
Studies in a Murine Model Confirm the Safety of Griffithsin and Advocate Its Further Development as a Microbicide Targeting HIV-1 and Other Enveloped Viruses


Abstract
:1. Introduction
2. Materials and Methods
2.1. Griffithsin (GRFT) and Other Lectin Reagents
2.2. In Vitro Effects of GRFT in Mouse Peripheral Blood Mononuclear Cell (mPBMCs)
2.2.1. Mouse PBMC Isolation and Culture
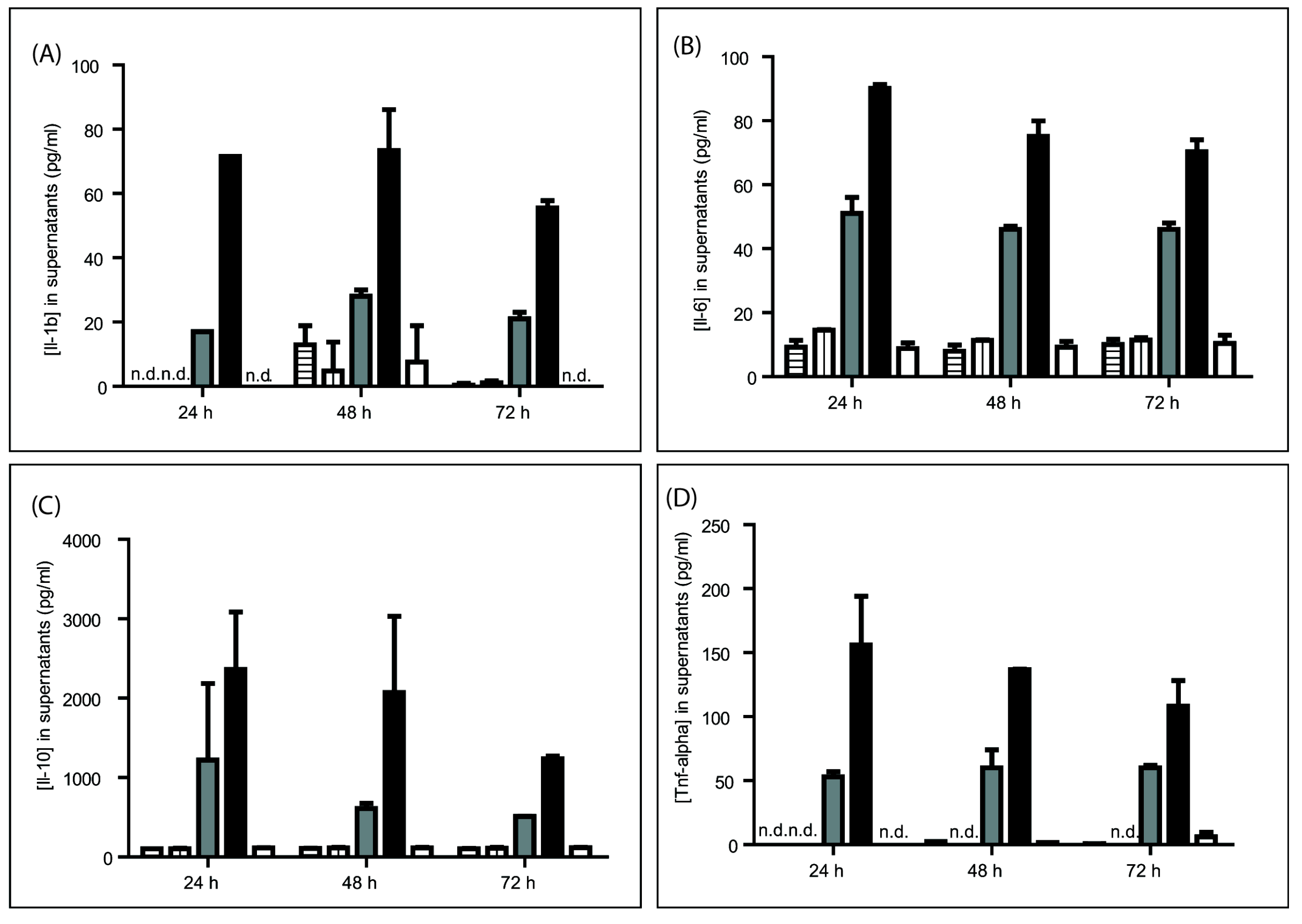
2.2.2. Cytokine Release
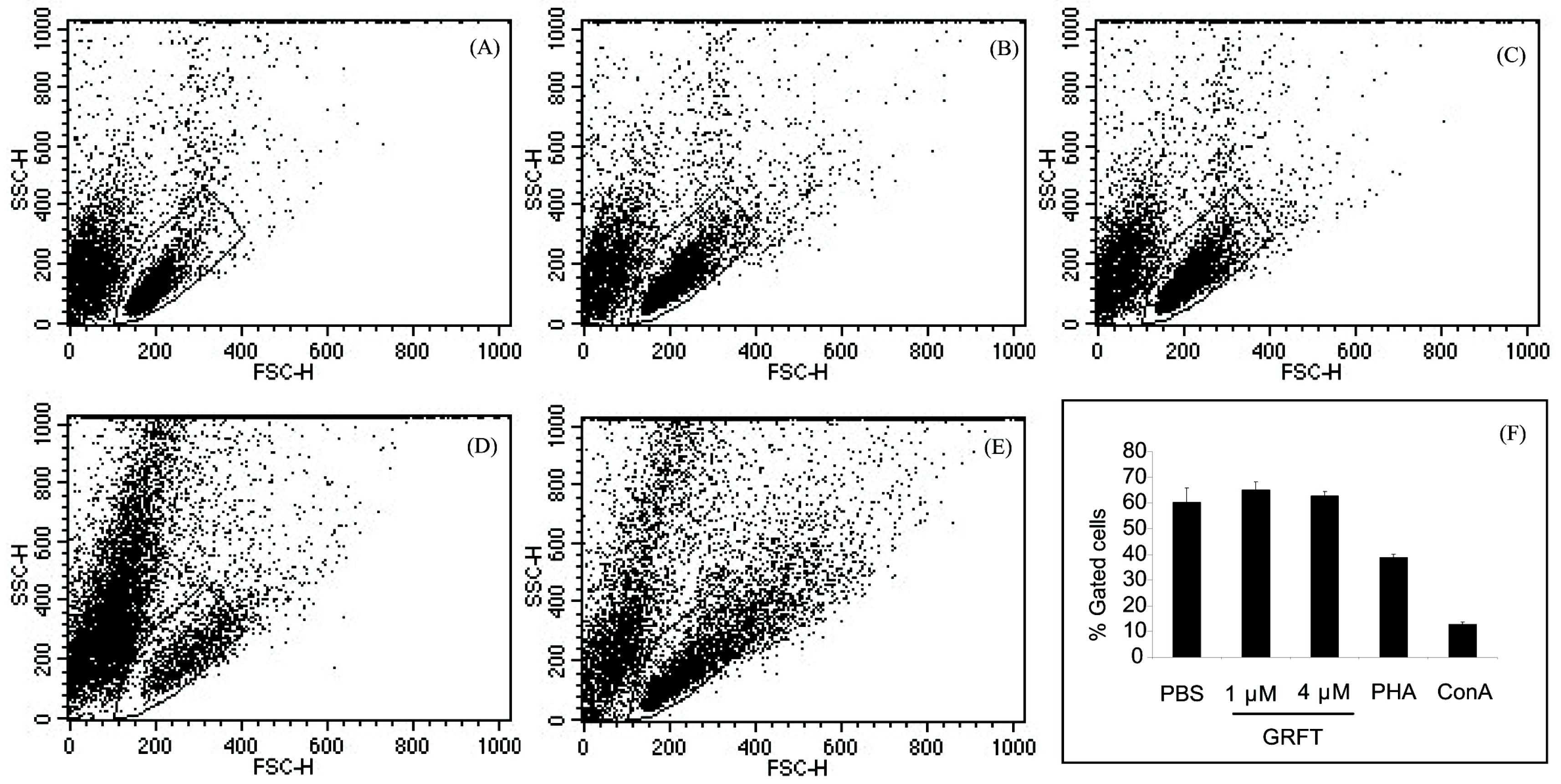
2.2.3. Cell Viability and Mitogenicity
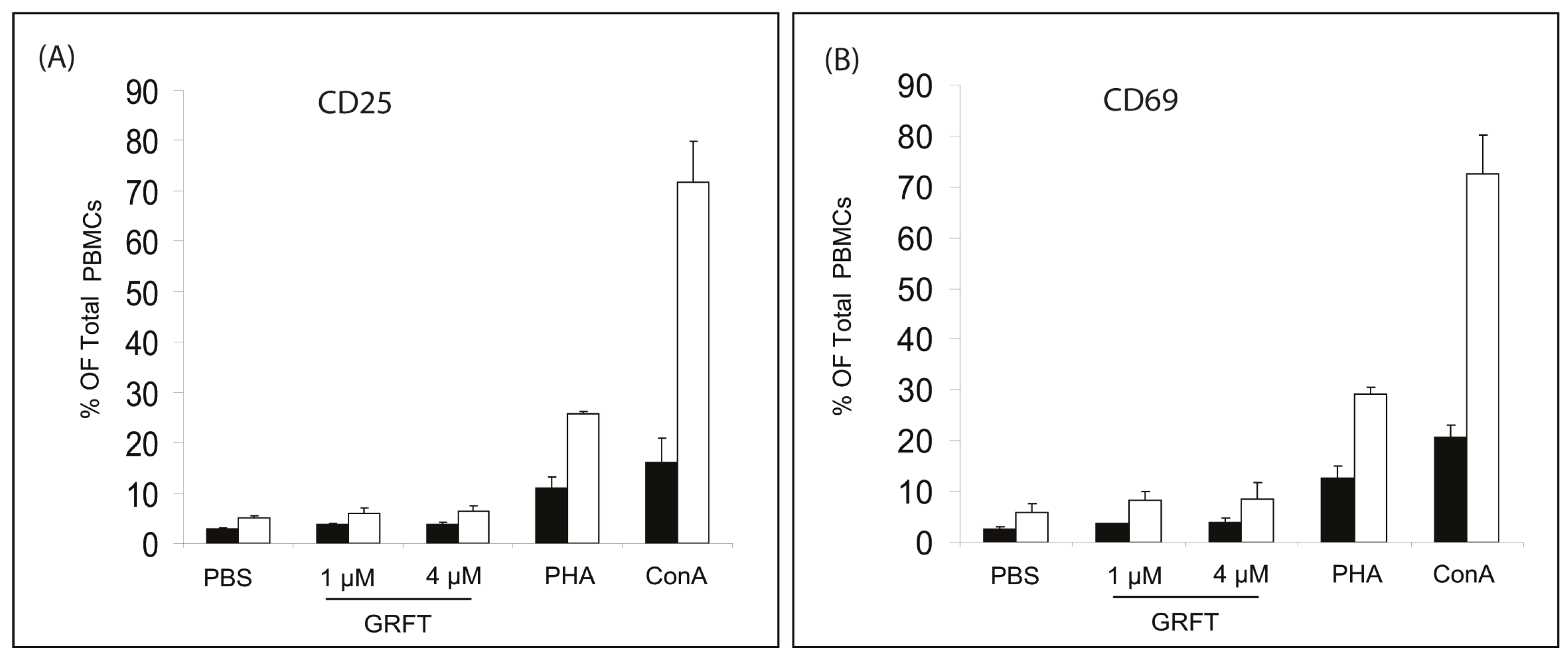
2.2.4. Surface Activation Marker Levels
2.3. In Vivo Effects of GRFT in a Mouse Model
2.3.1. Animal Housing
2.3.2. Subcutaneous (s.c.) Injections and Sample Collection
2.3.3. Intravaginal and Intraperitoneal Administrations, and Sample Collection
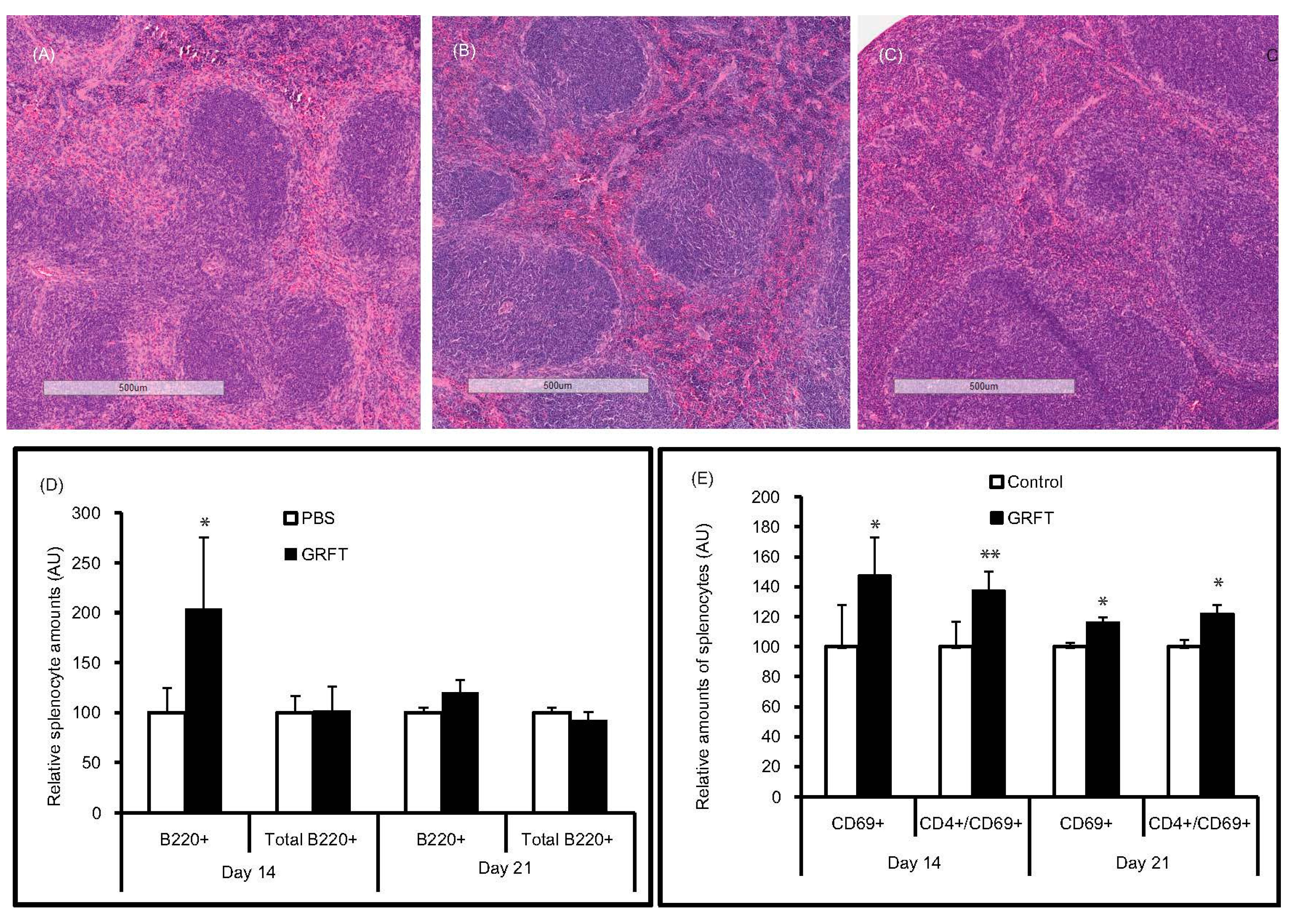
2.3.4. Splenocyte Activation
2.3.5. Histopathology
2.3.6. Hematology Parameters
2.3.7. Blood Chemistry
2.4. Statistical Analysis
3. Results
3.1. In Vitro Effects of GRFT on Purified mPBMCs
3.2. GRFT Toxicity in Mice after Parenteral Administration of a Single Dose
3.3. GRFT Toxicity in Mice after Administration of a Chronic Dose of 10 mg/kg
3.4. GRFT Toxicity in Mice after Administration of a Chronic Dose of 2 mg/kg
4. Discussion
4.1. GRFT’S Effects on mPBMCs Validate the Use of a Murine Model for Safety Studies
4.2. Even at Very High Doses, GRFT Does Not Cause Overt Toxicity in Mice
4.3. GRFT Does Not Significantly Affect Blood Chemistry Markers of Toxicity
4.4. Chronic Intravaginal or Systemic Administration of 2 mg/kg GRFT Is Safe in Mice
