Chikungunya Virus: Pathophysiology, Mechanism, and Modeling



Abstract
:
1. Introduction
1.1. Chikungunya Fever and CHIKV
1.2. History/Origins
1.3. Symptoms
2. Pathophysiology
2.1. Transmission
2.2. Route of Infection
2.3. Innate Immune Response
3. Diagnosis/Treatment
3.1. Diagnosis
3.2. Treatment
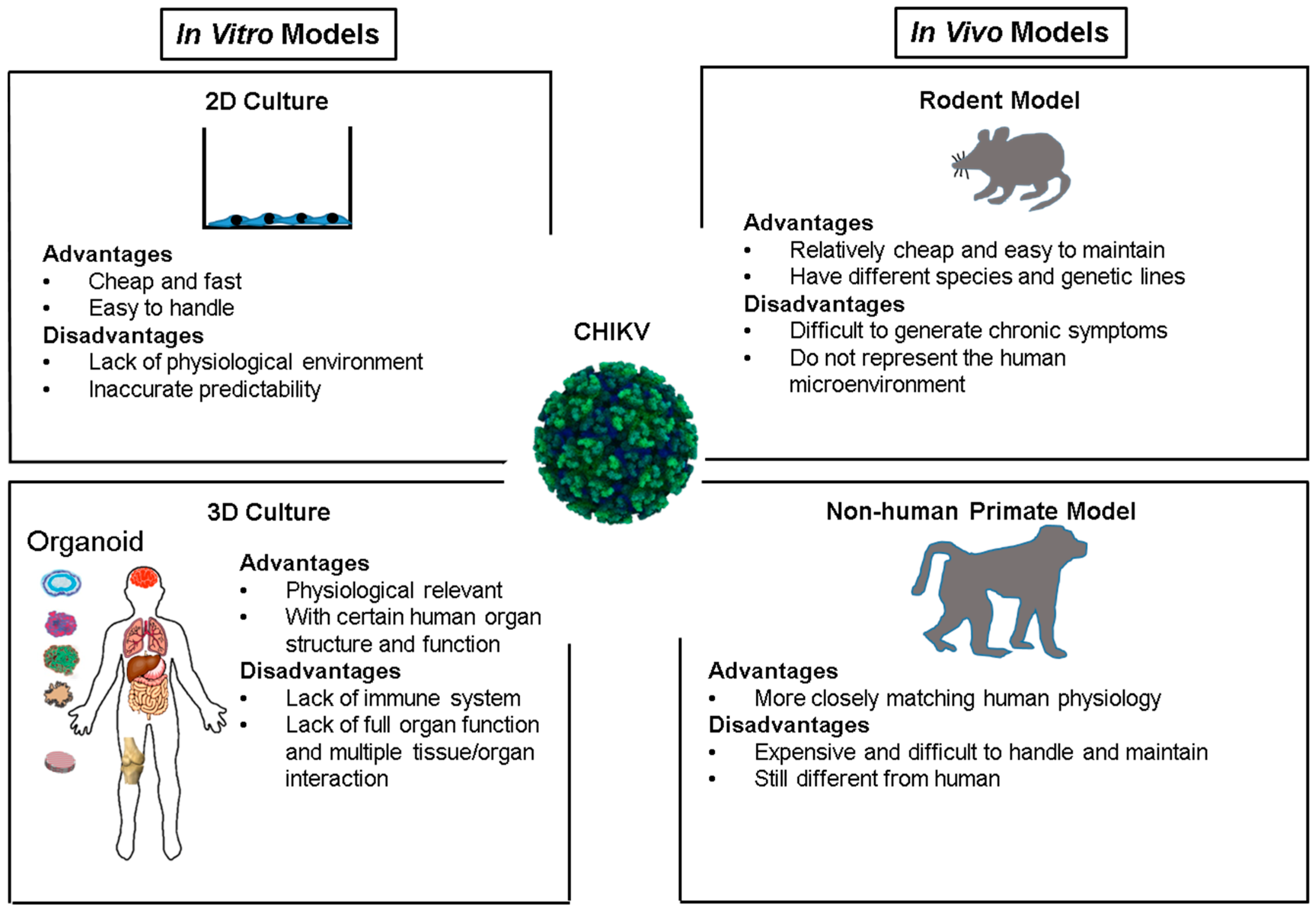
4. CHIKV Infection and Models
4.1. In Vitro Culture System
4.2. Rodent Models
4.3. Non-Human Primate Models
5. Challenges
6. Future Directions
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yactayo, S.; Staples, J.E.; Millot, V.; Cibrelus, L.; Ramon-Pardo, P. Epidemiology of chikungunya in the Americas. J. Infect. Dis. 2016, 214, S441–S445. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.; Kiran, D.H.N.; Manohar, I.; Kumar, D. Epidemiology, clinical manifestations, and diagnosis of chikungunya fever: Lessons learned from the re-emerging epidemic. Indian J. Dermatol. 2010, 55, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Caglioti, C.; Lalle, E.; Castilletti, C.; Carletti, F.; Capobianchi, M.R.; Bordi, L. Chikungunya virus infection: An overview. New Microbiol. 2013, 36, 211–227. [Google Scholar] [PubMed]
- Suhrbier, A.; Jaffar-Bandjee, M.C.; Gasque, P. Arthritogenic alphaviruses—An overview. Nat. Rev. Rheumatol. 2012, 8, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Rougeron, V.; Sam, I.C.; Caron, M.; Nkoghe, D.; Leroy, E.; Roques, P. Chikungunya, a paradigm of neglected tropical disease that emerged to be a new health global risk. J. Clin. Virol. 2015, 64, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Thiberville, S.D.; Moyen, N.; Dupuis-Maguiraga, L.; Nougairede, A.; Gould, E.A.; Roques, P.; de Lamballerie, X. Chikungunya fever: Epidemiology, clinical syndrome, pathogenesis and therapy. Antivir. Res. 2013, 99, 345–370. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Mamidi, P.; Kumar, A.; Basantray, I.; Bramha, U.; Dixit, A.; Maiti, P.K.; Singh, S.; Suryawanshi, A.R.; Chattopadhyay, S.; et al. Development of novel antibodies against non-structural proteins nsP1, nsP3 and nsP4 of chikungunya virus: Potential use in basic research. Arch. Virol. 2015, 160, 2749–2761. [Google Scholar] [CrossRef] [PubMed]
