
Characterization of an Immunodominant Epitope in the Endodomain of the Coronavirus Membrane Protein

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Cells, Antibodies, and Virus
2.2. Recombinant Plasmid Construction and Recombinant Protein Expression
2.3. Preparation of mAbs Targeting the M Protein
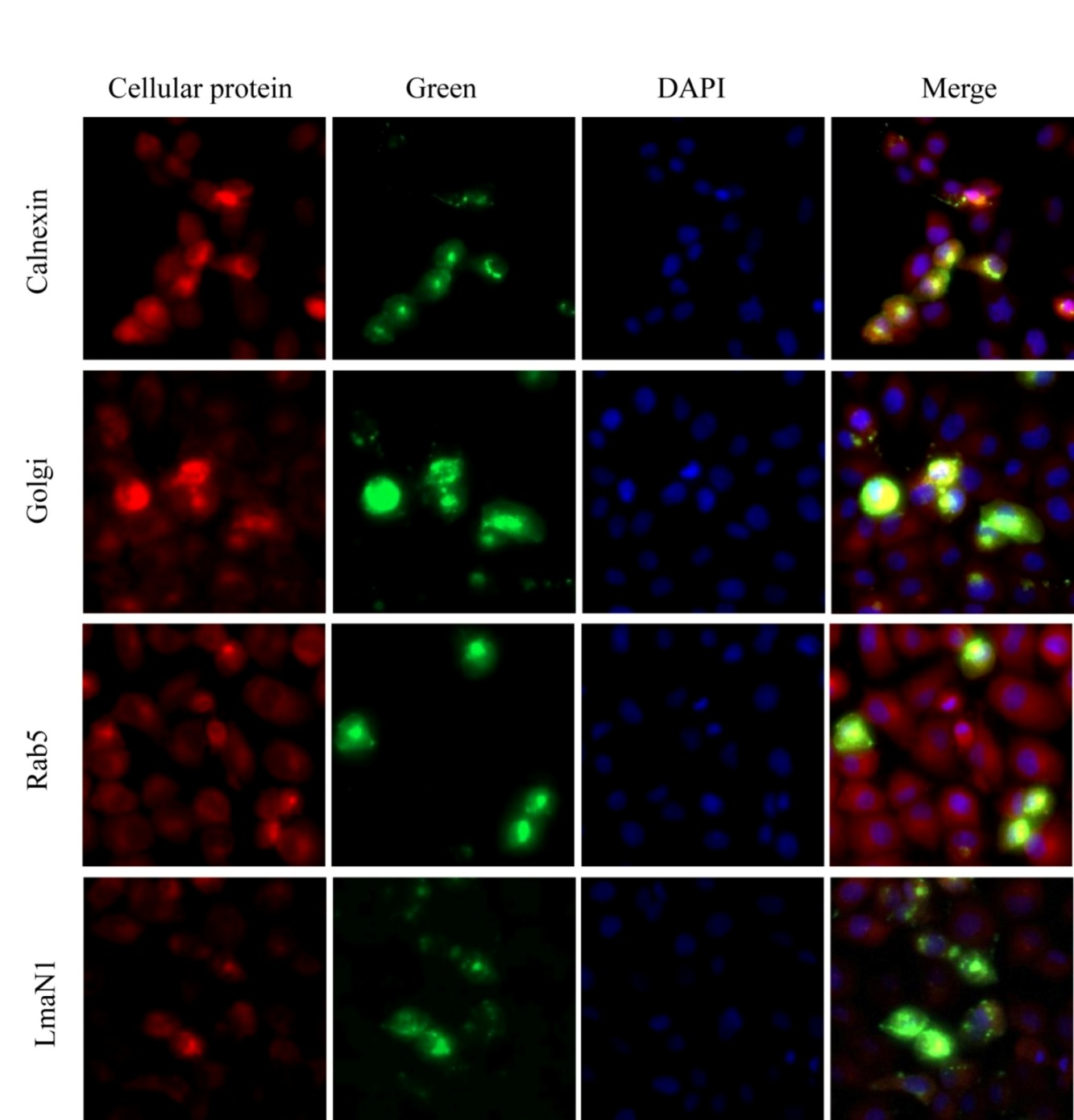
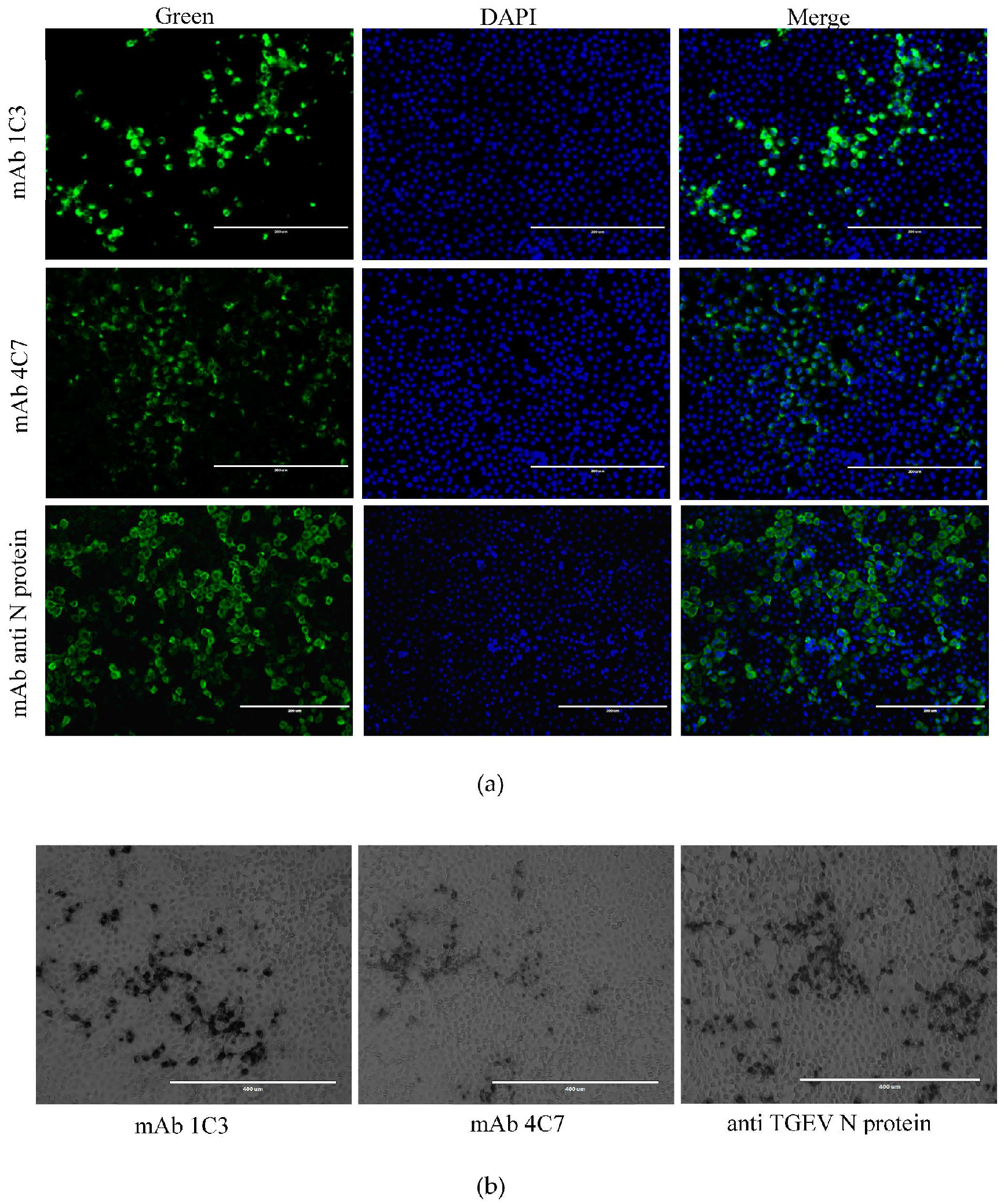
2.4. Immunofluorescence Assay (IFA)
2.5. Immunoperoxidase Monolayer Assay (IPMA)
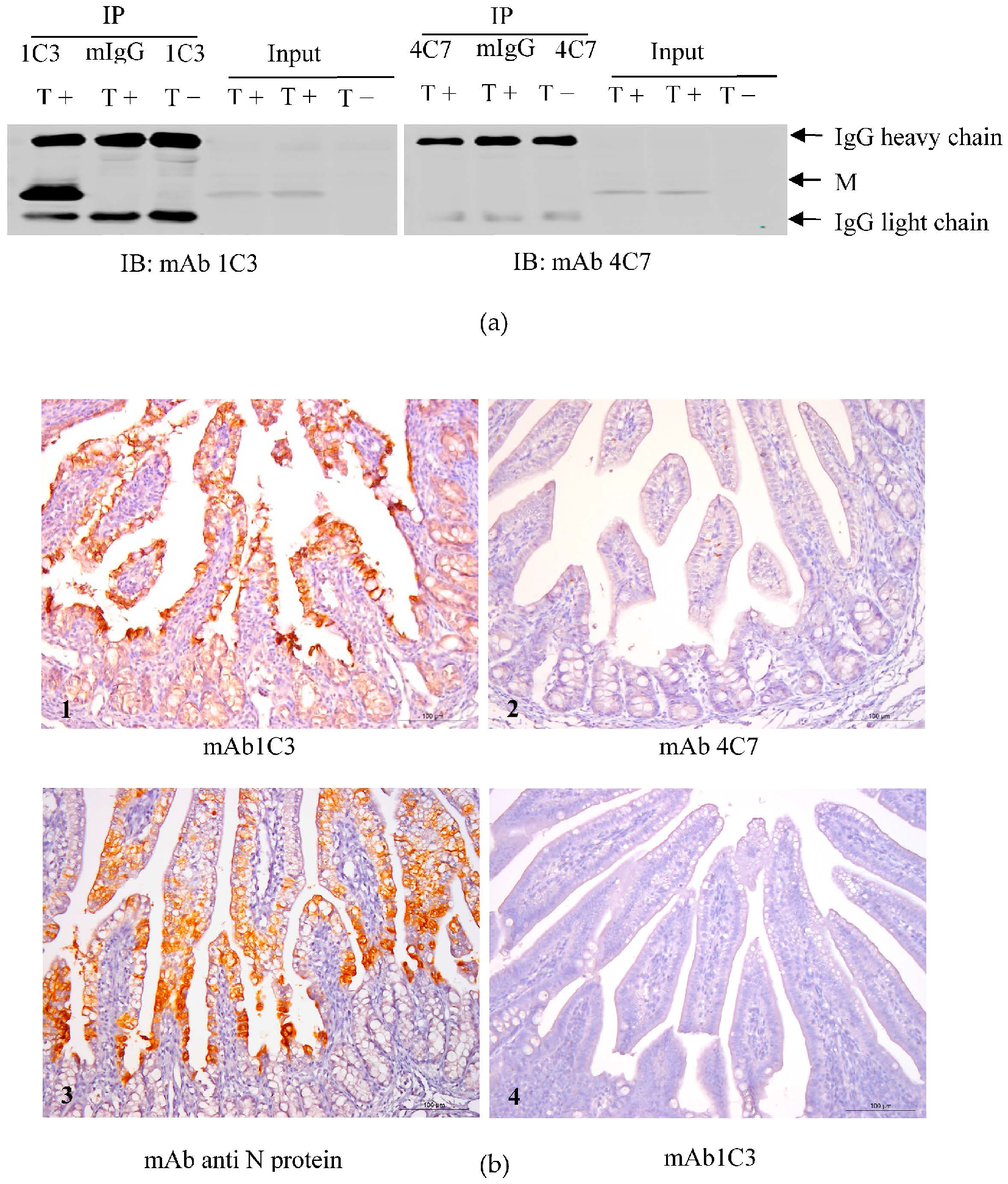
2.6. Immunoprecipitation of the TGEV M Protein
2.7. Polypeptide Design and Coupling
2.8. Animal Immunization with RS-15-KLH, YT-16-KLH, and YV-20-KLH
2.9. Peptide ELISA
2.10. Immunohistochemistry (IHC)
2.11. 3D Epitope Modelling
2.12. Animal Ethics
3. Results
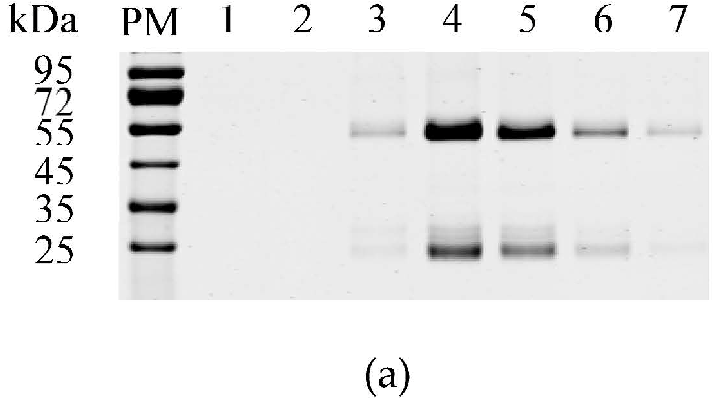
3.1. Expression and Purification of the GST-M Protein
3.2. Preparation of mAbs against the TGEV M Protein
3.3. Determination of the 1C3 and 4C7 mAb Epitopes
3.4. 3D Epitope Mapping
3.5. Reactivity of 1C3 and 4C7 with the TGEV M Protein in IFA and IPMA
3.6. Immunoprecipitation of 1C3 and 4C7 with the TGEV M Protein
3.7. mAb 1C3 Reacted with the M Protein in the Small Intestine
3.8. Optimizing of the IFA Method for the Detection of the M Protein
3.9. Antibody Responses to the Identified Epitopes
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- De Groot, R.J.; Baker, S.G.; Baric, R.S.; Enjuanes, L.; Gorbalenya, A.E. Coronaviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2011; pp. 774–796. [Google Scholar]
- Reguera, J.; Santiago, C.; Mudgal, G.; Ordono, D.; Enjuanes, L.; Casasnovas, J.M. Structural bases of coronavirus attachment to host aminopeptidase N and its inhibition by neutralizing antibodies. PLoS Pathog. 2012, 8, e1002859. [Google Scholar] [CrossRef] [PubMed]
- Perlman, S.; Netland, J. Coronaviruses post-SARS: Update on replication and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.M.; Cavanagh, D. The molecular biology of coronaviruses. Adv. Virus Res. 1997, 48, 1–100. [Google Scholar] [PubMed]
- Yang, D.; Leibowitz, J.L. The structure and functions of coronavirus genomic 3′ and 5′ ends. Virus Res. 2015, 206, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Jenwitheesuk, E.; Samudrala, R. Identifying inhibitors of the SARS coronavirus proteinase. Bioorg. Med. Chem. Lett. 2003, 13, 3989–3992. [Google Scholar] [CrossRef] [PubMed]