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- The Joint United Nations Programe on HIV/AIDS (UNAIDS). How AIDS Changed Everything—MDG6: 15 Years, 15 Lessons of Hope from the AIDS Response; UNAIDS: Geneva, Switzerland, 2015. [Google Scholar]
- Kouokam, J.C.; Palmer, K.E. Plant-expressed griffithsin: A protein with potent broad spectrum inhibitory effects against enveloped viruses. In Medicinal Plant Biotechnology; Arora, R., Ed.; CAB International: Oxfordshire, UK, 2010; pp. 334–345. [Google Scholar]
- Mori, T.; O’Keefe, B.R.; Sowder, R.C., 2nd; Bringans, S.; Gardella, R.; Berg, S.; Cochran, P.; Turpin, J.A.; Buckheit, R.W., Jr.; McMahon, J.B.; et al. Isolation and characterization of griffithsin, a novel HIV-inactivating protein, from the red alga Griffithsia sp. J. Biol. Chem. 2005, 280, 9345–9353. [Google Scholar] [CrossRef] [PubMed]
- Emau, P.; Tian, B.; O’Keefe, B.R.; Mori, T.; McMahon, J.B.; Palmer, K.E.; Jiang, Y.; Bekele, G.; Tsai, C.C. Griffithsin, a potent HIV entry inhibitor, is an excellent candidate for anti-HIV microbicide. J. Med. Primatol. 2007, 36, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Kagiampakis, I.; Gharibi, A.; Mankowski, M.K.; Snyder, B.A.; Ptak, R.G.; Alatas, K.; LiWang, P.J. Potent strategy to inhibit HIV-1 by binding both gp120 and gp41. Antimicrob. Agents Chemother. 2011, 55, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Ziolkowska, N.E.; Shenoy, S.R.; O’Keefe, B.R.; Wlodawer, A. Crystallographic studies of the complexes of antiviral protein griffithsin with glucose and N-acetylglucosamine. Protein Sci. 2007, 16, 1485–1489. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, B.R.; Giomarelli, B.; Barnard, D.L.; Shenoy, S.R.; Chan, P.K.; McMahon, J.B.; Palmer, K.E.; Barnett, B.W.; Meyerholz, D.K.; Wohlford-Lenane, C.L.; et al. Broad-Spectrum In Vitro Activity and In Vivo Efficacy of the Antiviral Protein Griffithsin against Emerging Viruses of the Family Coronaviridae. J. Virol. 2010, 84, 2511–2521. [Google Scholar] [CrossRef] [PubMed]
