
Use of Reporter Genes in the Generation of Vaccinia Virus-Derived Vectors

Abstract
:1. Introduction
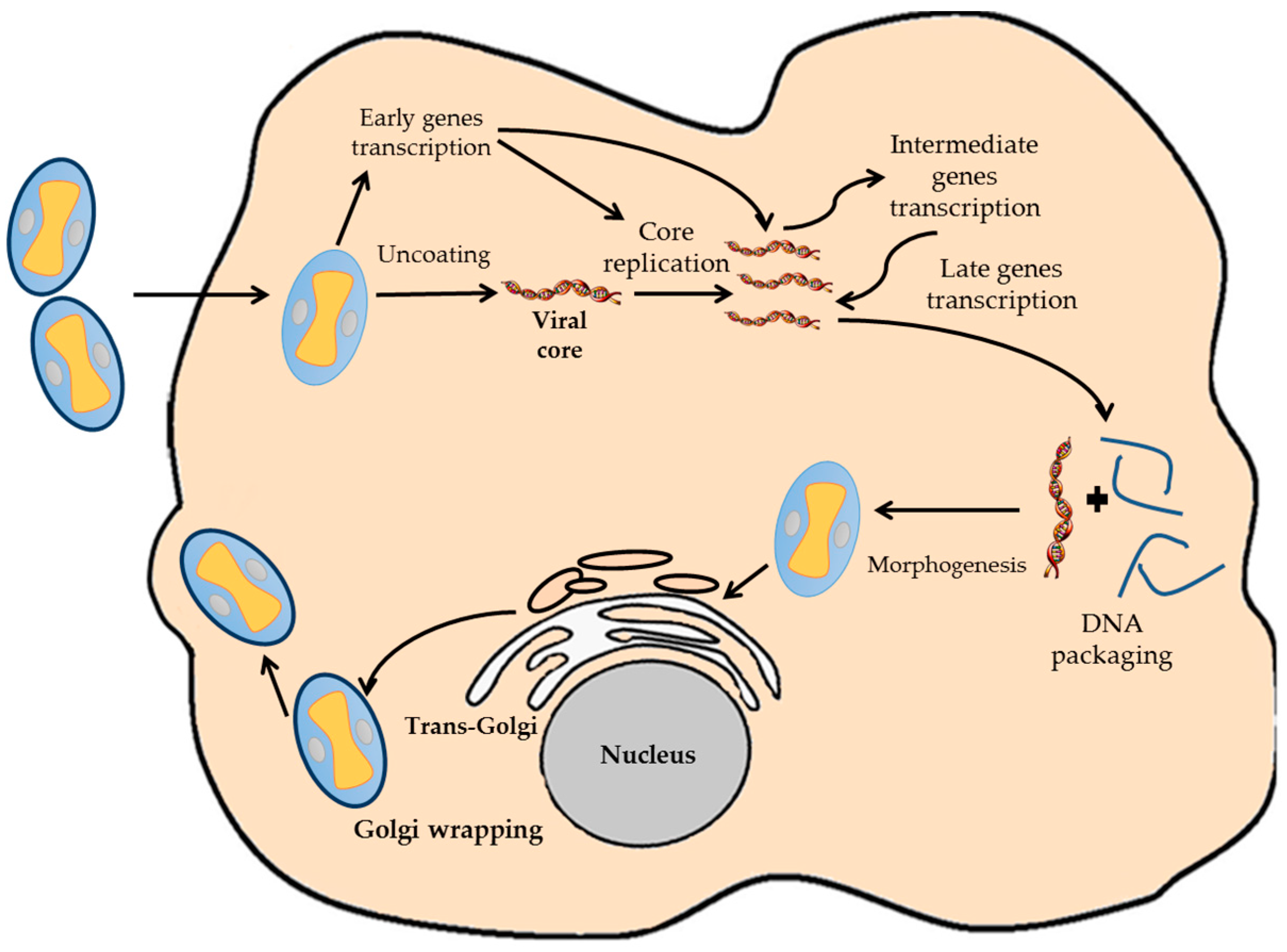
2. Biology of VACV
3. VACV as a Vector
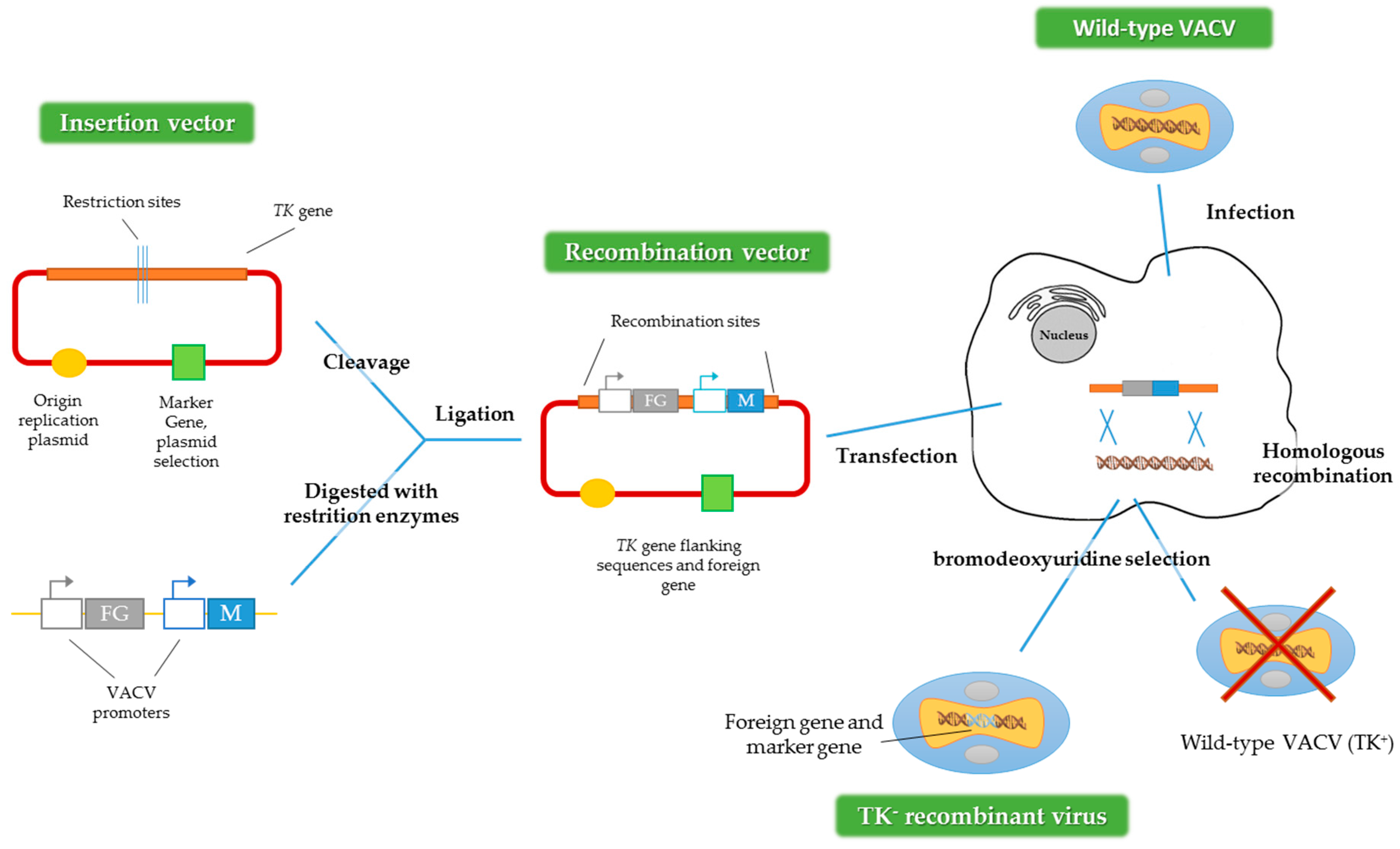
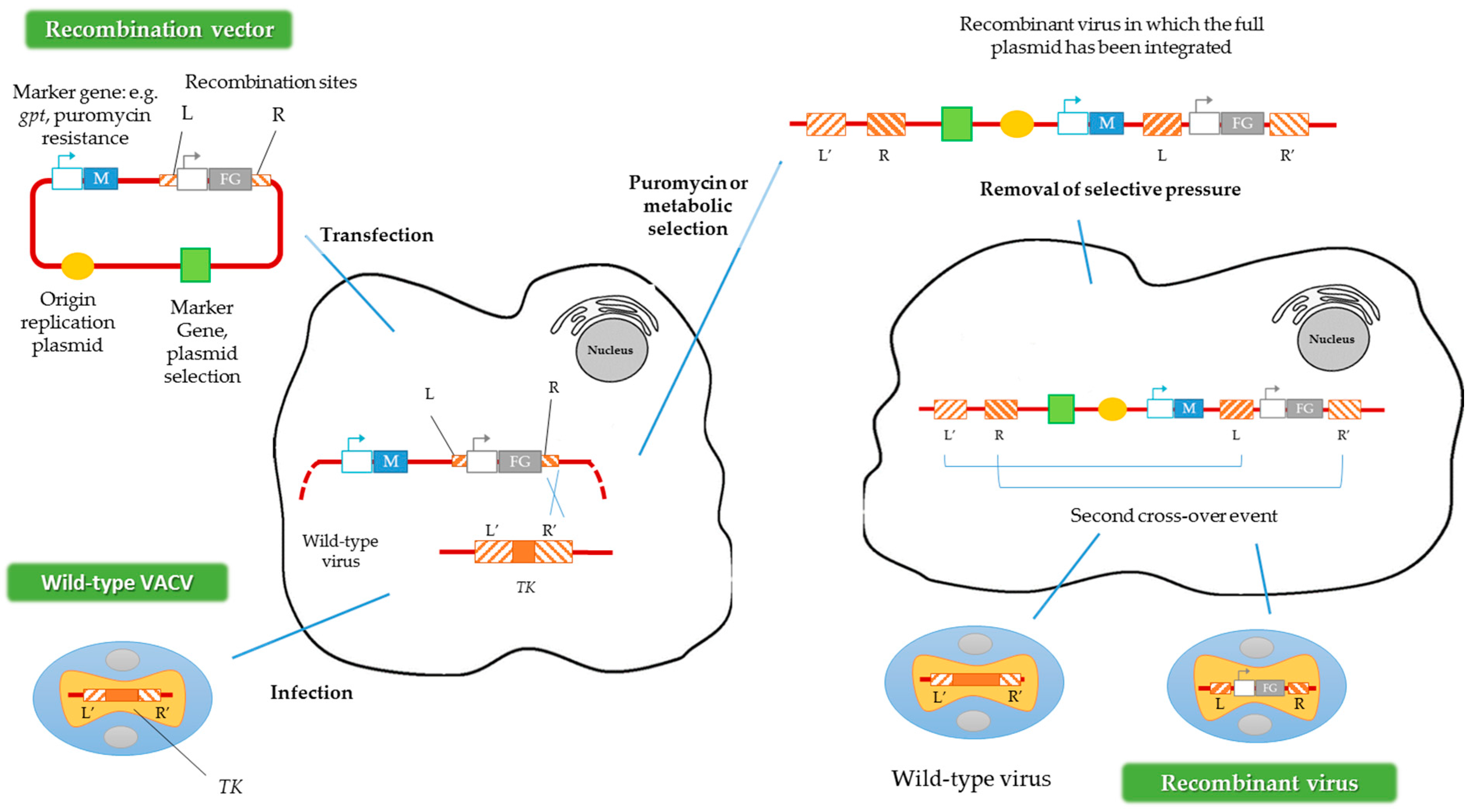
4. Design Considerations in the Generation of VACV Vectors
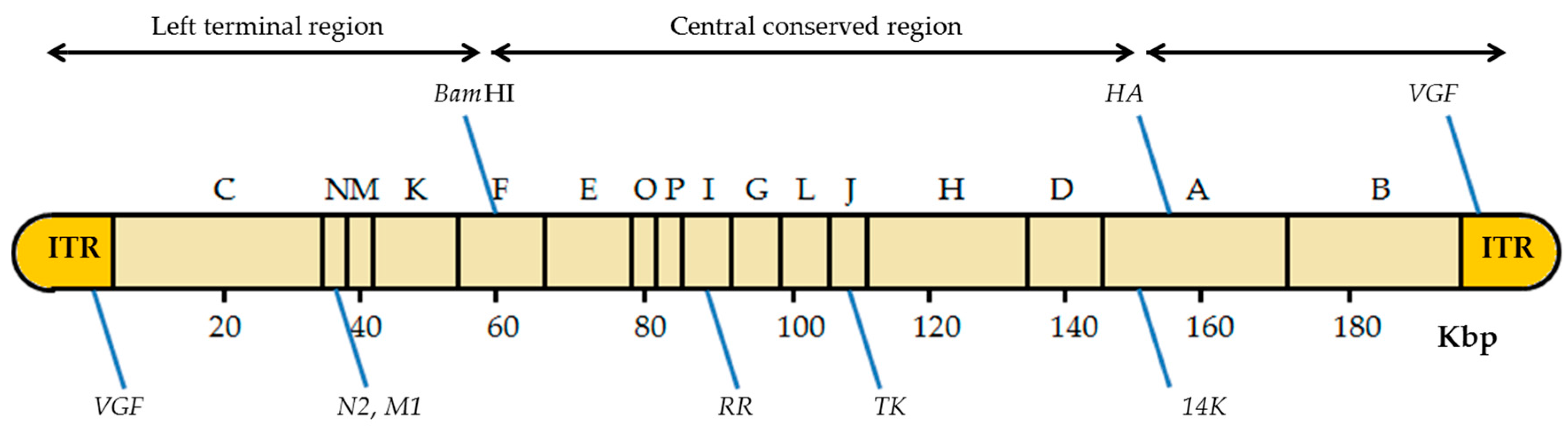
4.1. VACV Genome Insertion Sites
4.2. Reporter-Expressing Viruses
4.2.1. Chloramphenicol Acetyltransferase
4.2.2. β-Galactosidase
4.2.3. β-Glucuronidase
4.2.4. Florescent Proteins
4.2.5. Luciferases
5. Applications of Reporter-Expressing Viruses
5.1. In Vitro Applications
5.2. In Vivo Applications
6. Limitations of VACV Vectors
7. New Perspectives
8. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| acetyl-CoA | acetyl-coenzyme A |
| ATP | adenosine triphosphate |
| CAT | chloramphenicol acetyltransferase |
| CCD | charge-coupled device |
| CD | cytosine deaminase |
| CRISPR/Cas9 | clustered regularly interspaced short palindromic repeats/CRISPR-associated protein9 |
| Env | envelope |
| FMNH2 | flavin mononucleotide |
| gB | glycoprotein B |
| gD | glycoprotein D |
| GFP | green fluorescent protein |
| gG | glycoprotein G |
| GM-CSF | granulocyte-macrophage colony-stimulating factor |
| gpt | guanine phosphoribosyltransferase gene |
| GUS | β-glucuronidase |
| HA | hemagglutinin |
| HBsAg | hepatitis B virus surface antigen |
| HBV | hepatitis B virus |
| hEPO | human erythropoietin |
| HIV | human immunodeficiency virus |
| hNIS | sodium iodide symporter |
| HSV | herpes simplex virus |
| IPTG | isopropyl beta-D-thiogalactopyranoside |
| ITRs | inverted terminal repeats |
| LACK | Leishmania homolog of activated C kinase |
| lacZ | β-galactosidase gene |
| LS | large surface protein |
| LUC | luciferase |
| M1 | matrix 1 |
| MS | middle surface protein |
| MUG | 4-methylumbelliferyl beta-D-glucuronide |
| NF-kB | nuclear factor-kappa B |
| NP | nucleoprotein |
| NS1 | non-structural protein 1 |
| PA | polymerase acidic |
| PB1 | polymerase basic 1 |
| RT | retrotranscriptase |
| ONPG | ortho-nitrophenyl beta-galactoside |
| RFP | red fluorescent protein |
| RR | ribonucleotide reductase |
| SARS-CoV | severe acute respiratory syndrome-associated coronavirus |
| S-Gal | 3,4-cyclohexenoesculetin beta-D-galactopyranoside |
| SR | somatostatin receptor |
| TDS | transient dominant selection |
| TK | thymidine kinase |
| TK- | TK-defective phenotype |
| VACV | vaccinia virus |
| VGF | vaccinia growth factor |
| WR | Western Reserve strain |
| X-Gal | 5-bromo-4-chloro-3-indolyl beta-D-galactopyranoside |
| X-Gluc | 5-bromo-4-chloro-3-indolyl beta-D-glucuronide |
| YFP | yellow fluorescent protein. |
References
- Wells, K.D. Genetic engineering of mammals. Cell. Tissue Res. 2016, 363, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Gill, R.; Tuteja, R.; Tuteja, N. Genetic engineering of crops: A ray of hope for enhanced food security. Plant. Signal. Behav. 2014, 9, e28545. [Google Scholar] [CrossRef] [PubMed]
- Van Pijkeren, J.P.; Britton, R.A. Precision genome engineering in lactic acid bacteria. Microb. Cell. Fact. 2014, 13, S10. [Google Scholar] [CrossRef] [PubMed]
- Ura, T.; Okuda, K.; Shimada, M. Developments in viral vector-based vaccines. Vaccines 2014, 2, 624–641. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.P.; Haut, L.; Reyes-Sandoval, A.; Pinto, A.R. Recombinant viruses as vaccines against viral diseases. Braz. J. Med. Biol. Res. 2005, 38, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Eberwine, J.H. Mammalian cell transfection: The present and the future. Anal. Bioanal. Chem. 2010, 397, 3173–3178. [Google Scholar] [CrossRef] [PubMed]
- Nakano, E.; Panicali, D.; Paoletti, E. Molecular genetics of vaccinia virus: Demonstration of marker rescue. Proc. Natl. Acad. Sci. USA 1982, 79, 1593–1596. [Google Scholar] [CrossRef] [PubMed]
- Panicali, D.; Paoletti, E. Construction of poxviruses as cloning vectors: Insertion of the thymidine kinase gene from herpes simplex virus into the DNA of infectious vaccinia virus. Proc. Natl. Acad. Sci. USA 1982, 79, 4927–4931. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Sampedro, L.; Perdiguero, B.; Mejias-Perez, E.; Garcia-Arriaza, J.; Di Pilato, M.; Esteban, M. The evolution of poxvirus vaccines. Viruses 2015, 7, 1726–1803. [Google Scholar] [CrossRef] [PubMed]