- Anand, K.; Ziebuhr, J.; Wadhwani, P.; Mesters, J.R.; Hilgenfeld, R. Coronavirus main proteinase (3CLpro) structure: Basis for design of anti-SARS drugs. Science 2003, 300, 1763–1767. [Google Scholar] [CrossRef] [PubMed]
- Yount, B.; Curtis, K.M.; Baric, R.S. Strategy for systematic assembly of large RNA and DNA genomes: Transmissible gastroenteritis virus model. J. Virol. 2000, 74, 10600–10611. [Google Scholar] [PubMed]
- Sola, I.; Almazan, F.; Zuniga, S.; Enjuanes, L. Continuous and discontinuous RNA synthesis in coronaviruses. Annu. Rev. Virol. 2015, 2, 265–288. [Google Scholar] [CrossRef] [PubMed]
- Cruz, J.L.; Sola, I.; Becares, M.; Alberca, B.; Plana, J.; Enjuanes, L.; Zuñiga, S. Coronavirus gene 7 counteracts host defenses and modulates virus virulence. PLoS Pathog. 2011, 7, e1002090. [Google Scholar] [CrossRef] [PubMed]
- Zuniga, S.; Sola, I.; Moreno, J.L.; Sabella, P.; Plana-Duran, J.; Enjuanes, L. Coronavirus nucleocapsid protein is an RNA chaperone. Virology 2007, 357, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Almazan, F.; Gonzalez, J.M.; Penzes, Z.; Izeta, A.; Calvo, E.; Plana-Duran, J.; Enjuanes, L. Engineering the largest RNA virus genome as an infectious bacterial artificial chromosome. Proc. Natl. Acad. Sci. USA 2000, 97, 5516–5521. [Google Scholar] [CrossRef] [PubMed]
- McBride, C.E.; Machamer, C.E. A single tyrosine in the severe acute respiratory syndrome coronavirus membrane protein cytoplasmic tail is important for efficient interaction with spike protein. J. Virol. 2010, 84, 1891–1901. [Google Scholar] [CrossRef] [PubMed]
- Swift, A.M.; Machamer, C.E. A Golgi retention signal in a membrane-spanning domain of coronavirus E1 protein. J. Cell Biol. 1991, 115, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.; Patel, S.; Riddle, P. Lysosomal sorting mutants of coronavirus E1 protein, a Golgi membrane protein. J. Cell Sci. 1990, 95, 191–197. [Google Scholar] [PubMed]
- Locker, J.K.; Klumperman, J.; Oorschot, V.; Horzinek, M.C.; Geuze, H.J.; Rottier, P.J. The cytoplasmic tail of mouse hepatitis virus M protein is essential but not sufficient for its retention in the Golgi complex. J. Biol. Chem. 1994, 269, 28263–28269. [Google Scholar] [PubMed]
- Hsieh, Y.C.; Li, H.C.; Chen, S.C.; Lo, S.Y. Interactions between M protein and other structural proteins of severe, acute respiratory syndrome-associated coronavirus. J. Biomed. Sci. 2008, 15, 707–717. [Google Scholar] [CrossRef] [PubMed]