- Ishag, H.Z.; Li, C.; Huang, L.; Sun, M.X.; Wang, F.; Ni, B.; Malik, T.; Chen, P.Y.; Mao, X. Griffithsin inhibits Japanese encephalitis virus infection in vitro and in vivo. Arch. Virol. 2013, 158, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Nixon, B.; Stefanidou, M.; Mesquita, P.M.; Fakioglu, E.; Segarra, T.; Rohan, L.; Halford, W.; Palmer, K.E.; Herold, B.C. Griffithsin protects mice from genital herpes by preventing cell-to-cell spread. J. Virol. 2013, 87, 6257–6269. [Google Scholar] [CrossRef] [PubMed]
- Meuleman, P.; Albecka, A.; Belouzard, S.; Vercauteren, K.; Verhoye, L.; Wychowski, C.; Leroux-Roels, G.; Palmer, K.E.; Dubuisson, J. Griffithsin has antiviral activity against hepatitis C virus. Antimicrob. Agents Chemother. 2011, 55, 5159–5167. [Google Scholar] [CrossRef] [PubMed]
- Takebe, Y.; Saucedo, C.J.; Lund, G.; Uenishi, R.; Hase, S.; Tsuchiura, T.; Kneteman, N.; Ramessar, K.; Tyrrell, D.L.; Shirakura, M.; et al. Antiviral lectins from red and blue-green algae show potent in vitro and in vivo activity against hepatitis C virus. PLoS ONE 2013, 8, e64449. [Google Scholar] [CrossRef] [PubMed]
- Kouokam, J.C.; Huskens, D.; Schols, D.; Johannemann, A.; Riedell, S.K.; Walter, W.; Walker, J.M.; Matoba, N.; O’Keefe, B.R.; Palmer, K.E. Investigation of griffithsin’s interactions with human cells confirms its outstanding safety and efficacy profile as a microbicide candidate. PLoS ONE 2011, 6, e22635. [Google Scholar] [CrossRef] [PubMed]