- McFadden, G. Poxvirus tropism. Nat. Rev. Microbiol. 2005, 3, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S. The world health organization and global smallpox eradication. J. Epidemiol. Community Health 2008, 62, 909–912. [Google Scholar] [CrossRef] [PubMed]
- Broyles, S.S. Vaccinia virus transcription. J. Gen. Virol. 2003, 84, 2293–2303. [Google Scholar] [CrossRef] [PubMed]
- Hruby, D.E. Vaccinia virus vectors: New strategies for producing recombinant vaccines. Clin. Microbiol. Rev. 1990, 3, 153–170. [Google Scholar] [PubMed]
- Hughes, A.L.; Friedman, R. Poxvirus genome evolution by gene gain and loss. Mol. Phylogenet. Evol. 2005, 35, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Moss, B. Poxvirus cell entry: How many proteins does it take? Viruses 2012, 4, 688–707. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, F.I.; Bleck, C.K.; Mercer, J. Poxvirus host cell entry. Curr. Opin. Virol. 2012, 2, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Rubins, K.H.; Hensley, L.E.; Bell, G.W.; Wang, C.; Lefkowitz, E.J.; Brown, P.O.; Relman, D.A. Comparative analysis of viral gene expression programs during poxvirus infection: A transcriptional map of the vaccinia and monkeypox genomes. PLoS ONE 2008, 3, e2628. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.L.; Smith, G.L. Vaccinia virus morphogenesis and dissemination. Trends Microbiol. 2008, 16, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Yen, J.; Golan, R.; Rubins, K. Vaccinia virus infection & temporal analysis of virus gene expression: Part 1. J. Vis. Exp. 2009, 26, 1168. [Google Scholar]
- Bleckwenn, N.A.; Bentley, W.E.; Shiloach, J. Exploring vaccinia virus as a tool for large-scale recombinant protein expression. Biotechnol. Prog. 2003, 19, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Bleckwenn, N.A.; Bentley, W.E.; Shiloach, J. Evaluation of production parameters with the vaccinia virus expression system using microcarrier attached HeLa cells. Biotechnol. Prog. 2005, 21, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Masternak, K.; Wittek, R. Cis- and trans-acting elements involved in reactivation of vaccinia virus early transcription. J. Virol. 1996, 70, 8737–8746. [Google Scholar] [PubMed]
- Nascimento, I.P.; Leite, L.C. Recombinant vaccines and the development of new vaccine strategies. Braz. J. Med. Biol. Res. 2012, 45, 1102–1111. [Google Scholar] [CrossRef] [PubMed]
- Bleckwenn, N.A.; Golding, H.; Bentley, W.E.; Shiloach, J. Production of recombinant proteins by vaccinia virus in a microcarrier based mammalian cell perfusion bioreactor. Biotechnol. Bioeng. 2005, 90, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Coupar, B.E.; Oke, P.G.; Andrew, M.E. Insertion sites for recombinant vaccinia virus construction: Effects on expression of a foreign protein. J. Gen. Virol. 2000, 81, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Kim, M. Replicating poxviruses for human cancer therapy. J. Microbiol. 2015, 53, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Garcel, A.; Fauquette, W.; Dehouck, M.P.; Crance, J.M.; Favier, A.L. Vaccinia virus-induced smallpox postvaccinal encephalitis in case of blood-brain barrier damage. Vaccine 2012, 30, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Henderson, D.A.; Moss, B. Recombinant vaccinia virus vaccines, 3rd ed.; Saunders: Philadelphia, 1999. [Google Scholar]
- Moore, A.R.; Dong, B.; Chen, L.; Xiao, W. Vaccinia virus as a subhelper for AAV replication and packaging. Mol. Ther. Methods Clin. Dev. 2015, 2, 15044. [Google Scholar] [CrossRef] [PubMed]
- Falkner, F.G.; Moss, B. Transient dominant selection of recombinant vaccinia viruses. J. Virol. 1990, 64, 3108–3111. [Google Scholar] [PubMed]
- Falkner, F.G.; Moss, B. Escherichia coli gpt gene provides dominant selection for vaccinia virus open reading frame expression vectors. J. Virol. 1988, 62, 1849–1854. [Google Scholar] [PubMed]
- Marzook, N.B.; Procter, D.J.; Lynn, H.; Yamamoto, Y.; Horsington, J.; Newsome, T.P. Methodology for the efficient generation of fluorescently tagged vaccinia virus proteins. J. Vis. Exp. 2014, e51151. [Google Scholar] [CrossRef] [PubMed]
