
Aphis Glycines Virus 2, a Novel Insect Virus with a Unique Genome Structure


Abstract
:1. Introduction
2. Materials and Methods
2.1. Aphid Rearing
2.2. ApGlV2 Genome Assembly
2.3. Establishment of Single Soybean Aphid Families
2.4. ApGlV2 Virion Purification
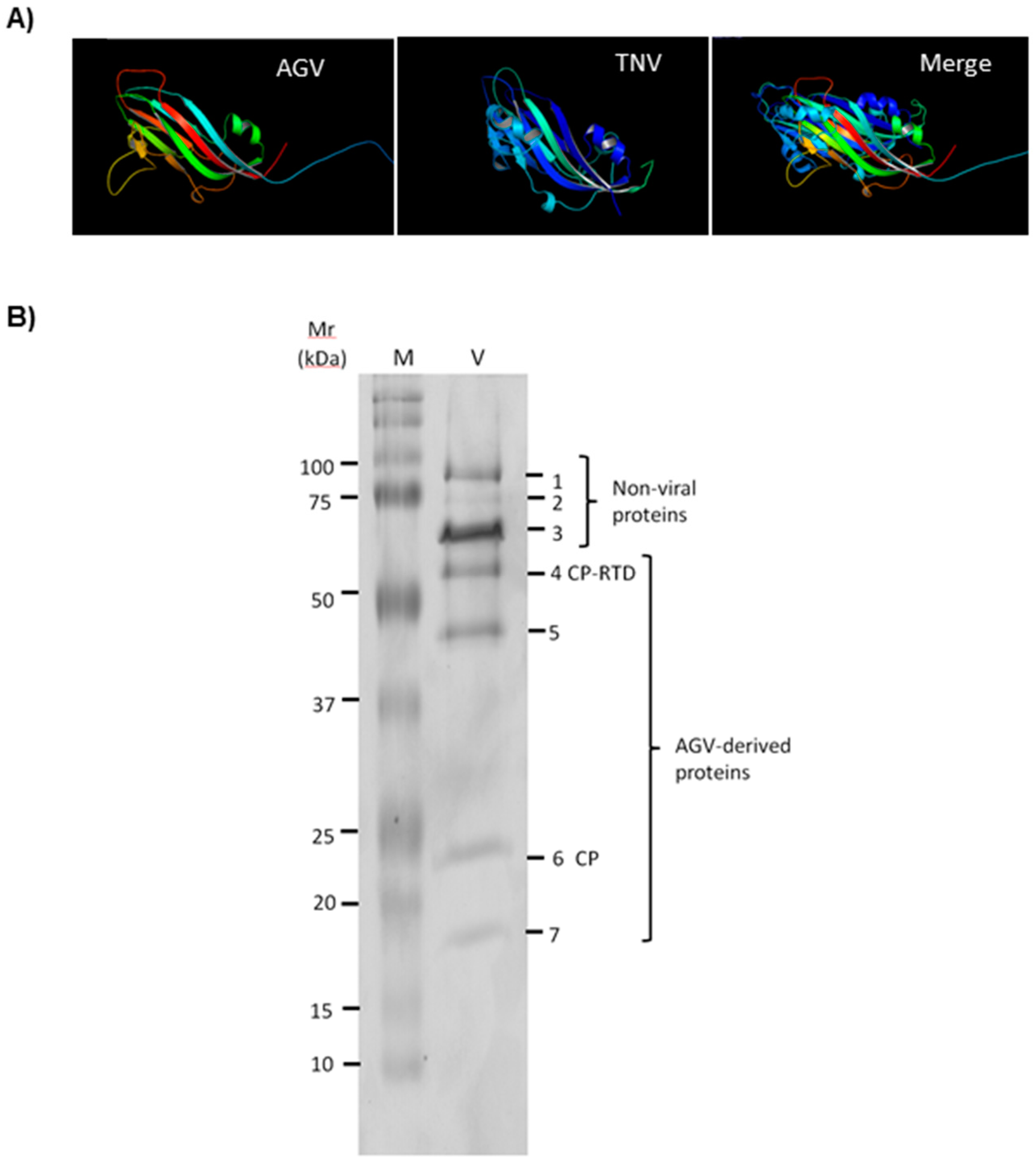
2.5. Characterization of ApGlV2 CP and CP-RTD Proteins
2.6. Transmission Electron Microscopy
2.7. RNA and DNA Purification from Aphids
2.8. PCR, RT-PCR, and RACE-PCR
2.9. Bioinformatics Analysis
3. Results
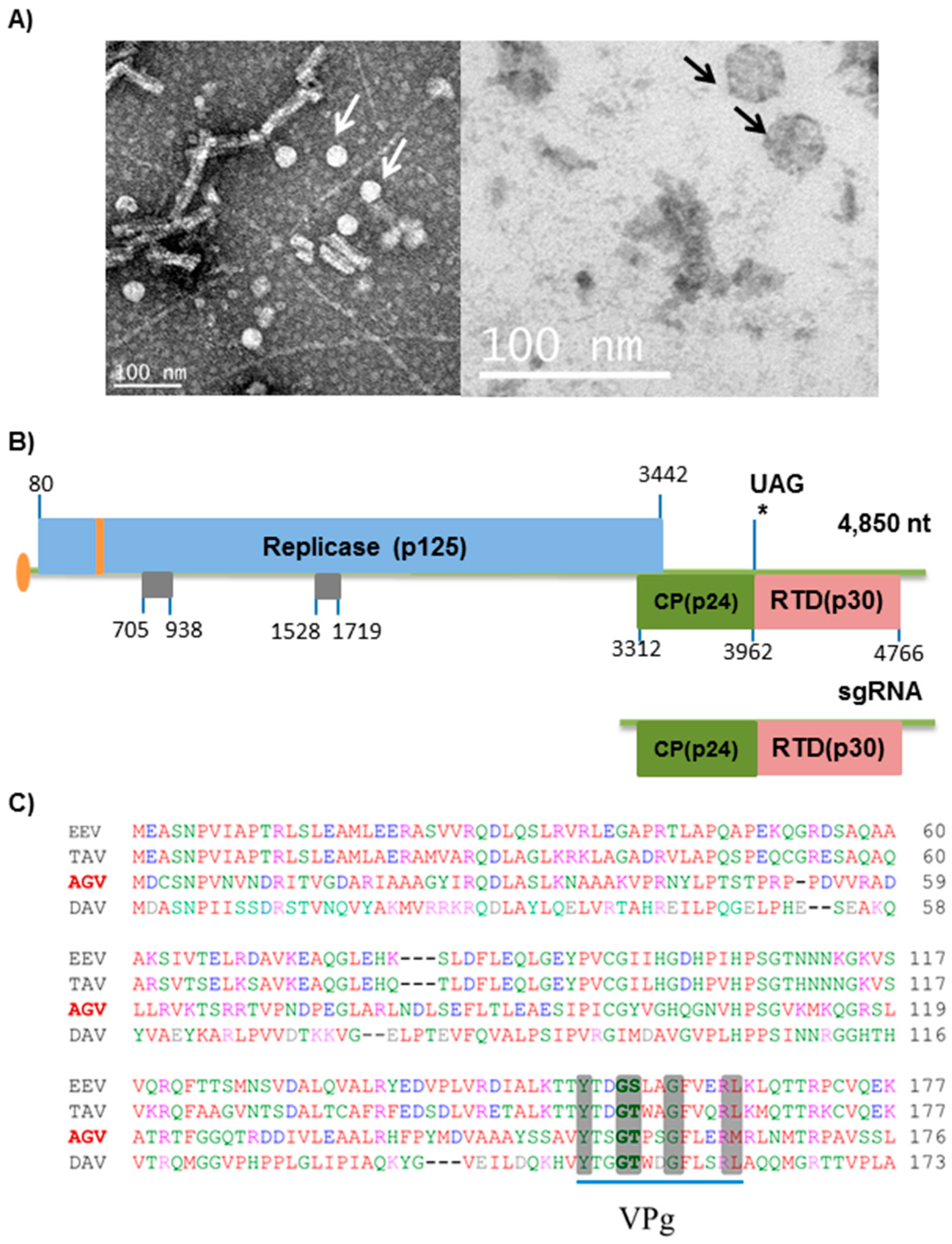
3.1. Discovery and Confirmation of the ApGlV2 Genome Sequence
3.2. ApGlV2 Genome Organization
3.3. Analysis of Putative Subgenomic RNA
3.4. ApGlV2 Encodes a Permuted RdRp
3.5. Analysis of ApGlV2 CP and Putative RTD
3.6. Distribution and Transmission of ApGlV2
3.7. ApGlV2 Is Not Detected in the Genome of A. glycines
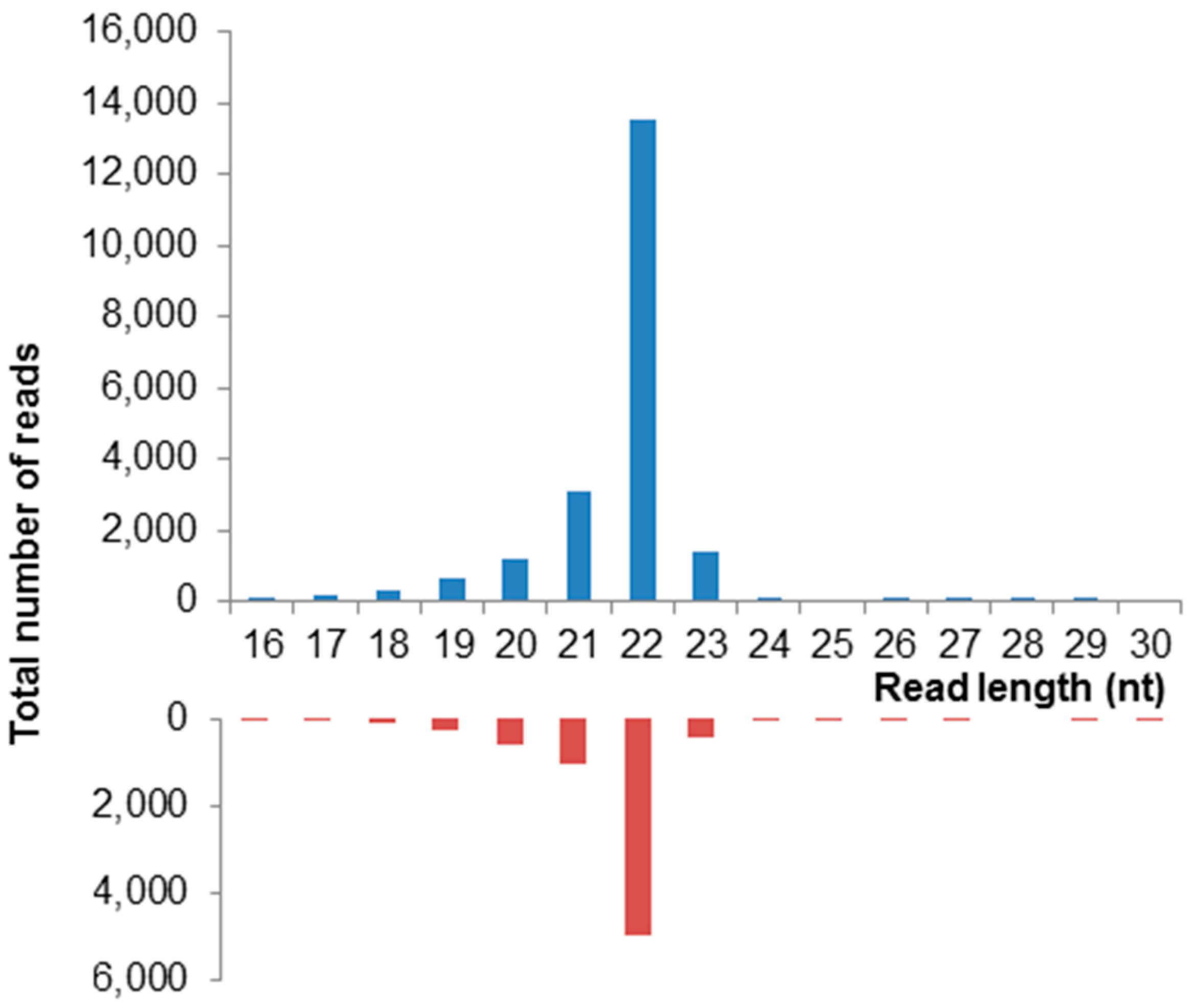
3.8. ApGlV2 Is Targeted by the RNAi Pathway
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Will, T.; Vilcinskas, A. Aphid-proof plants: Biotechnology-based approaches for aphid control. Adv. Biochem. Eng. Biotechnol. 2013, 136, 179–203. [Google Scholar] [PubMed]
- Nault, L.R. Arthropod transmission of plant viruses: A new synthesis. Annu. Entomol. Soc. Am. 1997, 90, 521–541. [Google Scholar] [CrossRef]
- Ragsdale, D.W.; Landis, D.A.; Brodeur, J.; Heimpel, G.E.; Desneux, N. Ecology and management of the soybean aphid in North America. Annu. Rev. Entomol. 2011, 56, 375–399. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Schaible, G.; Lynn, G.; Lubowski, R.; Lee, D. Economic impacts of the U.S. soybean aphid infestation: A multi-regional competitive dynamic analysis. Agric. Resour. Econ. Rev. 2008, 37, 227–242. [Google Scholar] [CrossRef]