- Mauck, C.K.; Lai, J.J.; Weiner, D.H.; Chandra, N.; Fichorova, R.N.; Dezzutti, C.S.; Hillier, S.L.; Archer, D.F.; Creinin, M.D.; Schwartz, J.L.; et al. Toward Early Safety Alert Endpoints: Exploring Biomarkers Suggestive of Microbicide Failure. AIDS Res. Hum. Retrovir. 2013, 29, 1475–1486. [Google Scholar] [CrossRef] [PubMed]
- Friend, D.R.; Kiser, P.F. Assessment of topical microbicides to prevent HIV-1 transmission: Concepts, testing, lessons learned. Antivir. Res. 2013, 99, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, L.; Ramjee, G.; Alary, M.; Vuylsteke, B.; Chandeying, V.; Rees, H.; Sirivongrangson, P.; Mukenge-Tshibaka, L.; Ettiegne-Traore, V.; Uaheowitchai, C.; et al. Effectiveness of COL-1492, a nonoxynol-9 vaginal gel, on HIV-1 transmission in female sex workers: A randomised controlled trial. Lancet 2002, 360, 971–977. [Google Scholar] [CrossRef]
- O’Keefe, B.R.; Vojdani, F.; Buffa, V.; Shattock, R.J.; Montefiori, D.C.; Bakke, J.; Mirsalis, J.; d’Andrea, A.L.; Hume, S.D.; Bratcher, B.; et al. Scaleable manufacture of HIV-1 entry inhibitor griffithsin and validation of its safety and efficacy as a topical microbicide component. Proc. Natl. Acad. Sci. USA 2009, 106, 6099–6104. [Google Scholar] [CrossRef] [PubMed]
- Barton, C.; Kouokam, J.C.; Lasnik, A.B.; Foreman, O.; Cambon, A.; Brock, G.; Montefiori, D.C.; Vojdani, F.; McCormick, A.A.; O’Keefe, B.R.; et al. Activity and effect of subcutaneous treatment with the broad spectrum antiviral lectin Griffithsin in two laboratory rodent models. Antimicrob. Agents Chemother. 2014, 58, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.W.; Svenson, R.H.; Yachnin, S. Purification of mitogenic proteins derived from Phaseolus vulgaris: Isolation of potent and weak phytohemagglutinins possessing mitogenic activity. Proc. Natl. Acad. Sci. USA 1969, 63, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Kruisbeek, A.M. Isolation of mouse mononuclear cells. Curr. Protoc. Immunol. 2001. [Google Scholar] [CrossRef]