- Kochneva, G.; Zonov, E.; Grazhdantseva, A.; Yunusova, A.; Sibolobova, G.; Popov, E.; Taranov, O.; Netesov, S.; Chumakov, P.; Ryabchikova, E. Apoptin enhances the oncolytic properties of vaccinia virus and modifies mechanisms of tumor regression. Oncotarget 2014, 5, 11269–11282. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, D.; Gonzalez-Aseguinolaza, G.; Rodriguez, J.R.; Vijayan, A.; Gherardi, M.; Rueda, P.; Casal, J.I.; Esteban, M. Vaccine efficacy against malaria by the combination of porcine parvovirus-like particles and vaccinia virus vectors expressing CS of Plasmodium. PLoS ONE 2012, 7, e34445. [Google Scholar] [CrossRef] [PubMed]
- Gomez, C.E.; Rodriguez, D.; Rodriguez, J.R.; Abaitua, F.; Duarte, C.; Esteban, M. Enhanced CD8+ T cell immune response against a V3 loop multi-epitope polypeptide (TAB13) of HIV-1 Env after priming with purified fusion protein and booster with modified vaccinia virus ankara (MVA-TAB) recombinant: A comparison of humoral and cellular immune responses with the vaccinia virus western reserve (WR) vector. Vaccine 2001, 20, 961–971. [Google Scholar] [PubMed]
- Brown, C.K.; Turner, P.C.; Moyer, R.W. Molecular characterization of the vaccinia virus hemagglutinin gene. J. Virol. 1991, 65, 3598–3606. [Google Scholar] [PubMed]
- O’Brien, T.C.; Tauraso, N.M. Vaccinia virus: Kinetics of the hemagglutination-inhibition test and preparation of hemagglutinin. Arch. Gesamte. Virusforsch. 1972, 36, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, E.; Lipinskas, B.R.; Samsonoff, C.; Mercer, S.; Panicali, D. Construction of live vaccines using genetically engineered poxviruses: Biological activity of vaccinia virus recombinants expressing the hepatitis B virus surface antigen and the herpes simplex virus glycoprotein D. Proc. Natl. Acad. Sci. USA 1984, 81, 193–197. [Google Scholar] [PubMed]
- Buller, R.M.; Chakrabarti, S.; Cooper, J.A.; Twardzik, D.R.; Moss, B. Deletion of the vaccinia virus growth factor gene reduces virus virulence. J. Virol. 1988, 62, 866–874. [Google Scholar] [PubMed]
- Buller, R.M.; Smith, G.L.; Cremer, K.; Notkins, A.L.; Moss, B. Decreased virulence of recombinant vaccinia virus expression vectors is associated with a thymidine kinase-negative phenotype. Nature 1985, 317, 813–815. [Google Scholar] [CrossRef] [PubMed]
- Child, S.J.; Palumbo, G.J.; Buller, R.M.; Hruby, D.E. Insertional inactivation of the large subunit of ribonucleotide reductase encoded by vaccinia virus is associated with reduced virulence in vivo. Virology 1990, 174, 625–629. [Google Scholar] [CrossRef]
- Rodriguez, D.; Rodriguez, J.R.; Rodriguez, J.F.; Trauber, D.; Esteban, M. Highly attenuated vaccinia virus mutants for the generation of safe recombinant viruses. Proc. Natl. Acad. Sci. USA 1989, 86, 1287–1291. [Google Scholar] [CrossRef] [PubMed]
- Condit, R.C.; Motyczka, A.; Spizz, G. Isolation, characterization, and physical mapping of temperature-sensitive mutants of vaccinia virus. Virology 1983, 128, 429–443. [Google Scholar] [CrossRef]
- Jiang, T.; Xing, B.; Rao, J. Recent developments of biological reporter technology for detecting gene expression. Biotechnol. Genet. Eng. Rev. 2008, 25, 41–75. [Google Scholar] [CrossRef] [PubMed]
- Rice, A.D.; Gray, S.A.; Li, Y.; Damon, I.; Moyer, R.W. An efficient method for generating poxvirus recombinants in the absence of selection. Viruses 2011, 3, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Gao, X.; Chard, L.S.; Ali, Z.; Ahmed, J.; Li, Y.; Liu, P.; Lemoine, N.R.; Wang, Y. A marker-free system for highly efficient construction of vaccinia virus vectors using CRISPR Cas9. Mol. Ther. Methods Clin. Dev. 2015, 2, 15035. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Zhang, W.; Wang, J.; Al Yaghchi, C.; Ahmed, J.; Chard, L.; Lemoine, N.R.; Wang, Y. Efficiently editing the vaccinia virus genome by using the CRISPR-Cas9 system. J. Virol. 2015, 89, 5176–5179. [Google Scholar] [CrossRef] [PubMed]
- Falzarano, D.; Groseth, A.; Hoenen, T. Development and application of reporter-expressing mononegaviruses: Current challenges and perspectives. Antiviral Res. 2014, 103, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Ghim, C.M.; Lee, S.K.; Takayama, S.; Mitchell, R.J. The art of reporter proteins in science: Past, present and future applications. BMB Rep. 2010, 43, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Swenson, E.S.; Price, J.G.; Brazelton, T.; Krause, D.S. Limitations of green fluorescent protein as a cell lineage marker. Stem Cells 2007, 25, 2593–2600. [Google Scholar] [CrossRef] [PubMed]
