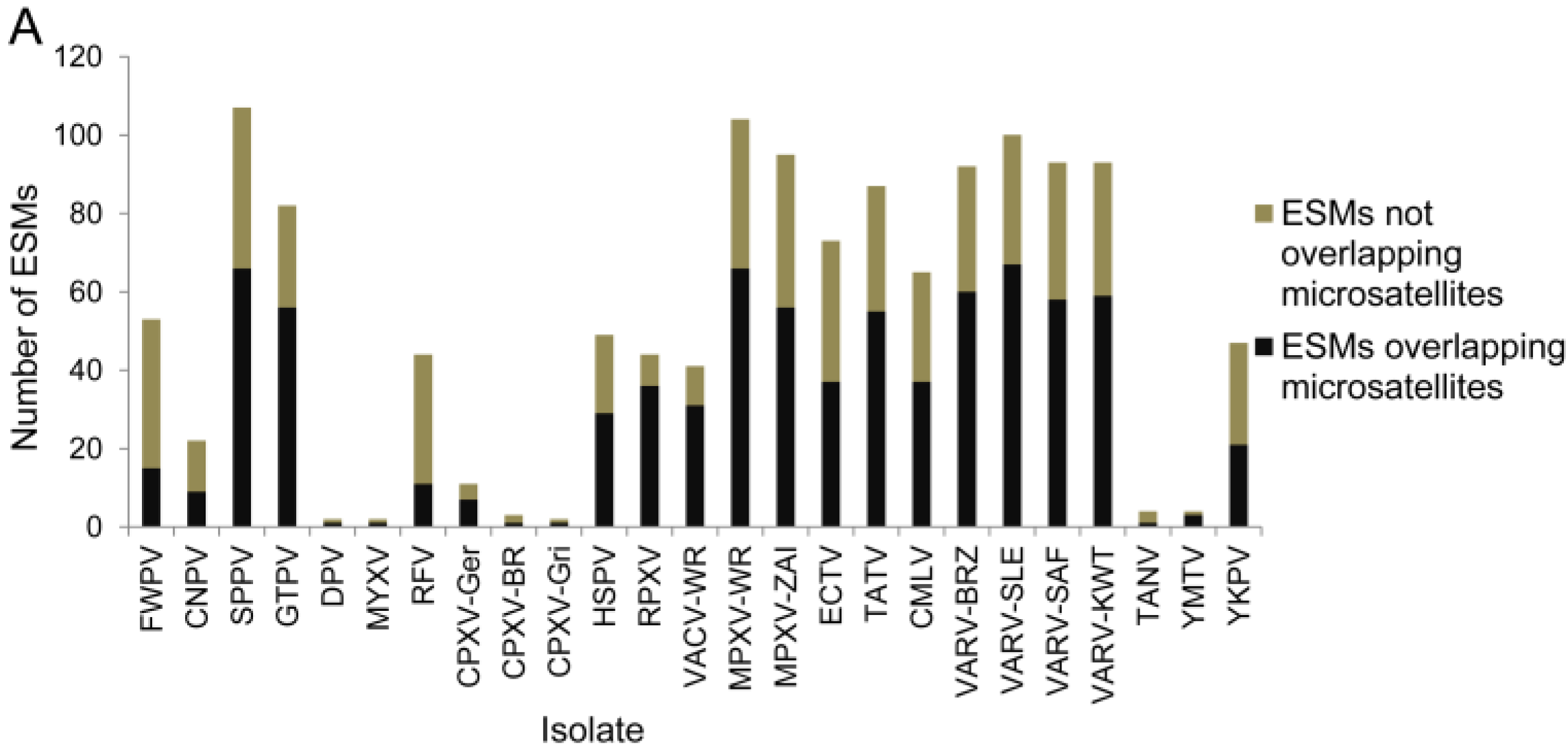{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Abbreviation | NCBI strain | Accession no. | Intact genes | Truncated genes | Fragmented genes | Length (bp) |
|---|---|---|---|---|---|---|---|
| Species | |||||||
| Avipoxvirus | |||||||
| Fowlpox virus | FWPV | Fowlpox virus strain Iowa | NC_002188 | 231 | 5 | 13 | 288,539 |
| Canarypox virus | CNPV | Canarypox virus strain ATCC VR111 | NC_005309 | 316 | 2 | 4 | 359,853 |
| Capripoxvirus | |||||||
| Sheeppox virus | SPPV | Sheeppox virus strain TU-V02127 | NC_004002 | 150 | 1 | 8 | 149,955 |
| Goatpox virus | GTPV | Goatpox virus strain Pellor | NC_004003 | 150 | 2 | 7 | 149,599 |
| Lumpy skin disease virus | LSDV | Lumpy skin disease virus strain Neethling 2490 | NC_003027 | 159 | 0 | 0 | 150,773 |
| Cervidpoxvirus | |||||||
| Mule deerpox virus | DPV | Deerpox virus W-848-83 | NC_006966 | 172 | 0 | 1 | 166,259 |
| Crocodylidpoxvirus | |||||||
| Nile crocodilepox virus | CRV | Crocodilepox virus strain Zimbabwe | NC_008030 | 175 | 0 | 0 | 190,054 |
| Leporipoxvirus | |||||||
| Myxoma virus | MYXV | Myxoma virus strain Lausanne | NC_001132 | 166 | 0 | 1 | 161,773 |
| Rabbit fibroma virus | RFV | Rabbit fibroma virus strain Kasza | NC_001266 | 161 | 0 | 4 | 159,857 |
| Molluscipoxvirus | |||||||
| Molluscum contagiosum virus | MOCV | Molluscum contagiosum virus strain subtype 1 | NC_001731 | 159 | 0 | 0 | 190,289 |
| Orthopoxvirus | |||||||
| Cowpox virus | CPXV-Ger | Cowpox virus strain Germany 91-3 | DQ437593 | 208 | 3 | 3 | 228,250 |
| Cowpox virus | CPXV-BR | Cowpox virus strain Brighton Red | AF482758 | 206 | 3 | 1 | 224,499 |
| Cowpox virus | CPXV-Gri | Cowpox virus strain GRI-90 | X94355 | 209 | 3 | 0 | 223,666 |
| Vaccinia virus | HSPV | Horsepox virus strain MNR-76 | DQ792504 | 181 | 22 | 4 | 212,633 |
| Vaccinia virus | RPXV | Rabbitpox virus strain Utrecht | AY484669 | 179 | 13 | 4 | 197,731 |
| Vaccinia virus | VACV-WR | Vaccinia virus strain WR (Western Reserve) | AY243312 | 178 | 12 | 3 | 194,711 |
| Monkeypox virus | MPXV-WR | Monkeypox virus strain MPXV-WRAIR7-61; Walter Reed 267 | AY603973 | 175 | 7 | 14 | 199,195 |
| Monkeypox virus | MPXV-ZAI | Monkeypox virus strain Zaire-96-I-16 | AF380138 | 176 | 7 | 14 | 196,858 |
| Ectromelia virus | ECTV | Ectromelia virus strain Moscow | AF012825 | 172 | 21 | 7 | 209,771 |
| Taterapox virus | TATV | Taterapox virus strain Dahomey 1968 | DQ437594 | 163 | 26 | 7 | 198,050 |
| Camelpox virus | CMLV | Camelpox virus strain M-96 from Kazakhstan | AF438165 | 174 | 14 | 8 | 205,719 |
| Variola virus | VARV-BRZ | Variola virus strain Brazil 1966 (v66-39 Sao Paulo) | DQ441419 | 162 | 18 | 12 | 188,062 |
| Variola virus | VARV-SLE | Variola virus strain Sierra Leone 1969 (V68-258) | DQ441437 | 162 | 17 | 11 | 187,014 |