- Siu, Y.L.; Teoh, K.T.; Lo, J.; Chan, C.M.; Kien, F.; Escriou, N.; Tsao, S.W.; Nicholls, J.M.; Altmeyer, R.; Peiris, J.S. The M, E, and N structural proteins of the severe acute respiratory syndrome coronavirus are required for efficient assembly, trafficking, and release of virus-like particles. J. Virol. 2008, 82, 11318–11330. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.T.; Wang, S.M.; Huang, K.J.; Lee, A.I.; Chiang, C.C.; Wang, C.T. Self-assembly of severe acute respiratory syndrome coronavirus membrane protein. J. Biol. Chem. 2010, 285, 12862–12872. [Google Scholar] [CrossRef] [PubMed]
- Baudoux, P.; Carrat, C.; Besnardeau, L.; Charley, B.; Laude, H. Coronavirus pseudoparticles formed with recombinant M and E proteins induce alpha interferon synthesis by leukocytes. J. Virol. 1998, 72, 8636–8643. [Google Scholar] [PubMed]
- Huang, Y.; Yang, Z.Y.; Kong, W.P.; Nabel, G.J. Generation of synthetic severe acute respiratory syndrome coronavirus pseudoparticles: Implications for assembly and vaccine production. J. Virol. 2004, 78, 12557–12565. [Google Scholar] [CrossRef] [PubMed]
- Mortola, E.; Roy, P. Efficient assembly and release of SARS coronavirus-like particles by a heterologous expression system. FEBS Lett. 2004, 576, 174–178. [Google Scholar] [CrossRef] [PubMed]
- De Haan, C.A.; Vennema, H.; Rottier, P.J. Assembly of the coronavirus envelope: Homotypic interactions between the M proteins. J. Virol. 2000, 74, 4967–4978. [Google Scholar] [CrossRef] [PubMed]
- Opstelten, D.J.; Raamsman, M.J.; Wolfs, K.; Horzinek, M.C.; Rottier, P.J. Envelope glycoprotein interactions in coronavirus assembly. J. Cell Biol. 1995, 131, 339–349. [Google Scholar] [CrossRef] [PubMed]
- De Haan, C.A.; Smeets, M.; Vernooij, F.; Vennema, H.; Rottier, P.J. Mapping of the coronavirus membrane protein domains involved in interaction with the spike protein. J. Virol. 1999, 73, 7441–7452. [Google Scholar] [PubMed]
- Youn, S.; Collisson, E.W.; Machamer, C.E. Contribution of trafficking signals in the cytoplasmic tail of the infectious bronchitis virus spike protein to virus infection. J. Virol. 2005, 79, 13209–13217. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.P.; Hogue, B.G. Protein interactions during coronavirus assembly. J. Virol. 1997, 71, 9278–9284. [Google Scholar] [PubMed]
- Escors, D.; Ortego, J.; Laude, H.; Enjuanes, L. The membrane M protein carboxy terminus binds to transmissible gastroenteritis coronavirus core and contributes to core stability. J. Virol. 2001, 75, 1312–1324. [Google Scholar] [CrossRef] [PubMed]