- Barrientos, L.G.; O’Keefe, B.R.; Bray, M.; Sanchez, A.; Gronenborn, A.M.; Boyd, M.R. Cyanovirin-N binds to the viral surface glycoprotein, GP1,2 and inhibits infectivity of Ebola virus. Antivir. Res. 2003, 58, 47–56. [Google Scholar] [CrossRef]
- Linderoth, A.; Prykhod’ko, O.; Pierzynowski, S.G.; Westrom, B.R. Enterally but not parenterally administered Phaseolus vulgaris lectin induces growth and precocious maturation of the gut in suckling rats. Biol. Neonate 2006, 89, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Chen, X.; Hase, K.; Sakamoto, A.; Nishigaki, C.; Ohno, H. Splenic CD19−CD35+B220+ cells function as an inducer of follicular dendritic cell network formation. Blood 2007, 110, 1215–1224. [Google Scholar] [CrossRef] [PubMed]
- Kosco, M.H.; Pflugfelder, E.; Gray, D. Follicular dendritic cell-dependent adhesion and proliferation of B cells in vitro. J. Immunol. 1992, 148, 2331–2339. [Google Scholar] [PubMed]
- Bear, H.D.; McFadden, A.W.; Kostuchenko, P.J.; Lipshy, K.A.; Hamad, G.G.; Turner, A.J.; Roberts, J.D.; Carr, M.; Carr, S.; Grant, S. Bryostatin 1 activates splenic lymphocytes and induces sustained depletion of splenocyte protein kinase C activity in vivo after a single intravenous administration. Anti-Cancer Drugs 1996, 7, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Esplugues, E.; Sancho, D.; Vega-Ramos, J.; Martinez, C.; Syrbe, U.; Hamann, A.; Engel, P.; Sanchez-Madrid, F.; Lauzurica, P. Enhanced antitumor immunity in mice deficient in CD69. J. Exp. Med. 2003, 197, 1093–1106. [Google Scholar] [CrossRef] [PubMed]