- Gorman, C.M.; Moffat, L.F.; Howard, B.H. Recombinant genomes which express chloramphenicol acetyltransferase in mammalian cells. Mol. Cell. Biol. 1982, 2, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Overbeek, P.A.; Lai, S.P.; Van Quill, K.R.; Westphal, H. Tissue-specific expression in transgenic mice of a fused gene containing RSV terminal sequences. Science 1986, 231, 1574–1577. [Google Scholar] [CrossRef] [PubMed]
- Juers, D.H.; Matthews, B.W.; Huber, R.E. LacZ beta-galactosidase: Structure and function of an enzyme of historical and molecular biological importance. Protein Sci. 2012, 21, 1792–1807. [Google Scholar] [CrossRef] [PubMed]
- Carroll, M.W.; Moss, B. E. coli beta-glucuronidase (GUS) as a marker for recombinant vaccinia viruses. BioTechniques 1995, 19, 352–354, 356. [Google Scholar] [PubMed]
- Bleckwenn, N.A.; Bentley, W.E.; Shiloach, J. Vaccinia virus-based expression of gp120 and eGFP: Survey of mammalian host cell lines. Biotechnol. Prog. 2005, 21, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Gahan, C.G. The bacterial lux reporter system: Applications in bacterial localisation studies. Curr. Gene Ther. 2012, 12, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Gould, S.J.; Subramani, S. Firefly luciferase as a tool in molecular and cell biology. Anal. Biochem. 1988, 175, 5–13. [Google Scholar] [CrossRef]
- Kirby, J.; Heath, P.R.; Shaw, P.J.; Hamdy, F.C. Gene expression assays. Adv. Clin. Chem. 2007, 44, 247–292. [Google Scholar] [PubMed]
- Cui, W.; Liu, L.; Kodibagkar, V.D.; Mason, R.P. S-Gal, a novel 1h MRI reporter for beta-galactosidase. Magn. Reson. Med. 2010, 64, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Brechling, K.; Moss, B. Vaccinia virus expression vector: Coexpression of beta-galactosidase provides visual screening of recombinant virus plaques. Mol. Cell. Biol. 1985, 5, 3403–3409. [Google Scholar] [PubMed]
- Villari, P.; Iannuzzo, M.; Torre, I. An evaluation of the use of 4-methylumbelliferyl-beta-d-glucuronide (MUG) in different solid media for the detection and enumeration of escherichia coli in foods. Lett. Appl. Microbiol. 1997, 24, 286–290. [Google Scholar] [CrossRef]
- Howley, P.M.; Spehner, D.; Drillien, R. A vaccinia virus transfer vector using a GUS reporter gene inserted into the I4L locus. Gene 1996, 172, 233–237. [Google Scholar] [CrossRef]
- Dvoracek, B.; Shors, T. Construction of a novel set of transfer vectors to study vaccinia virus replication and foreign gene expression. Plasmid 2003, 49, 9–17. [Google Scholar] [CrossRef]
- Tsuji, F.I. Early history, discovery, and expression of aequorea green fluorescent protein, with a note on an unfinished experiment. Microsc. Res. Tech. 2010, 73, 785–796. [Google Scholar] [PubMed]
- The Nobel Prize in Chemistry 2008. Available online: http://www.nobelprize.org/nobel_prizes/chemistry/laureates/2008/ (accessed on 19 May 2016).
- Coralli, C.; Cemazar, M.; Kanthou, C.; Tozer, G.M.; Dachs, G.U. Limitations of the reporter green fluorescent protein under simulated tumor conditions. Cancer Res. 2001, 61, 4784–4790. [Google Scholar] [PubMed]
- Prescher, J.A.; Contag, C.H. Guided by the light: Visualizing biomolecular processes in living animals with bioluminescence. Curr. Opin. Chem. Biol. 2010, 14, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Liu, Q.; Huang, W.; Yu, Y.; Wang, Y. Comparison of the replication characteristics of vaccinia virus strains Guang 9 and Tian Tan in vivo and in vitro. Arch. Virol. 2014, 159, 2587–2596. [Google Scholar] [CrossRef] [PubMed]
- Zaitseva, M.; Kapnick, S.M.; Meseda, C.A.; Shotwell, E.; King, L.R.; Manischewitz, J.; Scott, J.; Kodihalli, S.; Merchlinsky, M.; Nielsen, H.; et al. Passive immunotherapies protect WRvFire and IHD-J-Luc vaccinia virus-infected mice from lethality by reducing viral loads in the upper respiratory tract and internal organs. J. Virol. 2011, 85, 9147–9158. [Google Scholar] [CrossRef] [PubMed]
- Tsoneva, D.; Stritzker, J.; Bedenk, K.; Zhang, Q.; Frentzen, A.; Cappello, J.; Fischer, U.; Szalay, A.A. Drug-encoded biomarkers for monitoring biological therapies. PLoS ONE 2015, 10, e0137573. [Google Scholar] [CrossRef] [PubMed]
