Human Protoparvoviruses



Abstract
:
1. Introduction
2. Bufavirus
2.1. Discovery and Taxonomy
2.2. PCR-Based Epidemiology
2.3. Antibody-Based Epidemiology
2.4. Bufavirus-Like Animal Viruses
3. Tusavirus
4. Cutavirus
5. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cossart, Y.E.; Field, A.M.; Cant, B.; Widdows, D. Parvovirus-like particles in human sera. Lancet 1975, 1, 72–73. [Google Scholar] [CrossRef]
- Qiu, J.; Soderlund-Venermo, M.; Young, N.S. Human Parvoviruses. Clin. Microbiol. Rev. 2017, 30, 43–113. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.S.; Kapoor, A.; Lukashov, V.V.; Simmonds, P.; Hecht, F.; Delwart, E. New DNA viruses identified in patients with acute viral infection syndrome. J. Virol. 2005, 79, 8230–8236. [Google Scholar] [CrossRef] [PubMed]
- Allander, T.; Tammi, M.T.; Eriksson, M.; Bjerkner, A.; Tiveljung-Lindell, A.; Andersson, B. Cloning of a human parvovirus by molecular screening of respiratory tract samples. Proc. Natl. Acad. Sci. USA 2005, 102, 12891–12896. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.L.; Higgins, G.D.; Davidson, G.P.; Givney, R.C.; Ratcliff, R.M. A novel bocavirus associated with acute gastroenteritis in Australian children. PLoS. Pathog. 2009, 5, e1000391. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, A.; Slikas, E.; Simmonds, P.; Chieochansin, T.; Naeem, A.; Shaukat, S.; Alam, M.M.; Sharif, S.; Angez, M.; Zaidi, S.; et al. A newly identified bocavirus species in human stool. J. Infect. Dis. 2009, 199, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, A.; Simmonds, P.; Slikas, E.; Li, L.; Bodhidatta, L.; Sethabutr, O.; Triki, H.; Bahri, O.; Oderinde, B.S.; Baba, M.M.; et al. Human bocaviruses are highly diverse, dispersed, recombination prone, and prevalent in enteric infections. J. Infect. Dis. 2010, 201, 1633–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, T.G.; Vo, N.P.; Bonkoungou, I.J.; Kapoor, A.; Barro, N.; O’Ryan, M.; Kapusinszky, B.; Wang, C.; Delwart, E. Acute diarrhea in West African children: Diverse enteric viruses and a novel parvovirus genus. J. Virol. 2012, 86, 11024–11030. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.G.; Dreno, B.; da Costa, A.C.; Li, L.; Orlandi, P.; Deng, X.; Kapusinszky, B.; Siqueira, J.; Knol, A.C.; Halary, F.; et al. A new protoparvovirus in human fecal samples and cutaneous T cell lymphomas (mycosis fungoides). Virology 2016, 496, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.G.; Sdiri-Loulizi, K.; Aouni, M.; Ambert-Balay, K.; Pothier, P.; Deng, X.; Delwart, E. New parvovirus in child with unexplained diarrhea, Tunisia. Emerg. Infect. Dis. 2014, 20, 1911–1913. [Google Scholar] [CrossRef] [PubMed]