| Variola virus | VARV-SAF | Variola virus strain South Africa 1965 (103 T'vaal, Nelspruit) | DQ441436 | 162 | 17 | 11 | 185,881 |
| Variola virus | VARV-KWT | Variola virus strain Kuwait 1967 (K1629) | DQ441433 | 162 | 17 | 11 | 185,853 |
| Parapoxvirus | |||||||
| Orf virus | ORFV | Orf virus strain OV-SA00 | NC_005336 | 134 | 0 | 0 | 139,962 |
| Bovine papular stomatitis virus | BPSV | Bovine papular stomatitis virus strain BV-AR02 | NC_005337 | 136 | 0 | 0 | 134,431 |
| Suipoxvirus | |||||||
| Swinepox virus | SWPV | Swinepox virus strain Nebraska 17077-99 | NC_003389 | 151 | 0 | 0 | 146,454 |
| Yatapoxvirus | |||||||
| Tanapox virus | TANV | Tanapox virus strain Kenya | NC_009888 | 151 | 2 | 0 | 144,565 |
| Yaba monkey tumor virus | YMTV | Yaba monkey tumor virus strain Amano | NC_005179 | 139 | 2 | 0 | 134,721 |
| Unassigned | |||||||
| Yoka poxvirus (not classified) | YKPV | Yoka poxvirus | NC_015960.1 | 164 | 12 | 5 | 175,699 |
| Squirrelpox virus | SQPV | Squirrel poxvirus strain Red squirrel UK | NC_022563.1 | 142 | 0 | 0 | 148,803 |
2.2. Detection of Early Stop Mutations (ESMs)
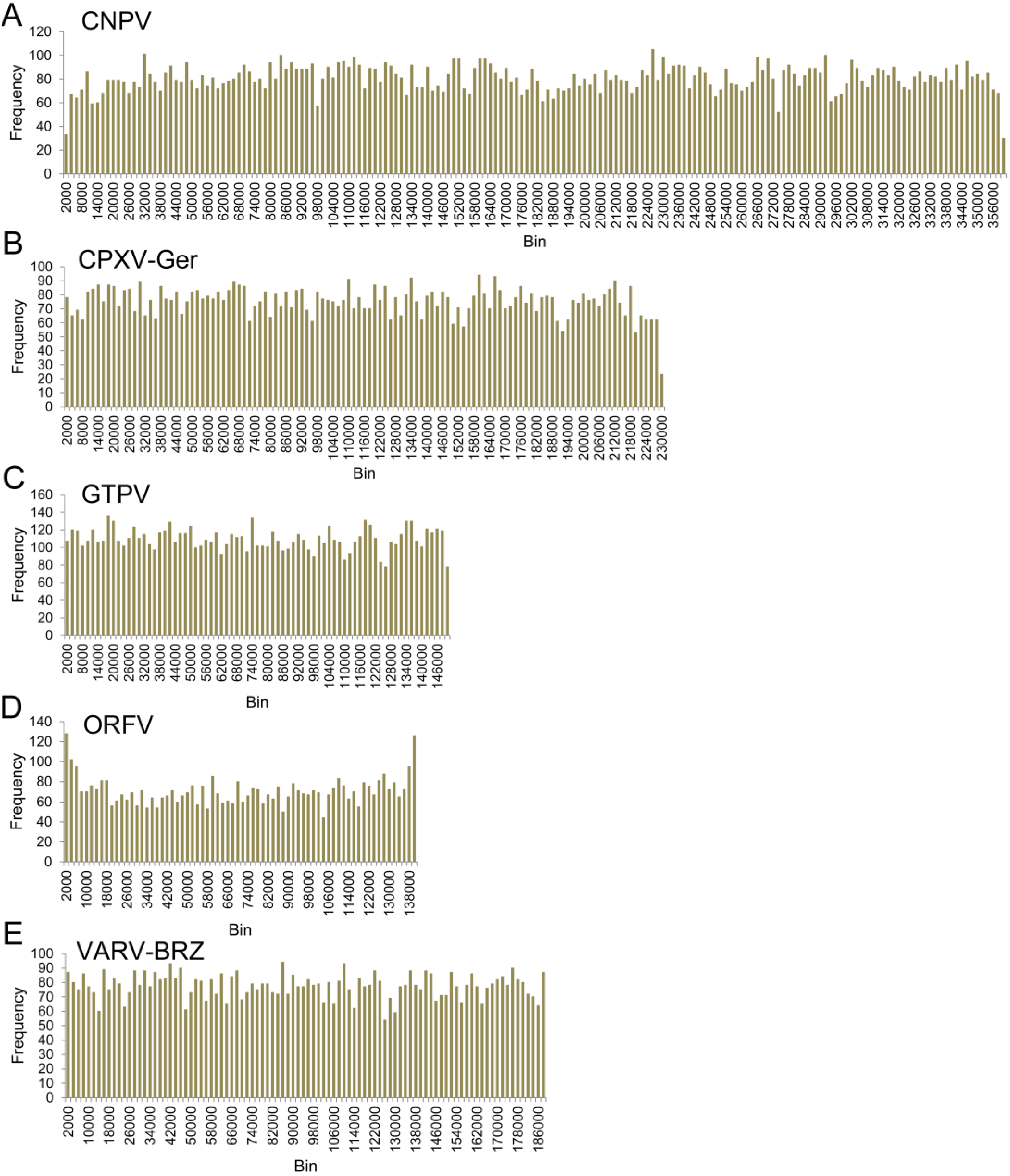
2.3. Microsatellite Identification
2.4. Statistical Analysis
3. Results
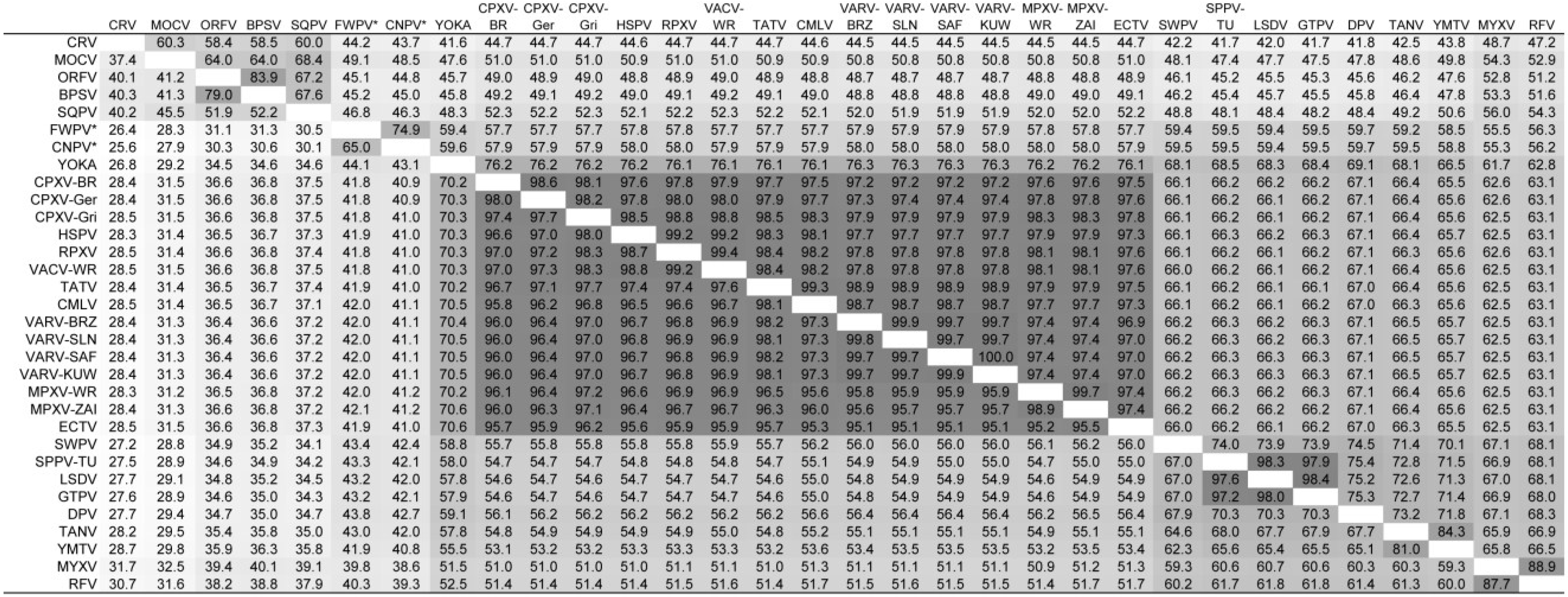
3.1. Assessment of Chordopoxvirus Gene Content

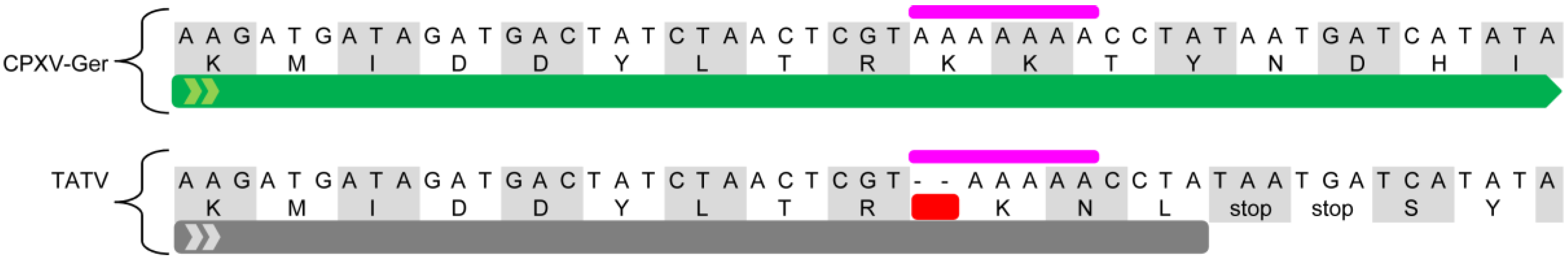
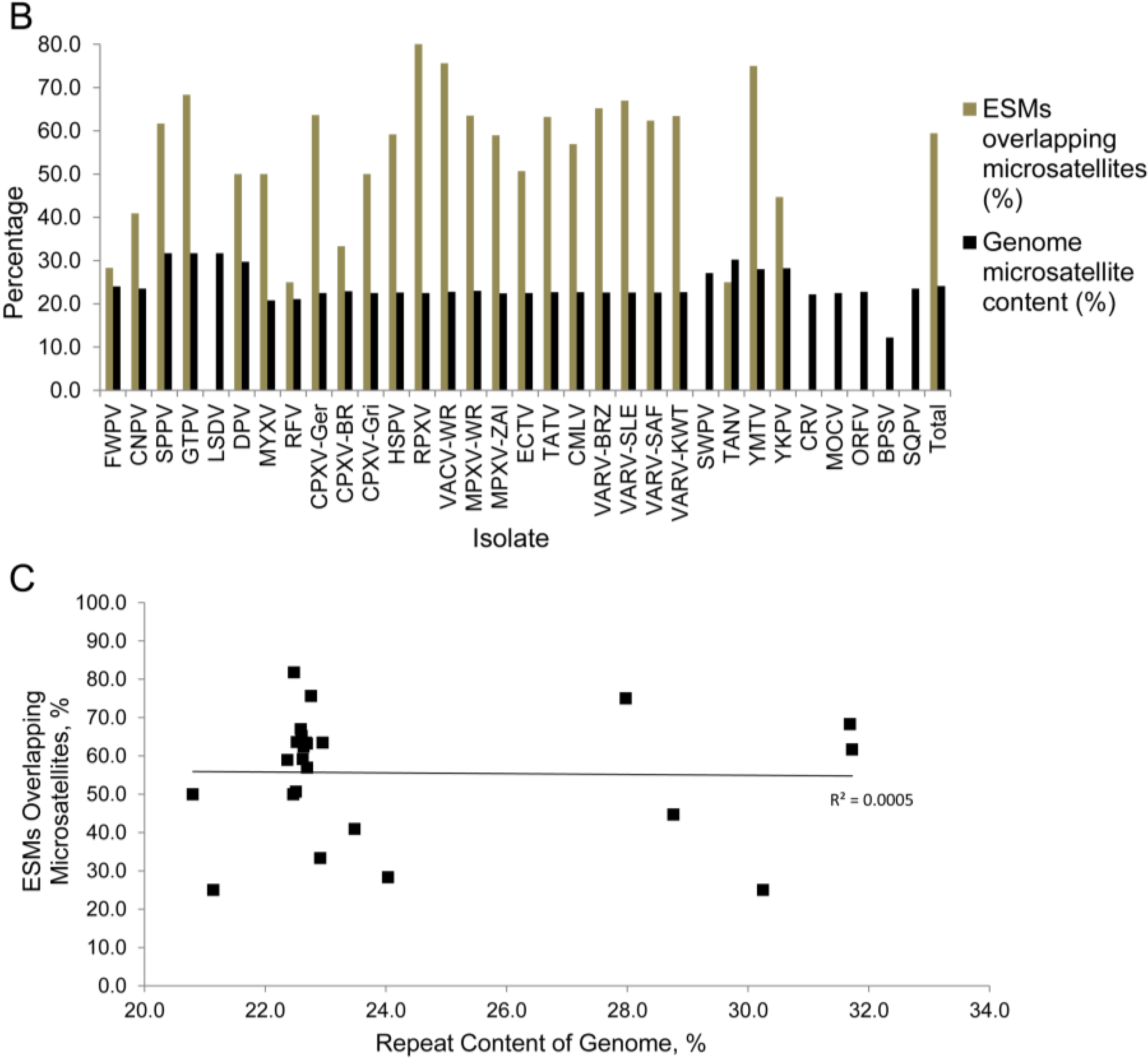
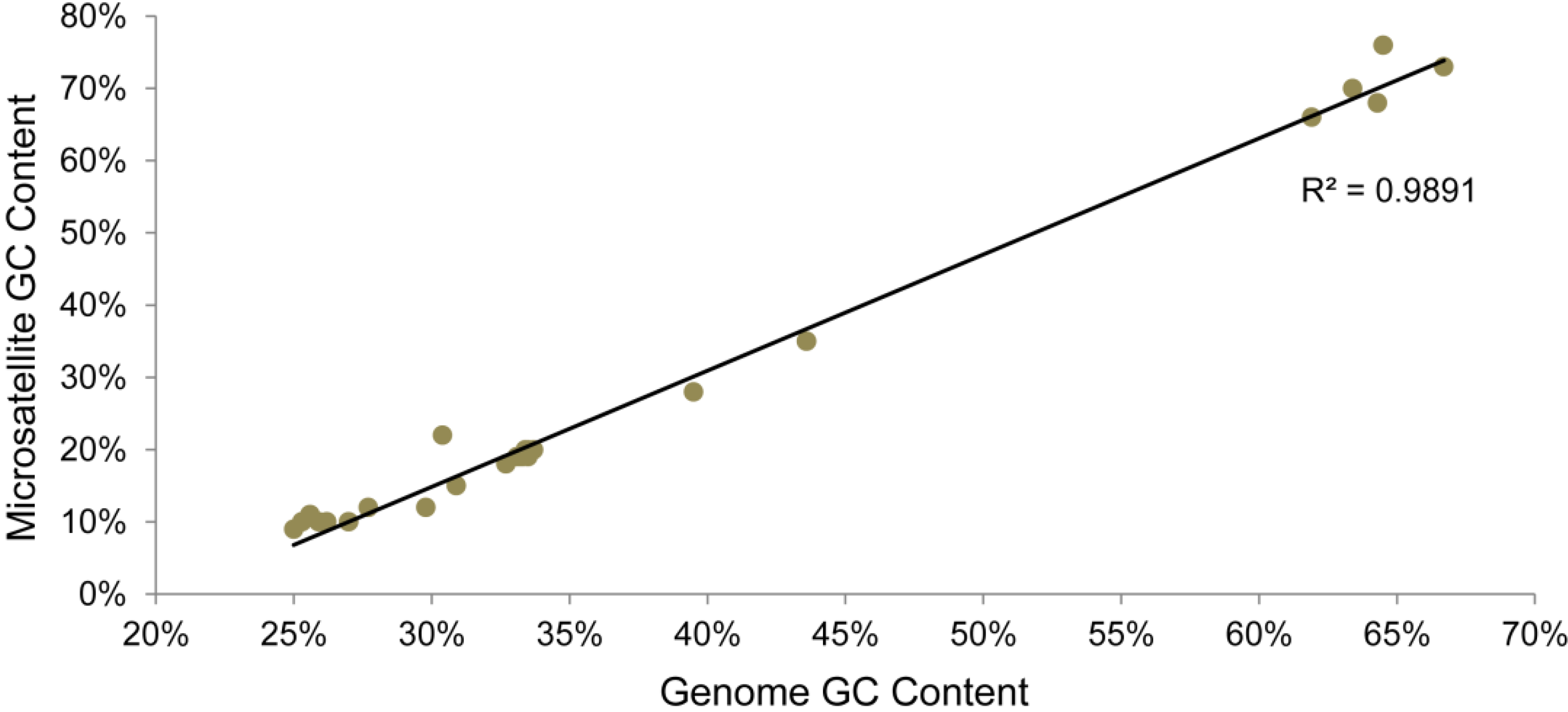
3.2. Assessment of Truncated and Fragmented Genes, and Early Stop Mutation Content
| Virus Genome | ESM Count | Non-intact genes | Non-intact genes/1000 bp | ESMs/non-intact gene | ESMs overlapping microsatellites | % of ESMs overlapping microsatellites | Genome microsatellite content (%) | GC% of genome | GC% of microsatellites |
|---|---|---|---|---|---|---|---|---|---|
| FWPV | 53 | 18 | 0.062 | 2.9 | 15 | 28.3 | 24.0 | 31 | 15 |
| CNPV | 22 | 6 | 0.017 | 3.7 | 9 | 40.9 | 23.5 | 30 | 22 |
| SPPV | 107 | 9 | 0.060 | 11.8 | 66 | 61.7 | 31.7 | 25 | 9 |
| GTPV | 82 | 9 | 0.060 | 9.1 | 56 | 68.3 | 31.7 | 25 | 10 |
| LSDV | 0 | 0 | 0 | - | 0 | - | 31.1 | 26 | 10 |
| DPV | 2 | 1 | 0.006 | 0.0 | 1 | 50.0 | 29.7 | 26 | 10 |
| CRV | 0 | 0 | 0 | - | 0 | - | 22.2 | 62 | 66 |
| MYXV | 2 | 1 | 0.006 | 1.0 | 1 | 50.0 | 20.8 | 44 | 35 |
| RFV | 44 | 4 | 0.025 | 11.0 | 11 | 25.0 | 21.1 | 40 | 28 |
| MOCV | 0 | 0 | 0 | - | 0 | - | 22.5 | 63 | 70 |
| CPXV-Ger | 11 | 6 | 0.026 | 1.8 | 7 | 63.6 | 22.5 | 34 | 20 |
| CPXV-BR | 3 | 4 | 0.018 | 0.8 | 1 | 33.3 | 22.9 | 33 | 20 |
| CPXV-Gri | 2 | 3 | 0.013 | 0.7 | 1 | 50.0 | 22.5 | 34 | 20 |
| HSPV | 49 | 26 | 0.122 | 1.9 | 29 | 59.2 | 22.6 | 33 | 19 |
| RPXV | 44 | 17 | 0.086 | 2.6 | 36 | 81.8 | 22.5 | 34 | 19 |
| VACV-WR | 41 | 15 | 0.077 | 2.7 | 31 | 75.6 | 22.8 | 33 | 19 |