- Michel, A.P.; Mittapalli, O.; Mian, R.M.A. Evolution of Soybean Aphid Biotypes: Understanding and Managing Virulence to Host-Plant Resistance; InTech: West Palm Beach, FL, USA, 2011; pp. 355–372. [Google Scholar]
- Van den Heuvel, J.F.; Hummelen, H.; Verbeek, M.; Dullemans, A.M.; van der Wilk, F. Characteristics of Acyrthosipon pisum virus, a newly identified virus infecting the pea aphid. J. Invertebr. Pathol. 1997, 70, 169–176. [Google Scholar] [CrossRef] [PubMed]
- D’Arcy, C.J.; Burnett, P.A.; Hewings, A.D. Detection, biological effects, and transmission of a virus of the aphid Rhopalosiphum padi. Virology 1981, 114, 268–272. [Google Scholar] [CrossRef]
- Ryabov, E.V. A novel virus isolated from the aphid Brevicoryne brassicae with similarity to Hymenoptera picorna-like viruses. J. Gen. Virol. 2007, 88, 2590–2595. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.V.; Keane, G.; Naish, N.; Evered, C.; Winstanley, D. Densovirus induces winged morphs in asexual clones of the rosy apple aphid, Dysaphis plantaginea. Proc. Natl. Acad. Sci. USA 2009, 106, 8465–8470. [Google Scholar] [CrossRef] [PubMed]
- Van Munster, M.; Dullemans, A.M.; Verbeek, M.; van den Heuvel, J.F.; Reinbold, C.; Brault, V.; Clerivet, A.; van der Wilk, F. A new virus infecting Myzus persicae has a genome organization similar to the species of the genus Densovirus. J. Gen. Virol. 2003, 84, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chougule, N.P.; Vijayendran, D.; Bonning, B.C. Deep sequencing of the transcriptomes of soybean aphid and associated endosymbionts. PLoS ONE 2012, 7, e45161. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Vijayendran, D.; Bonning, B.C. Next generation sequencing technologies for insect virus discovery. Viruses 2011, 3, 1849–1869. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, Y.; Bonning, B.C. RNA virus discovery in insects. Curr. Opin. Insect Sci. 2015, 8, 54–61. [Google Scholar] [CrossRef]
- Zeddam, J.L.; Gordon, K.H.; Lauber, C.; Alves, C.A.; Luke, B.T.; Hanzlik, T.N.; Ward, V.K.; Gorbalenya, A.E. Euprosterna elaeasa virus genome sequence and evolution of the Tetraviridae family: Emergence of bipartite genomes and conservation of the VPg signal with the dsRNA Birnaviridae family. Virology 2010, 397, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Gorbalenya, A.E.; Pringle, F.M.; Zeddam, J.L.; Luke, B.T.; Cameron, C.E.; Kalmakoff, J.; Hanzlik, T.N.; Gordon, K.H.; Ward, V.K. The palm subdomain-based active site is internally permuted in viral RNA-dependent RNA polymerases of an ancient lineage. J. Mol. Biol. 2002, 324, 47–62. [Google Scholar] [CrossRef]
- Ambrose, R.L.; Lander, G.C.; Maaty, W.S.; Bothner, B.; Johnson, J.E.; Johnson, K.N. Drosophila A virus is an unusual RNA virus with a T = 3 icosahedral core and permuted RNA-dependent RNA polymerase. J. Gen. Virol. 2009, 90, 2191–2200. [Google Scholar] [CrossRef] [PubMed]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Vijayendran, D.; Carrillo-Tripp, J.; Miller, W.A.; Bonning, B.C. Analysis of new aphid lethal paralysis virus (ALPV) isolates suggests evolution of two ALPV species. J. Gen. Virol. 2014, 95, 2809–2819. [Google Scholar] [CrossRef] [PubMed]