- Yahiro, T.; Wangchuk, S.; Tshering, K.; Bandhari, P.; Zangmo, S.; Dorji, T.; Tshering, K.; Matsumoto, T.; Nishizono, A.; Soderlund-Venermo, M.; et al. Novel human bufavirus genotype 3 in children with severe diarrhea, Bhutan. Emerg. Infect. Dis. 2014, 20, 1037–1039. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Liu, D.; Wang, Y.; Qu, F.; He, Y.; Sun, Z.; Shen, Q.; Li, W.; Fu, X.; Deng, X.; et al. Bufavirus Protoparvovirus in feces of wild rats in China. Virus Genes 2016, 52, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Cotmore, S.F.; Agbandje-McKenna, M.; Chiorini, J.A.; Mukha, D.V.; Pintel, D.J.; Qiu, J.; Soderlund-Venermo, M.; Tattersall, P.; Tijssen, P.; Gatherer, D.; et al. The family Parvoviridae. Arch. Virol. 2014, 159, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4674–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Vaisanen, E.; Kuisma, I.; Phan, T.G.; Delwart, E.; Lappalainen, M.; Tarkka, E.; Hedman, K.; Soderlund-Venermo, M. Bufavirus in feces of patients with gastroenteritis, Finland. Emerg. Infect. Dis. 2014, 20, 1077–1080. [Google Scholar] [CrossRef] [PubMed]
- Smits, S.L.; Schapendonk, C.M.; van Beek, J.; Vennema, H.; Schurch, A.C.; Schipper, D.; Bodewes, R.; Haagmans, B.L.; Osterhaus, A.D.; Koopmans, M.P. New viruses in idiopathic human diarrhea cases, the Netherlands. Emerg. Infect. Dis. 2014, 20, 1218–1222. [Google Scholar] [CrossRef] [PubMed]
- Altay, A.; Yahiro, T.; Bozdayi, G.; Matsumoto, T.; Sahin, F.; Ozkan, S.; Nishizono, A.; Soderlund-Venermo, M.; Ahmed, K. Bufavirus genotype 3 in Turkish children with severe diarrhoea. Clin. Microbiol. Infect. 2015, 21, e1–e4. [Google Scholar] [CrossRef] [PubMed]
- Chieochansin, T.; Vutithanachot, V.; Theamboonlers, A.; Poovorawan, Y. Bufavirus in fecal specimens of patients with and without diarrhea in Thailand. Arch. Virol. 2015, 160, 1781–1784. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.D.; Wang, W.; Lu, Q.B.; Zhao, J.; Guo, C.T.; Wang, H.Y.; Zhang, X.A.; Tong, Y.G.; Liu, W.; Cao, W.C. Identification of Bufavirus-1 and Bufavirus-3 in Feces of Patients with Acute Diarrhea, China. Sci. Rep. 2015, 5, 13272. [Google Scholar] [CrossRef] [PubMed]
- Ayouni, S.; Estienney, M.; Hammami, S.; Neji Guediche, M.; Pothier, P.; Aouni, M.; Belliot, G.; de Rougemont, A. Cosavirus, Salivirus and Bufavirus in Diarrheal Tunisian Infants. PLoS ONE 2016, 11, e0162255. [Google Scholar] [CrossRef] [PubMed]