- Manischewitz, J.; King, L.R.; Bleckwenn, N.A.; Shiloach, J.; Taffs, R.; Merchlinsky, M.; Eller, N.; Mikolajczyk, M.G.; Clanton, D.J.; Monath, T.; et al. Development of a novel vaccinia-neutralization assay based on reporter-gene expression. J. Infect. Dis. 2003, 188, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sheng, Y.; Chu, Y.; Ji, H.; Jiang, S.; Lan, T.; Li, M.; Chen, S.; Fan, Y.; Li, W.; et al. Seven major genomic deletions of vaccinia virus Tiantan strain are sufficient to decrease pathogenicity. Antiviral Res. 2016, 129, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.C.; Moyer, R.W. The vaccinia virus fusion inhibitor proteins SPI-3 (K2) and HA (A56) expressed by infected cells reduce the entry of superinfecting virus. Virology 2008, 380, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Ward, B.M. Visualization and characterization of the intracellular movement of vaccinia virus intracellular mature virions. J. Virol. 2005, 79, 4755–4763. [Google Scholar] [CrossRef] [PubMed]
- Warren, R.D.; Cotter, C.A.; Moss, B. Reverse genetics analysis of poxvirus intermediate transcription factors. J. Virol. 2012, 86, 9514–9519. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Lei, R.; Le Duff, Y.; Li, J.; Guo, F.; Wainberg, M.A.; Liang, C. The CRISPR/Cas9 system inactivates latent HIV-1 proviral DNA. Retrovirology 2015, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.M.; Ward, B.M. The A33-dependent incorporation of B5 into extracellular enveloped vaccinia virions is mediated through an interaction between their lumenal domains. J. Virol. 2012, 86, 8210–8220. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.L.; Benfield, C.T.; Maluquer de Motes, C.; Mazzon, M.; Ember, S.W.; Ferguson, B.J.; Sumner, R.P. Vaccinia virus immune evasion: Mechanisms, virulence and immunogenicity. J. Gen. Virol. 2013, 94, 2367–2392. [Google Scholar] [CrossRef] [PubMed]
- Unterholzner, L.; Sumner, R.P.; Baran, M.; Ren, H.; Mansur, D.S.; Bourke, N.M.; Randow, F.; Smith, G.L.; Bowie, A.G. Vaccinia virus protein C6 is a virulence factor that binds tbk-1 adaptor proteins and inhibits activation of IRF3 and IRF7. PLoS Pathog. 2011, 7, e1002247. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.C.; Damon, I.K.; Karem, K.L. A rapid, high-throughput vaccinia virus neutralization assay for testing smallpox vaccine efficacy based on detection of green fluorescent protein. J. Virol. Methods 2008, 150, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, R.; Deng, Y.; Lu, N.; Chen, H.; Meng, X.; Wang, W.; Wang, X.; Yan, K.; Qi, X.; et al. Protective efficacy of the conserved NP, PB1, and M1 proteins as immunogens in DNA- and vaccinia virus-based universal influenza a virus vaccines in mice. Clin. Vaccine Immunol. 2015, 22, 618–630. [Google Scholar] [CrossRef] [PubMed]
- Van den Worm, S.H.; Eriksson, K.K.; Zevenhoven, J.C.; Weber, F.; Zust, R.; Kuri, T.; Dijkman, R.; Chang, G.; Siddell, S.G.; Snijder, E.J.; et al. Reverse genetics of SARS-related coronavirus using vaccinia virus-based recombination. PLoS ONE 2012, 7, e32857. [Google Scholar] [CrossRef] [PubMed]
- Harrington, L.E.; Most Rv, R.; Whitton, J.L.; Ahmed, R. Recombinant vaccinia virus-induced T-cell immunity: Quantitation of the response to the virus vector and the foreign epitope. J. Virol. 2002, 76, 3329–3337. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Ren, X.; Ben, Y.; Ren, Y.; Wang, J.; Zhang, X.; Wan, Y.; Xu, J. Fusion-expressed ctb improves both systemic and mucosal T-cell responses elicited by an intranasal DNA priming/intramuscular recombinant vaccinia boosting regimen. J. Immunol. Res. 2014, 2014, 308732. [Google Scholar] [CrossRef] [PubMed]
- Yaghchi, C.A.; Zhang, Z.; Alusi, G.; Lemoine, N.R.; Wang, Y. Vaccinia virus, a promising new therapeutic agent for pancreatic cancer. Immunotherapy 2015, 7, 1249–1258. [Google Scholar] [CrossRef] [PubMed]
- Vemulapalli, R.; Cravero, S.; Calvert, C.L.; Toth, T.E.; Sriranganathan, N.; Boyle, S.M.; Rossetti, O.L.; Schurig, G.G. Characterization of specific immune responses of mice inoculated with recombinant vaccinia virus expressing an 18-kilodalton outer membrane protein of brucella abortus. Clin. Diagn. Lab. Immunol. 2000, 7, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Langford, C.J.; Edwards, S.J.; Smith, G.L.; Mitchell, G.F.; Moss, B.; Kemp, D.J.; Anders, R.F. Anchoring a secreted plasmodium antigen on the surface of recombinant vaccinia virus-infected cells increases its immunogenicity. Mol. Cell. Biol. 1986, 6, 3191–3199. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.L.; Godson, G.N.; Nussenzweig, V.; Nussenzweig, R.S.; Barnwell, J.; Moss, B. Plasmodium knowlesi sporozoite antigen: Expression by infectious recombinant vaccinia virus. Science 1984, 224, 397–399. [Google Scholar] [CrossRef] [PubMed]