- Sancho, D.; Gomez, M.; Viedma, F.; Esplugues, E.; Gordon-Alonso, M.; Garcia-Lopez, M.A.; de la Fuente, H.; Martinez, A.C.; Lauzurica, P.; Sanchez-Madrid, F. CD69 downregulates autoimmune reactivity through active transforming growth factor-beta production in collagen-induced arthritis. J. Clin. Investig. 2003, 112, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Naghashpour, M.; Cualing, H. Splenomegaly with sea-blue histiocytosis, dyslipidemia, and nephropathy in a patient with lecithin-cholesterol acyltransferase deficiency: A clinicopathologic correlation. Metabolism 2009, 58, 1459–1464. [Google Scholar] [CrossRef] [PubMed]
- Yong, J.M. Origins of serum alkaline phosphatase. J. Clin. Pathol. 1967, 20, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Fleming, D.T.; Wasserheit, J.N. From epidemiological synergy to public health policy and practice: The contribution of other sexually transmitted diseases to sexual transmission of HIV infection. Sex. Transm. Infect. 1999, 75, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Barnabas, R.V.; Wasserheit, J.N. Riddle of the Sphinx revisited: The role of STDs in HIV prevention. Sex. Transm. Dis. 2009, 36, 365–367. [Google Scholar] [CrossRef] [PubMed]
- Wasserheit, J.N. Epidemiological synergy. Interrelationships between human immunodeficiency virus infection and other sexually transmitted diseases. Sex. Transm. Dis. 1992, 19, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Levendosky, K.; Mizenina, O.; Martinelli, E.; Jean-Pierre, N.; Kizima, L.; Rodriguez, A.; Kleinbeck, K.; Bonnaire, T.; Robbiani, M.; Zydowsky, T.M.; et al. Griffithsin and Carrageenan Combination to Target Herpes Simplex Virus 2 and Human Papillomavirus. Antimicrob. Agents Chemother. 2015, 59, 7290–7298. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | Day 14 | Day 21 | ||