- Rottier, P.; Brandenburg, D.; Armstrong, J.; van der Zeijst, B.; Warren, G. Assembly in vitro of a spanning membrane protein of the endoplasmic reticulum: The E1 glycoprotein of coronavirus mouse hepatitis virus A59. Proc. Natl. Acad. Sci. USA 1984, 81, 1421–1425. [Google Scholar] [CrossRef] [PubMed]
- Kapke, P.A.; Tung, F.Y.; Hogue, B.G.; Brian, D.A.; Woods, R.D.; Wesley, R. The amino-terminal signal peptide on the porcine transmissible gastroenteritis coronavirus matrix protein is not an absolute requirement for membrane translocation and glycosylation. Virology 1988, 165, 367–376. [Google Scholar] [CrossRef]
- Lopez, L.A.; Jones, A.; Arndt, W.D.; Hogue, B.G. Subcellular localization of SARS-CoV structural proteins. Adv. Exp. Med. Biol. 2006, 581, 297–300. [Google Scholar] [PubMed]
- Nal, B.; Chan, C.; Kien, F.; Siu, L.; Tse, J.; Chu, K.; Kam, J.; Staropoli, I.; Crescenzo-Chaigne, B.; Escriou, N.; et al. Differential maturation and subcellular localization of severe acute respiratory syndrome coronavirus surface proteins S, M and E. J. Gen. Virol. 2005, 86, 1423–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escors, D.; Camafeita, E.; Ortego, J.; Laude, H.; Enjuanes, L. Organization of two transmissible gastroenteritis coronavirus membrane protein topologies within the virion and core. J. Virol. 2001, 75, 12228–12240. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, H.; Chen, J.; Shi, D.; Li, C.; Feng, L. EF1A interacting with nucleocapsid protein of transmissible gastroenteritis coronavirus and plays a role in virus replication. Vet. Microbiol. 2014, 172, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Kohler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Jungmann, A.; Nieper, H.; Muller, H. Apoptosis is induced by infectious bursal disease virus replication in productively infected cells as well as in antigen-negative cells in their vicinity. J. Gen. Virol. 2001, 82, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qiu, H.; Zhang, M.; Cai, X.; Qu, Y.; Hu, D.; Zhao, X.; Zhou, E.; Liu, S.; Xiao, Y. Distribution of highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV) in different stages of gestation sows: HP-PRRSV distribution in gestation sows. Vet. Immunol. Immunopathol. 2015, 166, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Gallo Cassarino, T.; Bertoni, M.; Bordoli, L. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Love, M.L.; Szebenyi, D.M.; Kriksunov, I.A.; Thiel, D.J.; Munshi, C.; Graeff, R.; Lee, H.C.; Hao, Q. ADP-ribosyl cyclase; crystal structures reveal a covalent intermediate. Structure 2004, 12, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Yano, A.; Miwa, Y.; Kanazawa, Y.; Ito, K.; Makino, M.; Imai, S.; Hanada, N.; Nisizawa, T. A novel method for enhancement of peptide vaccination utilizing T-cell epitopes from conventional vaccines. Vaccine 2013, 31, 1510–1515. [Google Scholar] [CrossRef] [PubMed]
- Kuo, L.; Hurst-Hess, K.R.; Koetzner, C.A.; Masters, P.S. Analyses of coronavirus assembly interactions with interspecies membrane and nucleocapsid protein chimeras. J. Virol. 2016, 90, 4357–4368. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, J.; Shi, H.; Chen, X.; Shi, D.; Feng, L.; Yang, B. Identification of a conserved linear B-cell epitope in the M protein of porcine epidemic diarrhea virus. Virol. J. 2012, 9. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Liu, S.; Han, Z.; Shao, Y.; Li, H.; Kong, X. Identification of a novel linear B-cell epitope in the M protein of avian infectious bronchitis coronaviruses. J. Microbiol. 2009, 47, 589–599. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhou, Y.; Siddiqui, P.; Niu, J.; Jiang, S. Identification of immunodominant epitopes on the membrane protein of the severe acute respiratory syndrome-associated coronavirus. J. Clin. Microbiol. 2005, 43, 3718–3726. [Google Scholar] [CrossRef] [PubMed]
- Laviada, M.D.; Videgain, S.P.; Moreno, L.; Alonso, F.; Enjuanes, L.; Escribano, J.M. Expression of swine transmissible gastroenteritis virus envelope antigens on the surface of infected cells: Epitopes externally exposed. Virus Res. 1990, 16, 247–254. [Google Scholar] [CrossRef]
- De Diego, M.; Laviada, M.D.; Enjuanes, L.; Escribano, J.M. Epitope specificity of protective lactogenic immunity against swine transmissible gastroenteritis virus. J. Virol. 1992, 66, 6502–6508. [Google Scholar] [PubMed]
- Rodak, L.; Smid, B.; Nevorankova, Z.; Valicek, L.; Smitalova, R. Use of monoclonal antibodies in blocking ELISA detection of transmissible gastroenteritis virus in faeces of piglets. J. Vet. Med. B Infect. Dis. Vet. Public Health 2005, 52, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Rottier, P.J.; Welling, G.W.; Welling-Wester, S.; Niesters, H.G.; Lenstra, J.A.; van der Zeijst, B.A. Predicted membrane topology of the coronavirus protein E1. Biochemistry 1986, 25, 1335–1339. [Google Scholar] [CrossRef] [PubMed]