- Goic, B.; Vodovar, N.; Mondotte, J.A.; Monot, C.; Frangeul, L.; Blanc, H.; Gausson, V.; Vera-Otarola, J.; Cristofari, G.; Saleh, M.C. RNA-mediated interference and reverse transcription control the persistence of RNA viruses in the insect model Drosophila. Nat. Immunol. 2013, 14, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Hall Bioedit 7.0.5. Available online: http://www.mbio.ncsu.edu/BioEdit/Bioedit.html (accessed on 15 June 2015).
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhang, Y. LOMETS: A local meta-threading-server for protein structure prediction. Nucleic Acids Res. 2007, 35, 3375–3382. [Google Scholar] [CrossRef] [PubMed]
- Expasy search domain program. Available online: http://prosite.expasy.org/ (accessed on 5 May 2015).
- ICTV International Committee on Taxonomy of Viruses—Virus Taxonomy: 2015 Release. Available online: http://www.ictvonline.org/virustaxonomy.asp (accessed on 28 August 2015).
- Bruyère, A.; Brault, V.; Ziegler-Graff, V.; Simonis, M.T.; Van den Heuvel, J.F.; Richards, K.; Guilley, H.; Jonard, G.; Herrbach, E. Effects of mutations in the beet western yellows virus readthrough protein on its expression and packaging and on virus accumulation, symptoms, and aphid transmission. Virology 1997, 230, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Firth, A.E.; Brierley, I. Non-canonical translation in RNA viruses. J. Gen. Virol. 2012, 93, 1385–1409. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Victoria, J.G.; Wang, C.; Jones, M.; Fellers, G.M.; Kunz, T.H.; Delwart, E. Bat guano virome: Predominance of dietary viruses from insects and plants plus novel mammalian viruses. J. Virol. 2010, 84, 6955–6965. [Google Scholar] [CrossRef] [PubMed]