- Vaisanen, E.; Paloniemi, M.; Kuisma, I.; Lithovius, V.; Kumar, A.; Franssila, R.; Ahmed, K.; Delwart, E.; Vesikari, T.; Hedman, K.; et al. Epidemiology of two human protoparvoviruses, bufavirus and tusavirus. Sci. Rep. 2016, 6, 39267. [Google Scholar] [CrossRef] [PubMed]
- Altan, E.; Del Valle Mendoza, J.; Deng, X.; Phan, T.G.; Sadeghi, M.; Delwart, E.L. Small Circular Rep-Encoding Single-Stranded DNA Genomes in Peruvian Diarrhea Virome. Genome Announc. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Altay Kocak, A.; Ocal, M.; Polat, M.; Kanik Yuksek, S.; Aktas Tapisiz, A.; Tezer, H.; Ozkul, A.; Ergunay, K.; Bozdayi, G.; Ahmed, K. Multicenter investigation of bufavirus in the etiology of viral central nervous system infections of adults and children. Mikrobiyol. Bul. 2017, 51, 191–194. [Google Scholar] [CrossRef] [PubMed]
- Handley, S.A.; Thackray, L.B.; Zhao, G.; Presti, R.; Miller, A.D.; Droit, L.; Abbink, P.; Maxfield, L.F.; Kambal, A.; Duan, E.; et al. Pathogenic simian immunodeficiency virus infection is associated with expansion of the enteric virome. Cell 2012, 151, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Orba, Y.; Anindita, P.D.; Ishii, A.; Ueno, K.; Hang’ombe, B.M.; Mweene, A.S.; Ito, K.; Sawa, H. Distinct Lineages of Bufavirus in Wild Shrews and Nonhuman Primates. Emerg. Infect. Dis. 2015, 21, 1230–1233. [Google Scholar] [CrossRef] [PubMed]
- Kemenesi, G.; Dallos, B.; Gorfol, T.; Estok, P.; Boldogh, S.; Kurucz, K.; Oldal, M.; Marton, S.; Banyai, K.; Jakab, F. Genetic diversity and recombination within bufaviruses: Detection of a novel strain in Hungarian bats. Infect. Genet. Evol. 2015, 33, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Hargitai, R.; Pankovics, P.; Kertesz, A.M.; Biro, H.; Boros, A.; Phan, T.G.; Delwart, E.; Reuter, G. Detection and genetic characterization of a novel parvovirus distantly related to human bufavirus in domestic pigs. Arch. Virol. 2016, 161, 1033–1037. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Schwarz, L.; Ullman, K.; Ahola, H.; Qiu, Y.; Ma, Z.; Hennig-Pauka, I. Identification of a novel bufavirus in domestic pigs by a viral metagenomic approach. J. Gen. Virol. 2016, 97, 1592–1596. [Google Scholar] [CrossRef] [PubMed]
- Kluge, M.; Campos, F.S.; Tavares, M.; de Amorim, D.B.; Valdez, F.P.; Giongo, A.; Roehe, P.M.; Franco, A.C. Metagenomic Survey of Viral Diversity Obtained from Feces of Subantarctic and South American Fur Seals. PLoS ONE 2016, 11, e0151921. [Google Scholar] [CrossRef] [PubMed]