- Neuman, B.W.; Kiss, G.; Kunding, A.H.; Bhella, D.; Baksh, M.F.; Connelly, S.; Droese, B.; Klaus, J.P.; Makino, S.; Sawicki, S.G. A structural analysis of M protein in coronavirus assembly and morphology. J. Struct. Biol. 2011, 174, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Kuo, L.; Masters, P.S. Evolved variants of the membrane protein can partially replace the envelope protein in murine coronavirus assembly. J. Virol. 2010, 84, 12872–12885. [Google Scholar] [CrossRef] [PubMed]
- Kuo, L.; Masters, P.S. Genetic evidence for a structural interaction between the carboxy termini of the membrane and nucleocapsid proteins of mouse hepatitis virus. J. Virol. 2002, 76, 4987–4999. [Google Scholar] [CrossRef] [PubMed]
- Hurst, K.R.; Kuo, L.; Koetzner, C.A.; Ye, R.; Hsue, B.; Masters, P.S. A major determinant for membrane protein interaction localizes to the carboxy-terminal domain of the mouse coronavirus nucleocapsid protein. J. Virol. 2005, 79, 13285–13297. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Lopez, L.A.; Bednar, V.; Hogue, B.G. Importance of the penultimate positive charge in mouse hepatitis coronavirus A59 membrane protein. J. Virol. 2007, 81, 5339–5348. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wu, D.; Shen, C.; Chen, K.; Shen, X.; Jiang, H. Severe acute respiratory syndrome coronavirus membrane protein interacts with nucleocapsid protein mostly through their carboxyl termini by electrostatic attraction. Int. J. Biochem. Cell Biol. 2006, 38, 589–599. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Leeson, A.; Ballantine, M.; Andonov, A.; Baker, L.; Dobie, F.; Li, Y.; Bastien, N.; Feldmann, H.; Strocher, U. Characterization of protein-protein interactions between the nucleocapsid protein and membrane protein of the SARS coronavirus. Virus Res. 2004, 105, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Ye, L.; Timani, K.A.; Li, S.; Zen, Y.; Zhao, M.; Zheng, H.; Wu, Z. Peptide domain involved in the interaction between membrane protein and nucleocapsid protein of SARS-associated coronavirus. J. Biochem. Mol. Biol. 2005, 38, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Anton, I.M.; Gonzalez, S.; Bullido, M.J.; Corsin, M.; Risco, C.; Langeveld, J.P.; Enjuanes, L. Cooperation between transmissible gastroenteritis coronavirus (TGEV) structural proteins in the in vitro induction of virus-specific antibodies. Virus Res. 1996, 46, 111–124. [Google Scholar] [CrossRef]