- Gherardi, M.M.; Ramirez, J.C.; Rodriguez, D.; Rodriguez, J.R.; Sano, G.; Zavala, F.; Esteban, M. IL-12 delivery from recombinant vaccinia virus attenuates the vector and enhances the cellular immune response against HIV-1 Env in a dose-dependent manner. J. Immunol. 1999, 162, 6724–6733. [Google Scholar] [PubMed]
- Valkenburg, S.A.; Li, O.T.; Mak, P.W.; Mok, C.K.; Nicholls, J.M.; Guan, Y.; Waldmann, T.A.; Peiris, J.S.; Perera, L.P.; Poon, L.L. IL-15 adjuvanted multivalent vaccinia-based universal influenza vaccine requires CD4+ T cells for heterosubtypic protection. Proc. Natl. Acad. Sci. USA 2014, 111, 5676–5681. [Google Scholar] [CrossRef] [PubMed]
- Brochier, B.; Aubert, M.F.; Pastoret, P.P.; Masson, E.; Schon, J.; Lombard, M.; Chappuis, G.; Languet, B.; Desmettre, P. Field use of a vaccinia-rabies recombinant vaccine for the control of sylvatic rabies in europe and North America. Rev. Sci. Tech. 1996, 15, 947–970. [Google Scholar] [CrossRef] [PubMed]
- Gherardi, M.M.; Najera, J.L.; Perez-Jimenez, E.; Guerra, S.; Garcia-Sastre, A.; Esteban, M. Prime-boost immunization schedules based on influenza virus and vaccinia virus vectors potentiate cellular immune responses against human immunodeficiency virus Env protein systemically and in the genitorectal draining lymph nodes. J. Virol. 2003, 77, 7048–7057. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.D.; Flexner, C.; Paradis, T.J.; Fuller, T.C.; Hirsch, M.S.; Schooley, R.T.; Moss, B. HIV-1 reverse transcriptase is a target for cytotoxic T lymphocytes in infected individuals. Science 1988, 240, 64–66. [Google Scholar] [CrossRef] [PubMed]
- Cheliapov, N.V.; Chernos, V.I.; Andzhaparidze, O.G. Analysis of antibody formation to the vaccinia virus in human subjects and rabbits in response to the administration of a recombinant vaccinia-hepatitis B vaccine. Vopr. Virusol. 1988, 33, 175–179. [Google Scholar] [PubMed]
- Grigorieva, I.M.; Grigoriev, V.G.; Zakharova, L.G.; Pashvykina, G.V.; Shevlyagin, V.Y.; Altstein, A.D. Immunogenicity of recombinant vaccinia viruses expressing hepatitis B virus surface antigen in mice. Immunol. Lett. 1993, 36, 267–271. [Google Scholar] [CrossRef]
- Kutinova, L.; Ludvikova, V.; Krystofova, J.; Otavova, M.; Simonova, V.; Nemeckova, S.; Hainz, P.; Vonka, V. Influence of the parental virus strain on the virulence and immunogenicity of recombinant vaccinia viruses expressing HBV Pres2-S protein or VZV glycoprotein I. Vaccine 1996, 14, 1045–1052. [Google Scholar] [CrossRef]
- Cheng, K.C.; Smith, G.L.; Moss, B. Hepatitis B virus large surface protein is not secreted but is immunogenic when selectively expressed by recombinant vaccinia virus. J. Virol. 1986, 60, 337–344. [Google Scholar] [PubMed]
- Cheng, K.C.; Moss, B. Selective synthesis and secretion of particles composed of the hepatitis B virus middle surface protein directed by a recombinant vaccinia virus: Induction of antibodies to pre-S and S epitopes. J. Virol. 1987, 61, 1286–1290. [Google Scholar] [PubMed]
- Rooney, J.F.; Wohlenberg, C.; Cremer, K.J.; Moss, B.; Notkins, A.L. Immunization with a vaccinia virus recombinant expressing herpes simplex virus type 1 glycoprotein D: Long-term protection and effect of revaccination. J. Virol. 1988, 62, 1530–1534. [Google Scholar] [PubMed]
- Martin, S.; Moss, B.; Berman, P.W.; Laskey, L.A.; Rouse, B.T. Mechanisms of antiviral immunity induced by a vaccinia virus recombinant expressing herpes simplex virus type 1 glycoprotein D: Cytotoxic t cells. J. Virol. 1987, 61, 726–734. [Google Scholar] [PubMed]