- Brault, V.; van den Heuvel, J.F.; Verbeek, M.; Ziegler-Graff, V.; Reutenauer, A.; Herrbach, E.; Garaud, J.C.; Guilley, H.; Richards, K.; Jonard, G. Aphid transmission of beet western yellows luteovirus requires the minor capsid read-through protein P74. EMBO J. 1995, 14, 650–659. [Google Scholar] [PubMed]
- Chay, C.A.; Gunasinge, U.B.; Dinesh-Kumar, S.P.; Miller, W.A.; Gray, S.M. Aphid transmission and systemic plant infection determinants of barley yellow dwarf luteovirus-PAV are contained in the coat protein readthrough domain and 17-kDa protein, respectively. Virology 1996, 219, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Peter, K.A.; Liang, D.; Palukaitis, P.; Gray, S.M. Small deletions in the potato leafroll virus readthrough protein affect particle morphology, aphid transmission, virus movement and accumulation. J. Gen. Virol. 2008, 89, 2037–2045. [Google Scholar] [CrossRef] [PubMed]
- Gildow, F.E.; D’Arcy, C.J. Cytopathology and experimental host range of Rhopalosiphum padi virus, a small isometric RNA virus infecting cereal grain aphids. J. Invertebr. Pathol. 1990, 55, 245–257. [Google Scholar] [CrossRef]
- Van Munster, M.; Janssen, A.; Clerivet, A.; van den Heuvel, J. Can plants use an entomopathogenic virus as a defense against herbivores? Oecologia 2005, 143, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Maori, E.; Tanne, E.; Sela, I. Reciprocal sequence exchange between non-retro viruses and hosts leading to the appearance of new host phenotypes. Virology 2007, 362, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Coates, B.S.; O’Neal, M.E.; Michel, A.P. Genome resequencing of the soybean aphid, Aphis glycines. (Unpublished; manuscript in preparation).