- Kirchdoerfer, R.N.; Cottrell, C.A.; Wang, N.; Pallesen, J.; Yassine, H.M.; Turner, H.L.; Corbett, K.S.; Graham, B.S.; McLellan, J.S.; Ward, A.B. Pre-fusion structure of a human coronavirus spike protein. Nature 2016, 531, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Lu, G.; Qi, J.; Li, Y.; Wu, Y.; Deng, Y.; Geng, H.; Li, H.; Wang, Q.; Xiao, H. Structure of the fusion core and inhibition of fusion by a heptad repeat peptide derived from the S protein of Middle East respiratory syndrome coronavirus. J. Virol. 2013, 87, 13134–13140. [Google Scholar] [CrossRef] [PubMed]
- Wicht, O.; Burkard, C.; de Haan, C.A.; van Kuppeveld, F.J.; Rottier, P.J.; Bosch, B.J. Identification and characterization of a proteolytically primed form of the murine coronavirus spike proteins after fusion with the target cell. J. Virol. 2014, 88, 4943–4952. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, H.; Hattermann, K.; Marzi, A.; Gramberg, T.; Geier, M.; Krumbiegel, M.; Kuate, S.; Uberla, K.; Niedrig, M.; Pohlmann, S. S protein of severe acute respiratory syndrome-associated coronavirus mediates entry into hepatoma cell lines and is targeted by neutralizing antibodies in infected patients. J. Virol. 2004, 78, 6134–6142. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, L.; Qin, C.; Ba, L.; Yi, C.E.; Zhang, F.; Wei, Q.; He, T.; Yu, W.; Yu, J. Recombinant modified vaccinia virus Ankara expressing the spike glycoprotein of severe acute respiratory syndrome coronavirus induces protective neutralizing antibodies primarily targeting the receptor binding region. J. Virol. 2005, 79, 2678–2688. [Google Scholar] [CrossRef] [PubMed]
- Risco, C.; Anton, I.M.; Sune, C.; Pedregosa, A.M.; Martín-Alonso, J.M.; Parra, F.; Carrascosa, J.L.; Enjuanes, L. Membrane protein molecules of transmissible gastroenteritis coronavirus also expose the carboxy-terminal region on the external surface of the virion. J. Virol. 1995, 69, 5269–5277. [Google Scholar] [PubMed]
- Fan, J.H.; Zuo, Y.Z.; Shen, X.Q.; Gu, W.Y.; Di, J.M. Development of an enzyme-linked immunosorbent assay for the monitoring and surveillance of antibodies to porcine epidemic diarrhea virus based on a recombinant membrane protein. J. Virol. Methods 2015, 225, 90–94. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Enzyme |