| MPXV-WR | 104 | 21 | 0.105 | 5.0 | 66 | 63.5 | 22.7 | 33 | 19 |
| MPXV-ZAI | 95 | 21 | 0.107 | 4.5 | 56 | 58.9 | 22.6 | 33 | 19 |
| ECTV | 73 | 28 | 0.133 | 2.6 | 37 | 50.7 | 22.5 | 33 | 19 |
| TATV | 87 | 33 | 0.167 | 2.6 | 55 | 63.2 | 22.7 | 33 | 19 |
| CMLV | 65 | 22 | 0.107 | 3.0 | 37 | 56.9 | 22.7 | 33 | 19 |
| VARV-BRZ | 92 | 30 | 0.160 | 3.1 | 60 | 65.2 | 22.6 | 33 | 18 |
| VARV-SLE | 100 | 28 | 0.150 | 3.6 | 67 | 67.0 | 22.6 | 33 | 18 |
| VARV-SAF | 93 | 28 | 0.151 | 3.3 | 58 | 62.4 | 22.6 | 33 | 18 |
| VARV-KWT | 93 | 28 | 0.151 | 3.3 | 59 | 63.4 | 22.7 | 33 | 18 |
| ORFV | 0 | 0 | 0 | - | 0 | - | 22.8 | 64 | 68 |
| BPSV | 0 | 0 | 0 | - | 0 | - | 12.2 | 65 | 76 |
| SWPV | 0 | 0 | 0 | - | 0 | - | 27.1 | 28 | 12 |
| TANV | 4 | 2 | 0.014 | 1.0 | 1 | 25.0 | 30.2 | 27 | 10 |
| YMTV | 4 | 2 | 0.015 | 1.5 | 3 | 75.0 | 28.0 | 30 | 12 |
| YKPV | 47 | 17 | 0.097 | 2.8 | 21 | 44.7 | 28.8 * | 26 | 11 |
| SQPV | 0 | 0 | 0 | - | 0 | 0.0 | 23.5 | 67 | 73 |
| Total | 1140 | 379 | 0.060* | 3.4 * | 784 | 68.8 * | 24.1 | 37 * | 26 * |



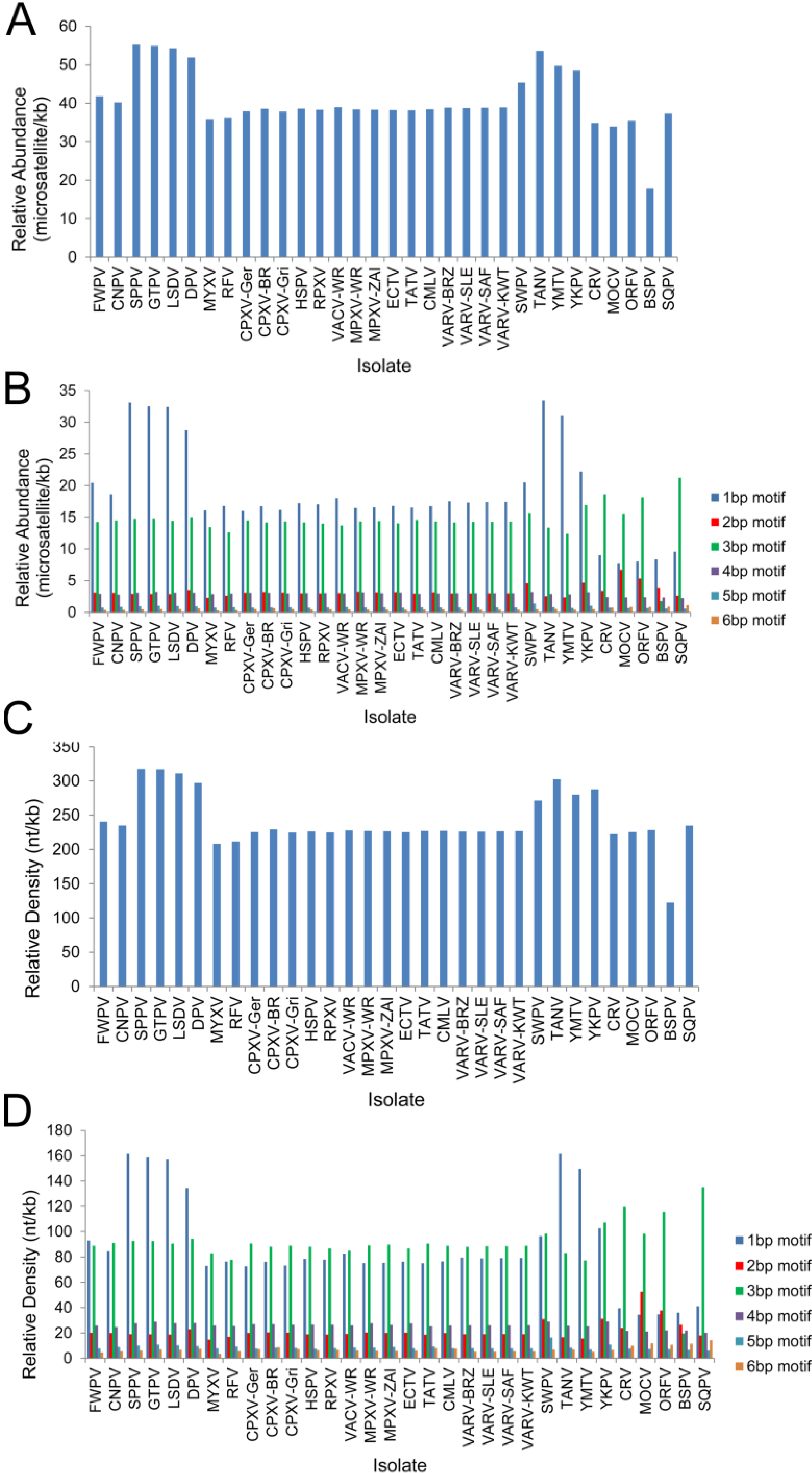
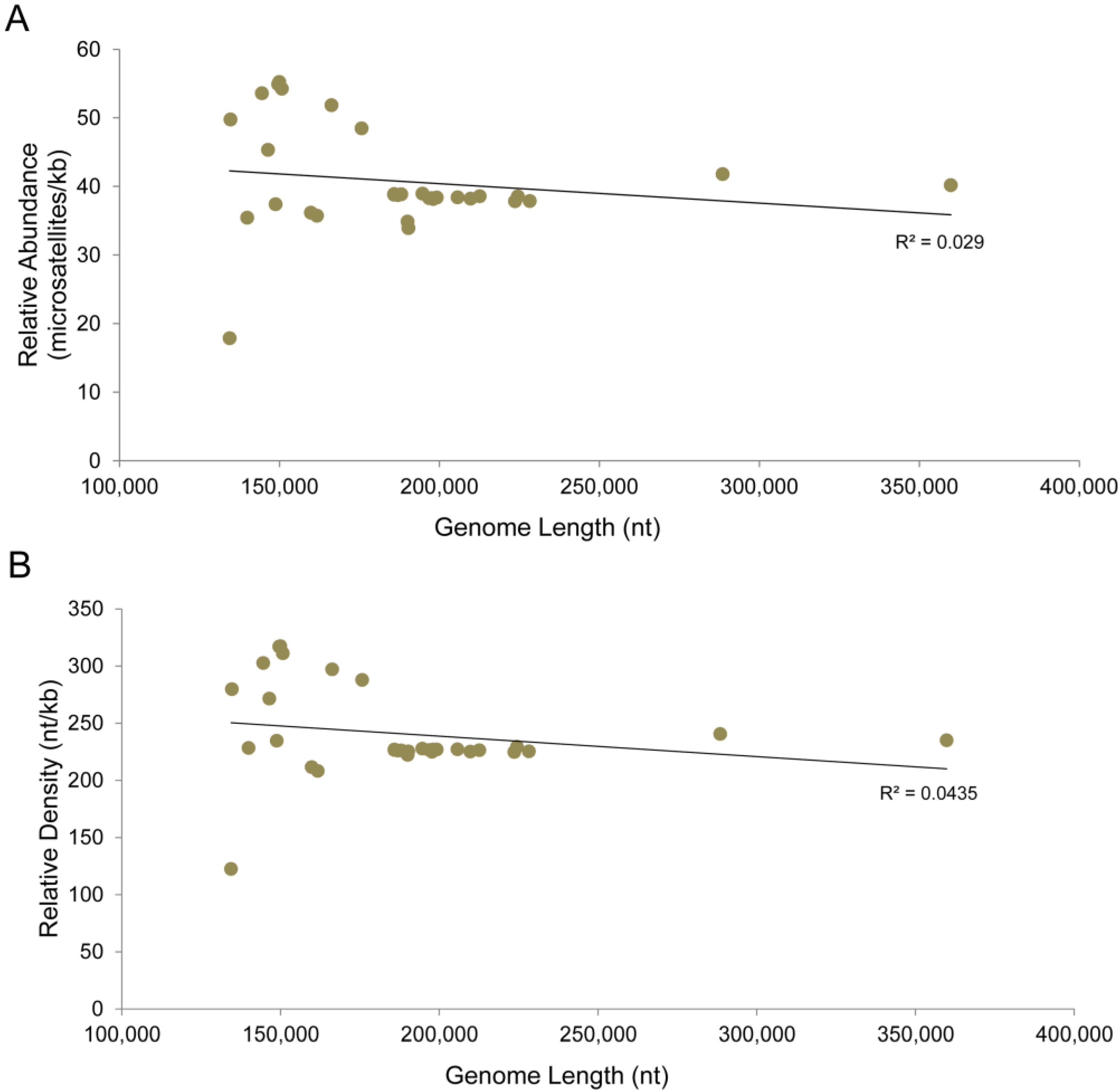
3.3. Chordopoxvirus Microsatellite Content
| Isolate | Total number | Total number of 1, 2, 3, 4, 5, 6 bp microsatellites | RA | RA of 1, 2, 3, 4, 5, 6 bp Microsatellites a | Total length of microsatellites | RD | RD of 1, 2, 3, 4, 5, 6 bp microsatellites | RA in noncoding regions | RA in coding regions |
|---|---|---|---|---|---|---|---|---|---|
| FWPV | 12063 | 5896, 894, 4110, 838, 217, 108 | 42 | 20, 3, 14, 3, 1, 0a | 69359 | 240 | 93, 20, 89, 26, 8, 5 | 49 | 40 |
| CNPV | 14465 | 6687, 1100, 5214, 999, 310, 147 | 40 | 19, 3, 14, 3, 1, 0a | 84508 | 235 | 85, 20, 91, 25, 9, 5 | 47 | 39 |
| SPPV | 8283 | 4964, 435, 2207, 461, 143, 73 | 55 | 33, 3, 15, 3, 1, 0a | 47574 | 317 | 162, 19, 93, 28, 10, 6 | 85 | 53 |
| GTPV | 8214 | 4863, 431, 2209, 481, 154, 76 | 55 | 33, 3, 15, 3, 1, 1 | 47406 | 317 | 159, 19, 92, 29, 11, 7 | 91 | 52 |
| LSDV | 8181 | 4888, 430, 2173, 463, 149, 78 | 54 | 32, 3, 14, 3, 1, 1 | 46886 | 311 | 157, 19, 91, 28, 10, 7 | 129 | 51 |
| DPV | 8620 | 4780, 583, 2490, 510, 154, 103 | 52 | 29, 4, 15, 3, 1, 1 | 49369 | 297 | 134, 23, 94, 28, 10, 8 | 81 | 49 |
| MYXV | 5780 | 2599, 373, 2174, 464, 123, 47 | 36 | 16, 2, 13, 3, 1, 0a | 33652 | 208 | 73, 15, 83, 26, 8, 4 | 88 | 33 |
| RFV | 5781 | 2683, 423, 2018, 469, 130, 58 | 36 | 17, 3, 13, 3, 1, 0a | 33800 | 211 | 76, 17, 78, 25, 9, 6 | 62 | 34 |
| CPXV-Ger | 8648 | 3650, 704, 3307, 698, 173, 116 | 38 | 16, 3, 14, 3, 1, 1 | 51415 | 225 | 73, 20, 91, 27, 8, 7 | 62 | 35 |
| CPXV-BR | 8656 | 3758, 716, 3178, 685, 172, 147 | 39 | 17, 3, 14, 3, 1, 1 | 51443 | 229 | 76, 20, 88, 27, 8, 9 | 57 | 36 |