- Mollerup, S.; Fridholm, H.; Vinner, L.; Kjartansdottir, K.R.; Friis-Nielsen, J.; Asplund, M.; Herrera, J.A.; Steiniche, T.; Mourier, T.; Brunak, S.; et al. Cutavirus in Cutaneous Malignant Melanoma. Emerg. Infect. Dis. 2017, 23, 363–365. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
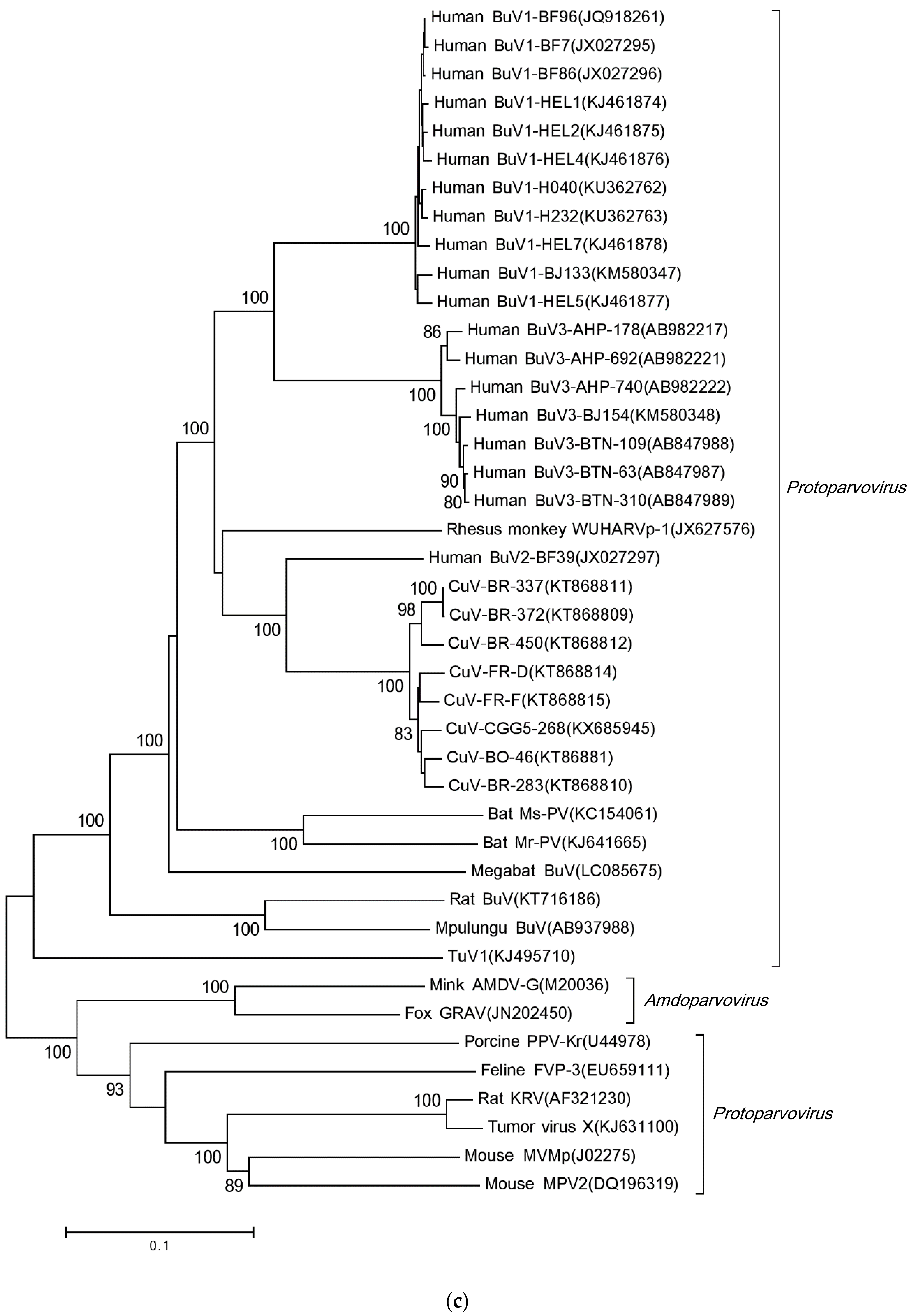{kind=link}
{kind=link}
| Study | Sample Type | Country | Age (Range) If Known | Sampling Time | n | Positive (%) | Symptoms | Genotype |
|---|---|---|---|---|---|---|---|---|
| [8] | Feces | Burkina Faso | <5 yr | November 2008–February 2010 | 98 | 4 (4.1%) | GE | Three BuV1, one BuV2 |
| Feces | Tunisia | “children” | NA | 63 | 1 (1.6%) | NPAFP | unknown | |
| Feces | Chile | “children” | NA | 100 | 0 (0.0%) | GE | - | |
| [11] | Feces | Bhutan | <5 yr | February 2010–January 2012 | 393 | 3 (0.8%) | GE | All BuV3 |
| [16] | Feces | Finland | median 51.5 yr (0–99 yr) | October 2012–March 2013; April–May 2013 | 629 | 7 (1.1%) | GE | Six BuV1, one unknown |
| [17] | Feces | The Netherlands | median 47 yr (0–97 yr) | 2005–2009 | 27 | 1 (3.7%) | GE | BuV3 |
| [19] | Feces | Thailand | median NA (0–97 yr) | January 2009–April 2014 | 1495 | 4 (0.3%) | GE | All BuV1 |
| Feces | Thailand | median NA (0–39 yr) | February 2010–July 2014 | 741 | 0 | HFMD | - | |
| [18] | Feces | Turkey | mean 19.5 mo (1–60 mo) | September 2004–June 2011 | 583 | 8 (1.4%) | GE | Six BuV3, two unknown |
| Feces | Turkey | mean 17.3 mo (NA), age matched | February–September 2013 | 148 | 0 | healthy | - | |
| [20] | Feces | China (General hospital, Beijing) | median 35 yr (1 mo–85 yr) | 2010–2014 | 520 | 9 (1.7%) | GE | Four BuV1, five BuV3 |
| “children” | NA | 76 | 0 | non-GE | - | |||
| Feces | China (Children’s hospital, Chongqing) | median 10 mo (1 day–14 yr) | 2010–2013 | 1357 | 0 | GE | - | |
| “children” | NA | 345 | 0 | non-GE | - | |||
| [21] | Feces | Tunisia | median 7.0 mo (0.5–60 mo) | October 2010–March 2012 | 203 | 2 (1.0%) | GE | Both BuV1 |
| [22] | Feces | Finland | median 14 mo (6 days–15.6 yr) | September 2009–August 2011 | 172 | 2 (1.2%) | GE | Unknown 1 |
| 545 | 0 | ARTI | - | |||||
| 238 | 1 (0.4%) | GE & ARTI | Unknown 1 | |||||
| nasal swab | Finland | median 14 mo (6 days–15.6 yr) | September 2009–August 2011 | 172 | 0 | GE | - | |
| 545 | 0 | ARTI | - | |||||
| 238 | 1 (0.4%) | GE & ARTI | Unknown 1 | |||||
| [24] | CSF | Turkey | median 32 yr (0–96 yr) | October 2011–April 2015 | 126 | 0 | febrile illness and/or CNS infection | - |