- Vijayendran, D.; Airs, P.M.; Dolezal, K.; Bonning, B.C. Arthropod viruses and small RNAs. J. Invertebr. Pathol. 2013, 114, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Poch, O.; Sauvaget, I.; Delarue, M.; Tordo, N. Identification of four conserved motifs among the RNA-dependent polymerase encoding elements. EMBO J. 1989, 8, 3867–3874. [Google Scholar] [PubMed]
- Koonin, E.V. The phylogeny of RNA-dependent RNA polymerases of positive-strand RNA viruses. J. Gen. Virol. 1991, 72 Pt 9, 2197–2206. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Dolja, V.V. Evolution and taxonomy of positive-strand RNA viruses: Implications of comparative analysis of amino acid sequences. Crit. Rev. Biochem. Mol. Biol. 1993, 28, 375–430. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.L.; Long, A.M.; Schultz, S.C. Structure of the RNA-dependent RNA polymerase of poliovirus. Structure 1997, 5, 1109–1122. [Google Scholar] [CrossRef]
- Van Dijk, A.A.; Makeyev, E.V.; Bamford, D.H. Initiation of viral RNA-dependent RNA polymerization. J. Gen. Virol. 2004, 85, 1077–1093. [Google Scholar] [CrossRef] [PubMed]
- Vasilakis, N.; Forrester, N.L.; Palacios, G.; Nasar, F.; Savji, N.; Rossi, S.L.; Guzman, H.; Wood, T.G.; Popov, V.; Gorchakov, R.; et al. Negevirus: A proposed new taxon of insect-specific viruses with wide geographic distribution. J. Virol. 2013, 87, 2475–2488. [Google Scholar] [CrossRef] [PubMed]
- Auguste, A.J.; Carrington, C.V.; Forrester, N.L.; Popov, V.L.; Guzman, H.; Widen, S.G.; Wood, T.G.; Weaver, S.C.; Tesh, R.B. Characterization of a novel Negevirus and a novel Bunyavirus isolated from Culex (Culex) declarator mosquitoes in Trinidad. J. Gen. Virol. 2014, 95, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Nabeshima, T.; Inoue, S.; Okamoto, K.; Posadas-Herrera, G.; Yu, F.; Uchida, L.; Ichinose, A.; Sakaguchi, M.; Sunahara, T.; Buerano, C.C.; et al. Tanay virus, a new species of virus isolated from mosquitoes in the Philippines. J. Gen. Virol. 2014, 95, 1390–1395. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Vakharia, V.N.; Tao, Y.J. The structure of a birnavirus polymerase reveals a distinct active site topology. Proc. Natl. Acad. Sci. USA 2007, 104, 7385–7390. [Google Scholar] [CrossRef] [PubMed]
- Garriga, D.; Navarro, A.; Querol-Audí, J.; Abaitua, F.; Rodríguez, J.F.; Verdaguer, N. Activation mechanism of a noncanonical RNA-dependent RNA polymerase. Proc. Natl. Acad. Sci. USA 2007, 104, 20540–20545. [Google Scholar] [CrossRef] [PubMed]