| CPXV-Gri | 8466 | 3613, 691, 3202, 668, 178, 114 | 38 | 16, 3, 14, 3, 1, 1 | 50251 | 225 | 73, 20, 89, 27, 8, 7 | 53 | 36 |
| HSPV | 8205 | 3662, 632, 3012, 643, 158, 98 | 39 | 17, 3, 14, 3, 1, 0a | 48109 | 226 | 79, 19, 88, 27, 8, 6 | 45 | 37 |
| RPXV | 7570 | 3370, 584, 2766, 596, 154, 100 | 38 | 17, 3, 14, 3, 1, 1 | 44447 | 225 | 78, 19, 87, 27, 8, 7 | 44 | 37 |
| VACV-WR | 7586 | 3506, 588, 2664, 574, 162, 92 | 39 | 18, 3, 14, 3, 1, 0a | 44323 | 228 | 83, 19, 85, 26, 9, 6 | 45 | 37 |
| MPXV-WR | 7645 | 3281, 643, 2854, 626, 155, 86 | 38 | 16, 3, 14, 3, 1, 0a | 45187 | 227 | 75, 20, 89, 28, 8, 6 | 44 | 37 |
| MPXV-ZAI | 7540 | 3262, 617, 2830, 589, 157, 85 | 38 | 17, 3, 14, 3, 1, 0a | 44559 | 226 | 75, 20, 90, 26, 9, 6 | 46 | 37 |
| ECTV | 8018 | 3519, 667, 2942, 651, 153, 86 | 38 | 17, 3, 14, 3, 1, 0a | 47220 | 225 | 76, 20, 87, 28, 8, 6 | 45 | 36 |
| TATV | 7559 | 3273, 582, 2880, 575, 157, 92 | 38 | 17, 3, 15, 3, 1, 0a | 44943 | 227 | 75, 19, 91, 25, 9, 8 | 42 | 37 |
| CMLV | 7900 | 3446, 644, 2945, 607, 159, 99 | 38 | 17, 3, 14, 3, 1, 0a | 46688 | 227 | 77, 20, 89, 26, 8, 8 | 44 | 37 |
| VARV-BRZ | 7305 | 3294, 558, 2665, 564, 146, 78 | 39 | 18, 3, 14, 3, 1, 0a | 42513 | 226 | 80, 19, 88, 26, 8, 5 | 43 | 38 |
| VARV-SLE | 7241 | 3237, 554, 2667, 561, 145, 77 | 39 | 17, 3, 14, 3, 1, 0a | 42251 | 226 | 79, 19, 89, 26, 8, 5 | 43 | 37 |
| VARV-SAF | 7214 | 3234, 553, 2650, 556, 143, 78 | 39 | 17, 3, 14, 3, 1, 0a | 42078 | 226 | 79, 19, 89, 26, 8, 5 | 43 | 37 |
| VARV-KWT | 7228 | 3238, 553, 2658, 557, 144, 78 | 39 | 17, 3, 14, 3, 1, 0a | 42140 | 227 | 79, 19, 89, 26, 8, 5 | 43 | 38 |
| SWPV | 6642 | 3001, 672, 2296, 467, 205, 74 | 45 | 20, 5, 16, 3, 1, 1 | 39737 | 271 | 96, 31, 98, 29, 16, 7 | 68 | 44 |
| TANV | 7746 | 4835, 371, 1929, 415, 117, 79 | 54 | 33, 3, 13, 3, 1, 1 | 43730 | 302 | 162, 17, 83, 26, 8, 7 | 67 | 52 |
| YMTV | 6703 | 4184, 322, 1668, 379, 93, 57 | 50 | 31, 2, 12, 3, 1, 0a | 37689 | 280 | 150, 15, 77, 25, 7, 5 | 63 | 48 |
| YKPV | 8517 | 3902, 821, 2973, 554, 180, 87 | 48 | 22, 5, 17, 3, 1, 0a | 50540 | 288 | 103, 31, 107, 29, 11, 7 | 60 | 47 |
| CRV | 6626 | 1711, 640, 3533, 460, 139, 145 | 35 | 9, 3, 19, 2, 1, 1 | 42209 | 222 | 40, 24, 119, 22, 8, 10 | 62 | 34 |
| MOCV | 6456 | 1475, 1274, 2960, 454, 130, 163 | 34 | 8, 7, 16, 2, 1, 1 | 42852 | 225 | 34, 52, 98, 21, 7, 12 | 41 | 32 |
| ORFV | 4959 | 1123, 748, 2540, 335, 94, 119 | 35 | 8, 5, 18, 2, 1, 1 | 31940 | 228 | 35, 38, 116, 22, 7, 11 | 30 | 36 |
| BSPV | 2404 | 1123, 522, 240, 318, 80, 121 | 18 | 8, 4, 2, 2, 1, 1 | 16437 | 122 | 36, 27, 20, 22, 7, 11 | 34 | 17 |
| SQPV | 5565 | 1423, 393, 3159, 337, 85, 168 | 37 | 10, 3, 21, 2, 1, 1 | 34911 | 235 | 41, 18, 135, 20, 6, 14 | 56 | 36 |