| [23] | Feces | Peru | Unknown | NA | 300 | Unknown 2 | GE | BuV3 |
| Genotype | Adults | Children | |||
|---|---|---|---|---|---|
| Asia, n = 12 | Finland, n = 163 | Finland, n = 228 | |||
| Middle East | India | China | |||
| BuV1 | 1 1 | 3 | 1 | 2 2 | |
| BuV2 | 1 1 | 1 | 4 | 4 2 | |
| BuV3 | 1 1 | 2 | |||
| Any BuV | 1 1 | 3 | 1 | 5 | 7 2 |
| Study | Sample Type | Country | n | Positive (%) | Symptoms/Diagnosis |
|---|---|---|---|---|---|
| [9] | Feces | Brazil | 245 | 4 (1.6%) | Diarrhea |
| [9] | Feces | Botswana | 100 | 1 (1.0%) | Diarrhea |
| [9] | Skin | France 1 | NA | 2 | CTCL, Mycosis fungoides |
| [9] | Skin | France | 15 | 2 (13.3%) | CTCL, Mycosis fungoides |
| [9] | Skin | France | 10 | 0 (0%) | Skin carcinoma |
| [9] | Skin | France 1 | NA | 0 | Parapsoriasis |
| [9] | Skin | 2 France; 6 NA | 8 | 0 (0%) | Parapsoriasis |
| [9] | Skin | NA | 8 | 0 (0%) | Eczema or eczematoid dermatitis |
| [9] | Skin | NA | 3 | 0 (0%) | Healthy |
| [31] | Skin | Denmark | 10 | 1 (10%) | Melanoma |
| % Nucleotide Identity | ||||||||
|---|---|---|---|---|---|---|---|---|
| BR-337, Feces [KT868811] | BR-283, Feces [KT868810] | BR-372, Feces [KT868809] | BR-450, Feces [KT868812] | BO-46, Feces [KT868813] | FR-D, Skin [KT868813] | FR-F, Skin [KT868813] | CGG5-268, Skin [KX685945] | |
| BR-337, feces [KT868811] | 100.0 | |||||||
| BR-283, feces [KT868810] | 94.4 | 100.0 | ||||||
| BR-372, feces [KT868809] | 99.5 | 94.6 | 100.0 | |||||
| BR-450, feces [KT868812] | 96.9 | 94.4 | 97.1 | 100.0 | ||||
| BO-46, feces [KT868813] | 93.9 | 96.0 | 94.1 | 93.9 | 100.0 | |||
| FR-D, skin [KT868813] | 93.8 | 96.2 | 93.8 | 93.9 | 95.0 | 100.0 | ||
| FR-F, skin [KT868813] | 94.4 | 96.6 | 94.5 | 94.5 | 95.5 | 95.9 | 100.0 | |
| CGG5-268, skin [KX685945] | 94.1 | 97.1 | 94.2 | 94.2 | 96.0 | 96.1 | 96.7 | 100.0 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Väisänen, E.; Fu, Y.; Hedman, K.; Söderlund-Venermo, M. Human Protoparvoviruses. Viruses 2017, 9, 354. https://doi.org/10.3390/v9110354
Väisänen E, Fu Y, Hedman K, Söderlund-Venermo M. Human Protoparvoviruses. Viruses. 2017; 9(11):354. https://doi.org/10.3390/v9110354
Chicago/Turabian StyleVäisänen, Elina, Yu Fu, Klaus Hedman, and Maria Söderlund-Venermo. 2017. "Human Protoparvoviruses" Viruses 9, no. 11: 354. https://doi.org/10.3390/v9110354
APA StyleVäisänen, E., Fu, Y., Hedman, K., & Söderlund-Venermo, M. (2017). Human Protoparvoviruses. Viruses, 9(11), 354. https://doi.org/10.3390/v9110354