|---|---|---|---|---|---|
| PBS | GRFT | PBS | GRFT | ||
| Alb | g/dL | 3.0 ± 0.4 | 3.0 ± 0.3 | 2.2 ± 0.4 | 2.9 ± 0.1 |
| ALP | AU | 100.0 ± 16.7 | 159.9 ± 31.0 1 | 100.0 ± 14.0 | 144.5 ± 18.0 1 |
| ALT | AU | 100.0 ± 36.7 | 86.5 ± 7.0 | 100.0 ± 73.0 | 57.1 ± 35.5 |
| AMS | AU | 100.0 ± 32.8 | 95.6 ± 7.5 | 100.0 ± 33.3 | 69.8 ± 6.2 |
| BG | mg/dL | 119.8 ± 24.7 | 124.8 ± 18.6 | 166.3 ± 28.3 | 138.0 ± 42.8 |
| BUN | mg/dL | 29.3 ± 3.3 | 25.0 ± 3.2 | 33.3 ± 6.7 | 26.0 ± 1.7 |
| Ca | mg/dL | 10.1 ± 0.4 | 10.9 ± 0.3 | 11.1 ± 1.2 | 10.6 ± 0.3 |
| Chol | mg/dL | 87.3 ± 5.4 | 118.8 ± 10.5 1 | 69.0 ± 14.2 | 103.0 ± 3.6 1 |
| CREAT | mg/dL | 0.4 ± 0.1 | 0.4 ± 0.0 | 0.2 ± 0.1 | 0.3 ± 0.1 |
| GLOB | g/dL | 3.1 ± 0.2 | 3.3 ± 0.1 | 4.1 ± 1.0 | 3.1 ± 0.1 |
| P | mg/dL | 10.4 ± 1.8 | 10.9 ± 0.9 | 10.3 ± 0.5 | 9.5 ± 0.9 |
| TBILI | mg/dL | 1.2 ± 0.5 | 1.2 ± 0.6 | 1.0 ± 0.5 | 0.5 ± 0.2 |
| TP | g/dL | 6.1 ± 0.6 | 6.3 ± 0.4 | 6.3 ± 0.6 | 5.9 ± 0.1 |
| Cell Type | Parameter | Unit | Day 14 | Day 21 | ||
|---|---|---|---|---|---|---|
| PBS | GRFT | PBS | GRFT | |||
| Leucocytes | WBC | k/μL | 7.9 ± 1.5 | 7.2 ± 0.5 | 8.3 ± 2.4 | 8.5 ± 1.6 |
| NE | k/μL | 1.7 ± 0.6 | 2.7 ± 0.2 | 2.1 ± 1.3 | 2.9 ± 1.0 | |
| LY | k/μL | 5.4 ± 0.7 | 3.6 ± 0.4 1 | 5.0 ± 0.6 | 4.3 ± 0.4 | |
| MO | k/μL | 0.6 ± 0.1 | 0.6 ± 0.1 | 0.9 ± 0.2 | 1.0 ± 0.5 | |
| EO | k/μL | 0.2 ± 0.1 | 0.3 ± 0.1 | 0.2 ± 0.3 | 0.3 ± 0.1 | |
| BA | k/μL | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.1 | 0.1 ± 0.0 | |
| Erythrocytes | RBC | M/µL | 9.4 ± 0.5 | 9.1 ± 0.7 | 9.8 ± 1.1 | 10.1 ± 2.3 |