4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- International Committee on Taxonomy of Viruses. Virus Taxonomy: Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier Academic Press: London, UK, 2011. [Google Scholar]
- Carroll, D.S.; Emerson, G.L.; Li, Y.; Sammons, S.; Olson, V.; Frace, M.; Nakazawa, Y.; Czerny, C.P.; Tryland, M.; Kolodziejek, J.; et al. Chasing jenner's vaccine: Revisiting cowpox virus classification. PLOS ONE 2011, 6, e23086. [Google Scholar]
- Zhao, G.; Droit, L.; Tesh, R.B.; Popov, V.L.; Little, N.S.; Upton, C.; Virgin, H.W.; Wang, D. The genome of yoka poxvirus. J. Virol. 2011, 85, 10230–10238. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, R.C.; Wang, C.; Hatcher, E.L.; Lefkowitz, E.J. Orthopoxvirus genome evolution: The role of gene loss. Viruses 2010, 2, 1933–1967. [Google Scholar] [CrossRef] [PubMed]
- Schramm, B.; Locker, J.K. Cytoplasmic organization of poxvirus DNA replication. Traffic 2005, 6, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Moss, B. Poxvirus DNA replication. Cold Spring Harb. Perspect. Biol. 2013, 5, a010199. [Google Scholar] [CrossRef]
- Lefkowitz, E.J.; Wang, C.; Upton, C. Poxviruses: Past, present and future. Virus Res. 2006, 117, 105–118. [Google Scholar] [CrossRef] [PubMed]
- McLysaght, A.; Baldi, P.F.; Gaut, B.S. Extensive gene gain associated with adaptive evolution of poxviruses. Proc. Natl. Acad. Sci. USA 2003, 100, 15655–15660. [Google Scholar] [CrossRef] [PubMed]
- Essbauer, S.; Pfeffer, M.; Meyer, H. Zoonotic poxviruses. Vet. Microbiol. 2010, 140, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.L. Genus Orthopoxvirus: Vaccinia virus. In Poxviruses; Mercer, A.A., Weber, O., Schmidt, A., Eds.; Birkhäuser Verlag: Basel, Switzerland, 2007; pp. 1–45. [Google Scholar]
- Emerson, G.L.; Li, Y.; Frace, M.A.; Olsen-Rasmussen, M.A.; Khristova, M.L.; Govil, D.; Sammons, S.A.; Regnery, R.L.; Karem, K.L.; Damon, I.K.; et al. The phylogenetics and ecology of the orthopoxviruses endemic to north america. PLOS ONE 2009, 4, e7666. [Google Scholar]
- Weli, S.C.; Tryland, M. Avipoxviruses: Infection biology and their use as vaccine vectors. Virol. J. 2011, 8, e49. [Google Scholar] [CrossRef]
- Mockett, B.; Binns, M.M.; Boursnell, M.E.; Skinner, M.A. Comparison of the locations of homologous fowlpox and vaccinia virus genes reveals major genome reorganization. J. Gen. Virol. 1992, 73, 2661–2668. [Google Scholar] [CrossRef] [PubMed]
- Tulman, E.R.; Afonso, C.L.; Lu, Z.; Zsak, L.; Kutish, G.F.; Rock, D.L. Genome of lumpy skin disease virus. J. Virol. 2001, 75, 7122–7130. [Google Scholar] [CrossRef] [PubMed]