- Webster, C.L.; Waldron, F.M.; Robertson, S.; Crowson, D.; Ferrari, G.; Quintana, J.F.; Brouqui, J.M.; Bayne, E.H.; Longdon, B.; Buck, A.H.; et al. The discovery, distribution, and evolution of viruses associated with Drosophila melanogaster. PLoS Biol. 2015, 13, e1002210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hang, J.; Klein, T.A.; Kim, H.C.; Yang, Y.; Jima, D.D.; Richardson, J.H.; Jarman, R.G. Genome sequences of five arboviruses in field-captured mosquitoes in a unique rural environment of South Korea. Genome Announc. 2016, 4, e01644-15. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Luo, Y.; Lu, R.; Lau, N.; Lai, E.C.; Li, W.X.; Ding, S.W. Virus discovery by deep sequencing and assembly of virus-derived small silencing RNAs. Proc. Natl. Acad. Sci. USA 2010, 107, 1606–1611. [Google Scholar] [CrossRef] [PubMed]
- Mayo, M.A.; Miller, W.A. The structure and expression of luteovirus and related genomes. In Luteoviruses; Smith, H.G., Barker, H., Jones, P., Eds.; CAB International: Wallingford, UK, 1998. [Google Scholar]
- Rush, C.M. Ecology and epidemiology of benyviruses and plasmodiophorid vectors. Annu. Rev. Phytopathol. 2003, 41, 567–592. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.G.; Barker, H. The Luteoviridae; CABI: Wallingford, UK, 1999. [Google Scholar]
- Rumnieks, J.; Tars, K. Crystal structure of the read-through domain from bacteriophage Qβ A1 protein. Protein Sci. 2011, 20, 1707–1712. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.; Gildow, F.E. Luteovirus-aphid interactions. Annu. Rev. Phytopathol. 2003, 41, 539–566. [Google Scholar] [CrossRef] [PubMed]
- McGrann, G.R.; Grimmer, M.K.; Mutasa-Göttgens, E.S.; Stevens, M. Progress towards the understanding and control of sugar beet rhizomania disease. Mol. Plant Pathol. 2009, 10, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Norén, O.; Sjöström, H. Structure, biosynthesis and regulation of lactase-phlorizin hydrolase. Food Nutr. Res. 2001, 45, 156–160. [Google Scholar] [CrossRef]
- Richards, S.; Gibbs, R.A.; Gerardo, N.M.; Moran, N.; Nakabachi, A.; Stern, D.; Tagu, D.; Wilson, A.C.C.; Muzny, D.; Kovar, C.; et al. Genome Sequence of the pea aphid Acyrthosiphon pisum. PLoS Biol. 2010, 8, e1000313. [Google Scholar]
- Nayak, A.; Berry, B.; Tassetto, M.; Kunitomi, M.; Acevedo, A.; Deng, C.; Krutchinsky, A.; Gross, J.; Antoniewski, C.; Andino, R. Cricket paralysis virus antagonizes Argonaute 2 to modulate antiviral defense in Drosophila. Nat. Struct. Mol. Biol. 2010, 17, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Van Rij, R.P.; Saleh, M.C.; Berry, B.; Foo, C.; Houk, A.; Antoniewski, C.; Andino, R. The RNA silencing endonuclease Argonaute 2 mediates specific antiviral immunity in Drosophila melanogaster. Genes Dev. 2006, 20, 2985–2995. [Google Scholar] [CrossRef] [PubMed]
- Gordon, K.H.; Hanzlik, T.N. Tetraviruses. In The Insect Viruses; Springer: Berlin, Germany, 1998; pp. 269–299. [Google Scholar]