- Cantin, E.M.; Eberle, R.; Baldick, J.L.; Moss, B.; Willey, D.E.; Notkins, A.L.; Openshaw, H. Expression of herpes simplex virus 1 glycoprotein B by a recombinant vaccinia virus and protection of mice against lethal herpes simplex virus 1 infection. Proc. Natl. Acad. Sci. USA 1987, 84, 5908–5912. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin-Taylor, E.; Willey, D.E.; Cantin, E.M.; Eberle, R.; Moss, B.; Openshaw, H. A recombinant vaccinia virus expressing herpes simplex virus type 1 glycoprotein B induces cytotoxic T lymphocytes in mice. J. Gen. Virol. 1988, 69(Pt. 7), 1731–1734. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, V.; Smith, G.L. Expression and characterization of herpes simplex virus type 1 (HSV-1) glycoprotein G (gG) by recombinant vaccinia virus: Neutralization of HSV-1 infectivity with anti-gG antibody. J. Gen. Virol. 1987, 68(Pt. 10), 2587–2598. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.L.; Murphy, B.R.; Moss, B. Construction and characterization of an infectious vaccinia virus recombinant that expresses the influenza hemagglutinin gene and induces resistance to influenza virus infection in hamsters. Proc. Natl. Acad. Sci. USA 1983, 80, 7155–7159. [Google Scholar] [CrossRef] [PubMed]
- Goodman, A.G.; Heinen, P.P.; Guerra, S.; Vijayan, A.; Sorzano, C.O.; Gomez, C.E.; Esteban, M. A human multi-epitope recombinant vaccinia virus as a universal T cell vaccine candidate against influenza virus. PLoS ONE 2011, 6, e25938. [Google Scholar] [CrossRef] [PubMed]
- Ramos, I.; Alonso, A.; Marcen, J.M.; Peris, A.; Castillo, J.A.; Colmenares, M.; Larraga, V. Heterologous prime-boost vaccination with a non-replicative vaccinia recombinant vector expressing lack confers protection against canine visceral leishmaniasis with a predominant Th1-specific immune response. Vaccine 2008, 26, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Cross, M.L.; Fleming, S.B.; Cowan, P.E.; Scobie, S.; Whelan, E.; Prada, D.; Mercer, A.A.; Duckworth, J.A. Vaccinia virus as a vaccine delivery system for marsupial wildlife. Vaccine 2011, 29, 4537–4543. [Google Scholar] [CrossRef] [PubMed]
- Hruby, D.E.; Hodges, W.M.; Wilson, E.M.; Franke, C.A.; Fischetti, V.A. Expression of streptococcal M protein in mammalian cells. Proc. Natl. Acad. Sci. USA 1988, 85, 5714–5717. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.M.; McFadden, G. Oncolytic poxviruses. Annu. Rev. Virol. 2014, 1, 119–141. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, D.C.; Kyula, J.N.; Rosenfelder, N.; Chao-Chu, J.; Kramer-Marek, G.; Khan, A.A.; Roulstone, V.; McLaughlin, M.; Melcher, A.A.; Vile, R.G.; et al. Oncolytic vaccinia virus as a vector for therapeutic sodium iodide symporter gene therapy in prostate cancer. Gene Ther. 2016, 23, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Pugalenthi, A.; Mojica, K.; Ady, J.W.; Johnsen, C.; Love, D.; Chen, N.G.; Aguilar, R.J.; Szalay, A.A.; Fong, Y. Recombinant vaccinia virus GLV-1h68 is a promising oncolytic vector in the treatment of cholangiocarcinoma. Cancer Gene Ther. 2015, 22, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Breitbach, C.J.; Lee, J.; Park, J.O.; Lim, H.Y.; Kang, W.K.; Moon, A.; Mun, J.H.; Sommermann, E.M.; Maruri Avidal, L.; et al. Phase 1b trial of biweekly intravenous Pexa-Vec (JX-594), an oncolytic and immunotherapeutic vaccinia virus in colorectal cancer. Mol. Ther. 2015, 23, 1532–1540. [Google Scholar] [CrossRef] [PubMed]
- Bauzon, M.; Hermiston, T. Armed therapeutic viruses - a disruptive therapy on the horizon of cancer immunotherapy. Front. Immunol. 2014, 5, 74. [Google Scholar] [CrossRef] [PubMed]
- Merrick, A.E.; Ilett, E.J.; Melcher, A.A. Jx-594, a targeted oncolytic poxvirus for the treatment of cancer. Curr. Opin. Investig. Drugs. 2009, 10, 1372–1382. [Google Scholar] [PubMed]
- McCart, J.A.; Ward, J.M.; Lee, J.; Hu, Y.; Alexander, H.R.; Libutti, S.K.; Moss, B.; Bartlett, D.L. Systemic cancer therapy with a tumor-selective vaccinia virus mutant lacking thymidine kinase and vaccinia growth factor genes. Cancer Res. 2001, 61, 8751–8757. [Google Scholar] [PubMed]
- Lun, X.; Ruan, Y.; Jayanthan, A.; Liu, D.J.; Singh, A.; Trippett, T.; Bell, J.; Forsyth, P.; Johnston, R.N.; Narendran, A. Double-deleted vaccinia virus in virotherapy for refractory and metastatic pediatric solid tumors. Mol. Oncol. 2013, 7, 944–954. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.H.; Chen, N.G.; Zhang, Q.; Le, H.T.; Aguilar, R.J.; Yu, Y.A.; Cappello, J.; Szalay, A.A. Vaccinia virus-mediated expression of human erythropoietin in tumors enhances virotherapy and alleviates cancer-related anemia in mice. Mol. Ther. 2013, 21, 2054–2062. [Google Scholar] [CrossRef] [PubMed]