- Afonso, C.L.; Tulman, E.R.; Lu, Z.; Zsak, L.; Osorio, F.A.; Balinsky, C.; Kutish, G.F.; Rock, D.L. The genome of swinepox virus. J. Virol. 2002, 76, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Bracht, A.J.; Armien, A.G.; Carrillo, C.; O'Hearn, E.S.; Fabian, A.W.; Moran, K.E.; Lu, Z.; Ariyakumer, D.S.; Rasmussen, J.M.; Metwally, S.A. Isolation and characterization of a cervidpoxvirus from a goitered gazelle (gazella subgutturosa) from a zoologic park in minnesota. J. Zoo Wildl. Med. 2013, 44, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Darby, A.C.; McInnes, C.J.; Kjaer, K.H.; Wood, A.R.; Hughes, M.; Martensen, P.M.; Radford, A.D.; Hall, N.; Chantrey, J. Novel host-related virulence factors are encoded by squirrelpox virus, the main causative agent of epidemic disease in red squirrels in the UK. PLOS ONE 2014, 9, e96439. [Google Scholar]
- MacNeil, A.; Lederman, E.; Reynolds, M.G.; Ragade, N.J.; Talken, R.; Friedman, D.; Hall, W.; Shwe, T.; Li, Y.; Zhao, H.; et al. Diagnosis of bovine-associated parapoxvirus infections in humans: Molecular and epidemiological evidence. Zoonoses Public Health 2010, 57, e161–e164. [Google Scholar]
- Afonso, C.L.; Delhon, G.; Tulman, E.R.; Lu, Z.; Zsak, A.; Becerra, V.M.; Zsak, L.; Kutish, G.F.; Rock, D.L. Genome of deerpox virus. J. Virol. 2005, 79, 966–977. [Google Scholar] [CrossRef] [PubMed]
- Hanson, D.; Diven, D.G. Molluscum contagiosum. Dermatol. Online J. 2003, 9, e2. [Google Scholar]
- Afonso, C.L.; Tulman, E.R.; Delhon, G.; Lu, Z.; Viljoen, G.J.; Wallace, D.B.; Kutish, G.F.; Rock, D.L. Genome of crocodilepox virus. J. Virol. 2006, 80, 4978–4991. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Carroll, D.S.; Gardner, S.N.; Walsh, M.C.; Vitalis, E.A.; Damon, I.K. On the origin of smallpox: Correlating variola phylogenics with historical smallpox records. Proc. Natl. Acad. Sci. USA 2007, 104, 15787–15792. [Google Scholar] [CrossRef] [PubMed]
- Likos, A.M.; Sammons, S.A.; Olson, V.A.; Frace, A.M.; Li, Y.; Olsen-Rasmussen, M.; Davidson, W.; Galloway, R.; Khristova, M.L.; Reynolds, M.G.; et al. A tale of two clades: Monkeypox viruses. J. Gen. Virol. 2005, 86, 2661–2672. [Google Scholar]
- Drake, J.W.; Hwang, C.B. On the mutation rate of herpes simplex virus type 1. Genetics 2005, 170, 969–970. [Google Scholar] [CrossRef] [PubMed]
- Duffy, S.; Shackelton, L.A.; Holmes, E.C. Rates of evolutionary change in viruses: Patterns and determinants. Nat. Rev. Genet. 2008, 9, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Odom, M.R.; Hendrickson, R.C.; Lefkowitz, E.J. Poxvirus protein evolution: Family wide assessment of possible horizontal gene transfer events. Virus Res. 2009, 144, 233–249. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, E.L.; Hendrickson, R.C.; Lefkowitz, E.J. Identification of nucleotide-level changes impacting gene content and genome evolution in orthopoxviruses. J. Virol. 2014, 88, 13651–13668. [Google Scholar] [CrossRef] [PubMed]