- Sparks, M.E.; Gundersen-Rindal, D.E.; Harrison, R.L. Complete genome sequence of a novel iflavirus from the transcriptome of Halyomorpha halys, the brown marmorated stink bug. Genome Announc. 2013, 1, e00910-13. [Google Scholar] [CrossRef] [PubMed]
- Murakami, R.; Suetsugu, Y.; Kobayashi, T.; Nakashima, N. The genome sequence and transmission of an iflavirus from the brown planthopper, Nilaparvata lugens. Virus Res. 2013, 176, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Tripp, J.; Bonning, B.C.; Miller, W.A. Challenges associated with research on RNA viruses of insects. Curr. Opin. Insect Sci. 2015, 8, 62–68. [Google Scholar] [CrossRef]
- Bonning, B.C. Editorial overview: Virus-insect interactions: Progress and pitfalls. Curr. Opin. Insect Sci. 2015, 8, 7–9. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′–3′) | ApGlV2 Genome Position |
|---|---|---|
| Tet-F | AGTGGCTGCGCATGCTCGTT | 1695–1714 |
| Tet-R | ACGCGCCTCTCCGTTGAACT | 2604–2623 |
| ApGlV2 3′-F | CAGTACAGCAATACGGCTCATT | 4284–4305 |
| ApGlV2 3′-R | AAGGGCCTTATTACTTTTCACACTCTCTC | 4474–4502 |
| ApGlV2 5′-F | GCAGGACCTTGCCTCGCTCAAA | 160–181 |
| ApGlV2-RdRp-F | AGTGGCTGCGCATGCTCGTT | 1695–2014 |
| ApGlV2-RdRp-R | ACGCGCCTCTCCGTTGAACT | 2603–2623 |
| ApGlV2 R/C-F | CACGCGCGGAATCTTTGCAG | 2976–2995 |
| ApGlV2 R/C-R | TCGGTCTTGGCGGCGTCATA | 3825–3845 |
| ApGlV2-CP-F | AGCAGAGCTCAACACGACGAACCAAG | 3315–3329 |
| ApGlV2-CP-R | AGCAAGCTTCTAAGCTCTCGTG | 3953–3965 |
| ApGlV2 C/T-F | TGTGACTCCGACACCGTCGAA | 3885–3905 |
| ApGlV2 C/T-R | GCACCGGGAGAAATCCCAGAGT | 4424–4445 |
| ApGlV2 3′ RACE | TCTCCCGGTGCCTCGTCTCACCACAGG | 4435–4461 |
| ApGlV2-sgRNA 5′ RACE | CCCGAGAGTTTTATTTATGCTGGTGGACGATATGGGCAGAGACA | 3748–3791 |
| ApGlV2 5′ RACE | AAGTGCCGTAGCGCTGCCTCGAGCAC | 476–501 |
| ALPV-F | TGAACTTCGTGCAACGAACACTGTT | N/A |
| ALPV-R | TCCGCCTGCGTTAGGAAGAAGA | N/A |
| RhPV-F | AATCTGGCGTTGACGCGCTC | N/A |
| RhPV-R | TCCCCCATCATCAACATAGATGCGT | N/A |
| qRT-PCR ApGlV2-F | TCCCCGCCACGTGAAGTGAA | 2624–2643 |
| qRT-PCR ApGlV2-R | GCTACTGCGTGCGTGGTGAA | 2822–2841 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Vijayendran, D., 1; Chen, Y.; Bonning, B.C. Aphis Glycines Virus 2, a Novel Insect Virus with a Unique Genome Structure. Viruses 2016, 8, 315. https://doi.org/10.3390/v8110315
Liu S, Vijayendran D 1, Chen Y, Bonning BC. Aphis Glycines Virus 2, a Novel Insect Virus with a Unique Genome Structure. Viruses. 2016; 8(11):315. https://doi.org/10.3390/v8110315
Chicago/Turabian StyleLiu, Sijun, Diveena Vijayendran, 1, Yuting Chen, and Bryony C. Bonning. 2016. "Aphis Glycines Virus 2, a Novel Insect Virus with a Unique Genome Structure" Viruses 8, no. 11: 315. https://doi.org/10.3390/v8110315
APA StyleLiu, S., Vijayendran, D., 1, Chen, Y., & Bonning, B. C. (2016). Aphis Glycines Virus 2, a Novel Insect Virus with a Unique Genome Structure. Viruses, 8(11), 315. https://doi.org/10.3390/v8110315