| Hb | g/dL | 14.2 ± 1.0 | 13.2 ± 1.1 | 15.0 ± 0.8 | 15.1 ± 4.0 | |
| HCT | % | 57.9 ± 5.3 | 56.2 ± 5.5 | 59.4 ± 6.2 | 60.1 ± 14.9 | |
| MCV | fL | 61.4 ± 2.6 | 61.7 ± 1.2 | 60.6 ± 3.4 | 59.3 ± 1.5 | |
| MCH | pg | 15.1 ± 0.4 | 14.5 ± 0.6 | 15.3 ± 0.9 | 14.8 ± 0.7 | |
| MCHC | g/dL | 24.5 ± 0.6 | 23.5 ± 1.0 | 25.3 ± 1.7 | 25.0 ± 0.6 | |
| RDW | % | 17.3 ± 0.9 | 18.2 ± 0.5 | 18.0 ± 0.4 | 18.5 ± 0.4 | |
| Thrombocytes | PLT | k/μL | 1571.3 ± 100.8 | 1397.0 ± 302.5 | 850.0 ± 43.5 | 949.0 ± 211.5 |
| MPV | fL | 3.6 ± 0.1 | 3.7 ± 0.3 | 3.6 ± 0.2 | 3.8 ± 0.2 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kouokam, J.C.; Lasnik, A.B.; Palmer, K.E. Studies in a Murine Model Confirm the Safety of Griffithsin and Advocate Its Further Development as a Microbicide Targeting HIV-1 and Other Enveloped Viruses. Viruses 2016, 8, 311. https://doi.org/10.3390/v8110311
Kouokam JC, Lasnik AB, Palmer KE. Studies in a Murine Model Confirm the Safety of Griffithsin and Advocate Its Further Development as a Microbicide Targeting HIV-1 and Other Enveloped Viruses. Viruses. 2016; 8(11):311. https://doi.org/10.3390/v8110311
Chicago/Turabian StyleKouokam, Joseph Calvin, Amanda B. Lasnik, and Kenneth E. Palmer. 2016. "Studies in a Murine Model Confirm the Safety of Griffithsin and Advocate Its Further Development as a Microbicide Targeting HIV-1 and Other Enveloped Viruses" Viruses 8, no. 11: 311. https://doi.org/10.3390/v8110311
APA StyleKouokam, J. C., Lasnik, A. B., & Palmer, K. E. (2016). Studies in a Murine Model Confirm the Safety of Griffithsin and Advocate Its Further Development as a Microbicide Targeting HIV-1 and Other Enveloped Viruses. Viruses, 8(11), 311. https://doi.org/10.3390/v8110311