- Schlotterer, C. Evolutionary dynamics of microsatellite DNA. Chromosoma 2000, 109, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Behe, M.J. Tracts of adenosine and cytidine residues in the genomes of prokaryotes and eukaryotes. DNA Seq. 1998, 8, 375–383. [Google Scholar] [PubMed]
- Dieringer, D.; Schlotterer, C. Two distinct modes of microsatellite mutation processes: Evidence from the complete genomic sequences of nine species. Genome Res. 2003, 13, 2242–2251. [Google Scholar] [CrossRef] [PubMed]
- Coenye, T.; Vandamme, P. Characterization of mononucleotide repeats in sequenced prokaryotic genomes. DNA Res. 2005, 12, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Ellegren, H. Microsatellites: Simple sequences with complex evolution. Nat. Rev. Genet. 2004, 5, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Moxon, E.R.; Rainey, P.B.; Nowak, M.A.; Lenski, R.E. Adaptive evolution of highly mutable loci in pathogenic bacteria. Curr. Biol. 1994, 4, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Toth, G.; Gaspari, Z.; Jurka, J. Microsatellites in different eukaryotic genomes: Survey and analysis. Genome Res. 2000, 10, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Kaufer, B.B.; Jarosinski, K.W.; Osterrieder, N. Herpesvirus telomeric repeats facilitate genomic integration into host telomeres and mobilization of viral DNA during reactivation. J. Exp. Med. 2011, 208, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Baroudy, B.M.; Moss, B. Sequence homologies of diverse length tandem repetitions near ends of vaccinia virus genome suggest unequal crossing over. Nucleic Acids Res. 1982, 10, 5673–5679. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Mittal, N.; Sharma, P.C. Genome wide survey of microsatellites in ssdna viruses infecting vertebrates. Gene 2014, 552, 209–218. [Google Scholar] [CrossRef] [PubMed]
- George, B.; Mashhood Alam, C.; Jain, S.K.; Sharfuddin, C.; Chakraborty, S. Differential distribution and occurrence of simple sequence repeats in diverse geminivirus genomes. Virus Genes 2012, 45, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Tan, Z.; Jiang, J.; Li, M.; Chen, H.; Shen, G.; Yu, R. Similar distribution of simple sequence repeats in diverse completed human immunodeficiency virus type 1 genomes. FEBS Lett. 2009, 583, 2959–2963. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Tan, Z.; Zeng, G. Microsatellite is an important component of complete hepatitis c virus genomes. Infect. Genet. Evol. 2011, 11, 1646–1654. [Google Scholar] [CrossRef] [PubMed]
- Houng, H.S.; Lott, L.; Gong, H.; Kuschner, R.A.; Lynch, J.A.; Metzgar, D. Adenovirus microsatellite reveals dynamics of transmission during a recent epidemic of human adenovirus serotype 14 infection. J. Clin. Microbiol. 2009, 47, 2243–2248. [Google Scholar] [CrossRef] [PubMed]