|---|---|---|
| F-GST-M | CCGCTCGAGGAACGCTATTGTGC | Xho I |
| R-GST-M | CGGAATTCTTATACCATATGTA | Eco RI |
| F-M (49–228)-6p | GTGGATCCGAACGCTATTGTGCTATGAA | Bam HI |
| R-M (49–228)-6p | GACTCGAGGAATTGAGGTCTTCCATATT | Xho I |
| F-M (199–378)-6p | GTGGATCC ACTGTGCTACAATATGGAAG | Bam HI |
| R-M (199–378)-6p | GACTCGAGAAATGTAACAATTGCACCTG | Xho I |
| F-M (349–528)-6p | GTGGATCCTTTAGTATTGCAGGTGCAAT | Bam HI |
| R-M (349–528)-6p | GACTCGAGACCAGTTGGCACACCTTCGA | Xho I |
| F-M (499–678)-6p | GTGGATCCGTGCTTCCTCTCGAAGGTGT | Bam HI |
| R-M (499–678)-6p | GACTCGAGTGCTTTCAACTTCTTGCCAA | Xho I |
| F-M (649–789)-6p | GTGGATCCTACACACTTGTTGGCAAGAA | Bam HI |
| R-M (649–789)-6p | GACTCGAGTTATACCATATGTAATAATT | Xho I |
| F-M (649–738)-6p | GTGGATCCTACACACTTGTTGGCAAGAA | Bam HI |
| R-M (649–738)-6p | GACTCGAGCTCTGTTGAGTAATCACCAG | Xho I |
| F-M (700–789)-6p | GTGGATCCTACTATGTAAAATCTAAAGC | Bam HI |
| R-M (700–789)-6p | GACTCGAGTTATACCATATGTAATAATT | Xho I |
| Residues | Amino Acid Sequence | Residues | Amino Acid Sequence |
|---|---|---|---|
| 217–236 | YTLVGKKLKASSATGWAYYV | 230–249 | TGWAYYVKSKAGDYSTEART |
| 243–262 | YSTEARTDNLSEQEKLLHMV | 234–248 | YYVKSKAGDYSTEAR |
| 243–257 | YSTEARTDNLSEQEK | 244–258 | STEARTDNLSEQEKL |
| 245–259 | TEARTDNLSEQEKLL | 246–260 | EARTDNLSEQEKLLH |
| 247–261 | ARTDNLSEQEKLLHM | 248–262 | RTDNLSEQEKLLHMV |
| RS-15 | RGDYSTEARTGGGGS | YT-16 | YSTEARTGGYSTEART |
| YV-20 | YSTEARTDNLSEQEKLLHMV |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, H.; Zhang, X.; Shi, H.; Chen, J.; Shi, D.; Zhu, Y.; Feng, L. Characterization of an Immunodominant Epitope in the Endodomain of the Coronavirus Membrane Protein. Viruses 2016, 8, 327. https://doi.org/10.3390/v8120327
Dong H, Zhang X, Shi H, Chen J, Shi D, Zhu Y, Feng L. Characterization of an Immunodominant Epitope in the Endodomain of the Coronavirus Membrane Protein. Viruses. 2016; 8(12):327. https://doi.org/10.3390/v8120327
Chicago/Turabian StyleDong, Hui, Xin Zhang, Hongyan Shi, Jianfei Chen, Da Shi, Yunnuan Zhu, and Li Feng. 2016. "Characterization of an Immunodominant Epitope in the Endodomain of the Coronavirus Membrane Protein" Viruses 8, no. 12: 327. https://doi.org/10.3390/v8120327
APA StyleDong, H., Zhang, X., Shi, H., Chen, J., Shi, D., Zhu, Y., & Feng, L. (2016). Characterization of an Immunodominant Epitope in the Endodomain of the Coronavirus Membrane Protein. Viruses, 8(12), 327. https://doi.org/10.3390/v8120327