- McCart, J.A.; Mehta, N.; Scollard, D.; Reilly, R.M.; Carrasquillo, J.A.; Tang, N.; Deng, H.; Miller, M.; Xu, H.; Libutti, S.K.; et al. Oncolytic vaccinia virus expressing the human somatostatin receptor SSTR2: Molecular imaging after systemic delivery using 111in-pentetreotide. Mol. Ther. 2004, 10, 553–561. [Google Scholar] [PubMed]
- Mei, Y.; Wang, Y.; Chen, H.; Sun, Z.S.; Ju, X.D. Recent progress in CRISPR/Cas9 technology. J. Genet. Genomics. 2016, 43, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, T.; Kohyama, M.; Hirayasu, K.; Arase, H. Engineering large viral DNA genomes using the CRISPR-Cas9 system. Microbiol. Immunol. 2014, 58, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Zhang, K.; Li, J. Application of CRISPR/Cas9 technology to HBV. Int. J. Mol. Sci. 2015, 16, 26077–26086. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reporter Gene | Origin | Product | Detection | Reference |
|---|---|---|---|---|
| CAT | Escherichia coli | Chloramphenicol acetyltransferase | Thin-layer chromatography autoradiography, ELISA | [51,52] |
| LacZ | Escherichia coli | β-galactosidase | Colorimetry | [53] |
| GUS | Escherichia coli | β-glucuronidase | Colorimetry or fluorescence | [54] |
| GFP | Aequorea victoria (jellyfish) | Green fluorescent protein | Fluorescence | [50,55] |
| LUC or luxCDABE | Photinus pyralis (firefly) and bacteria | Luciferase | Luminescence | [56,57] |
| Pathogenic Agent | Antigen | Features | Reference | ||
|---|---|---|---|---|---|
| Site of Insertion | Reporter Gene | ||||
| Viral | HIV | Env | TK or HA gene | LacZ, LUC | [90,92] |
| Env (TAB 13) | HA gene | LacZ | [35] | ||
| RT | Not mentioned | LacZ | [93] | ||
| Hepatitis B virus | HBsAg | TK gene or BamHI site | Not mentioned | [38,94,95] | |
| PreS2-S | TK gene | Not mentioned | [96] | ||
| LS | TK gene | Not mentioned | [97] | ||
| MS | TK gene | Not mentioned | [98] | ||
| Herpes simplex virus 1 | gD | TK gene or BamHI site | Not mentioned | [38,99,100] | |
| gB | Not mentioned | Not mentioned | [101,102] | ||
| gG | Not mentioned | Not mentioned | [103] | ||
| Influenza | HA | TK gene | Not mentioned | [104] | |
| M1, NS1, NP, PB1, PA | TK gene | Not mentioned | [105] | ||
| Protist | Plasmodium yoelii | Circumsporozoite | TK gene | LacZ | [34] |
| Plasmodium knowlesi | Sporozoite antigen | TK gene | Not mentioned | [88] | |
| Plasmodium falciparum | S antigen | TK gene | Not mentioned | [87] | |
| Leishmania infantum | LACK | TK and HA gene | LacZ and GUS | [106] | |
| Animal | Echinococcus granulosus | E95 antigen | TK gene | LacZ | [107] |
| Bacterial | Brucella abortus | 18-kDa antigen | TK gene | LacZ | [86] |
| Streptococcus pyogenes | M protein | TK gene | Not mentioned | [108] | |
| Virus | Target Cancer | Features | Reference | |
|---|---|---|---|---|
| Inactive Genes | Additional Genes | |||
| Initial virus | ||||
| JX-594 | Melanoma, hepatocellular carcinoma, colorectal cancer | TK inactive, | LacZ and GM-CSF | [112,114] |
| GLV-1h68 | Colorectal cancer, prostate cancer, salivary gland carcinoma | TK, HA and F14.5L inactive | GFP, LacZ and GUS | [111] |
| vvDD | Sarcomas, neuroblastoma | TK and VGF inactive | CD | [115,116] |
| Enhanced virus | ||||
| GLV-1h153 | Pancreatic cancer | GLV-1h68 expressing hNIS | [110] | |
| GLV-1h210 | Lung cancer | GLV-1h68 expressing hEPO | [117] | |
| vvDD-SR-RFP | Sarcomas, neuroblastoma | TK and VGF inactive | CD, RFP, SR | [118] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Ali, S.; Baldanta, S.; Fernández-Escobar, M.; Guerra, S. Use of Reporter Genes in the Generation of Vaccinia Virus-Derived Vectors. Viruses 2016, 8, 134. https://doi.org/10.3390/v8050134
Al Ali S, Baldanta S, Fernández-Escobar M, Guerra S. Use of Reporter Genes in the Generation of Vaccinia Virus-Derived Vectors. Viruses. 2016; 8(5):134. https://doi.org/10.3390/v8050134
Chicago/Turabian StyleAl Ali, Sally, Sara Baldanta, Mercedes Fernández-Escobar, and Susana Guerra. 2016. "Use of Reporter Genes in the Generation of Vaccinia Virus-Derived Vectors" Viruses 8, no. 5: 134. https://doi.org/10.3390/v8050134
APA StyleAl Ali, S., Baldanta, S., Fernández-Escobar, M., & Guerra, S. (2016). Use of Reporter Genes in the Generation of Vaccinia Virus-Derived Vectors. Viruses, 8(5), 134. https://doi.org/10.3390/v8050134