- Mudunuri, S.B.; Nagarajaram, H.A. Imex: Imperfect microsatellite extractor. Bioinformatics 2007, 23, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Szpara, M.L.; Tafuri, Y.R.; Parsons, L.; Shamim, S.R.; Verstrepen, K.J.; Legendre, M.; Enquist, L.W. A wide extent of inter-strain diversity in virulent and vaccine strains of alphaherpesviruses. PLOS Pathog. 2011, 7, e1002282. [Google Scholar] [CrossRef]
- Viral Bioinformatics Resource Center. Available online: www.vbrc.org (accessed on 5 June 2008).
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. Mafft: A novel method for rapid multiple sequence alignment based on fast fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Bratke, K.A.; McLysaght, A. Identification of multiple independent horizontal gene transfers into poxviruses using a comparative genomics approach. BMC Evol. Biol. 2008, 8, e67. [Google Scholar] [CrossRef]
- Ouyang, Q.; Zhao, X.; Feng, H.; Tian, Y.; Li, D.; Li, M.; Tan, Z. High gc content of simple sequence repeats in herpes simplex virus type 1 genome. Gene 2012, 499, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Tulman, E.R.; Afonso, C.L.; Lu, Z.; Zsak, L.; Kutish, G.F.; Rock, D.L. The genome of canarypox virus. J. Virol. 2004, 78, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Elde, N.C.; Child, S.J.; Eickbush, M.T.; Kitzman, J.O.; Rogers, K.S.; Shendure, J.; Geballe, A.P.; Malik, H.S. Poxviruses deploy genomic accordions to adapt rapidly against host antiviral defenses. Cell 2012, 150, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.P.; Biagini, P.; Lefeuvre, P.; Golden, M.; Roumagnac, P.; Varsani, A. Recombination in eukaryotic single stranded DNA viruses. Viruses 2011, 3, 1699–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krupovic, M.; Koonin, E.V. Evolution of eukaryotic single-stranded DNA viruses of the bidnaviridae family from genes of four other groups of widely different viruses. Sci. Rep. 2014, 4, 5347. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatcher, E.L.; Wang, C.; Lefkowitz, E.J. Genome Variability and Gene Content in Chordopoxviruses: Dependence on Microsatellites. Viruses 2015, 7, 2126-2146. https://doi.org/10.3390/v7042126
Hatcher EL, Wang C, Lefkowitz EJ. Genome Variability and Gene Content in Chordopoxviruses: Dependence on Microsatellites. Viruses. 2015; 7(4):2126-2146. https://doi.org/10.3390/v7042126
Chicago/Turabian StyleHatcher, Eneida L., Chunlin Wang, and Elliot J. Lefkowitz. 2015. "Genome Variability and Gene Content in Chordopoxviruses: Dependence on Microsatellites" Viruses 7, no. 4: 2126-2146. https://doi.org/10.3390/v7042126
APA StyleHatcher, E. L., Wang, C., & Lefkowitz, E. J. (2015). Genome Variability and Gene Content in Chordopoxviruses: Dependence on Microsatellites. Viruses, 7(4), 2126-2146. https://doi.org/10.3390/v7042126