- Petitdemange, C.; Wauquier, N.; Vieillard, V. Control of immunopathology during chikungunya virus infection. J. Allergy Clin. Immunol. 2015, 135, 846–855. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, O.; Albert, M.L. Biology and pathogenesis of chikungunya virus. Nat. Rev. Microbiol. 2010, 8, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Mason, P.J.; Haddow, A.J. An epidemic of virus disease in Southern Province, Tanganyika Territory, in 1952–1953. Trans. R. Soc. Trop. Med. Hyg. 1957, 51, 238–240. [Google Scholar] [CrossRef]
- Mohan, A. Chikungunya fever: Clinical manifestations & management. Indian J. Med. Res. 2006, 124, 471–474. [Google Scholar] [PubMed]
- Kosasih, H.; de Mast, Q.; Widjaja, S.; Sudjana, P.; Antonjaya, U.; Ma’roef, C.; Riswari, S.F.; Porter, K.R.; Burgess, T.H.; Alisjahbana, B.; et al. Evidence for Endemic Chikungunya Virus Infections in Bandung, Indonesia. PLoS Negl. Trop. Dis. 2013, 7, e2483. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.K.; Chua, K.B.; Hooi, P.S.; Rahimah, M.A.; Kumari, S.; Tharmaratnam, M.; Chuah, S.K.; Smith, D.W.; Sampson, I.A. Chikungunya infection—An emerging disease in Malaysia. Southeast Asian J. Trop. Med. Public Health 2001, 32, 447–451. [Google Scholar] [PubMed]
- Laras, K.; Sukri, N.C.; Larasati, R.P.; Bangs, M.J.; Kosim, R.; Djauzi, A.; Wandra, T.; Master, J.; Kosasih, H.; Hartati, S.; et al. Tracking the re-emergence of epidemic chikungunya virus in Indonesia. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Arankalle, V.A.; Shrivastava, S.; Cherian, S.; Gunjikar, R.S.; Walimbe, A.M.; Jadhav, S.M.; Sudeep, A.B.; Mishra, A.C. Genetic divergence of Chikungunya viruses in India (1963–2006) with special reference to the 2005–2006 explosive epidemic. J. Gen. Virol. 2007, 88, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, K.; Harichandrakumar, K.T.; Kumari, A.K.; Das, L.K. Burden of Chikungunya in India: Estimates of disability adjusted life years (DALY) lost in 2006 epidemic. J. Vector Borne Dis. 2009, 46, 26–35. [Google Scholar] [PubMed]
- Singh, S.K.; Unni, S.K. Chikungunya virus: Host pathogen interaction. Rev. Med. Virol. 2011, 21, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Lemant, J.; Boisson, V.; Winer, A.; Thibault, L.; André, H.; Tixier, F.; Lemercier, M.; Antok, E.; Cresta, M.P.; Grivard, P.; et al. Serious acute chikungunya virus infection requiring intensive care during the reunion island outbreak in 2005–2006. Crit. Care Med. 2008, 36, 2536–2541. [Google Scholar] [CrossRef] [PubMed]
- Angelini, P.; Macini, P.; Finarelli, A.C.; Pol, C.; Venturelli, C.; Bellini, R.; Dottori, M. Chikungunya epidemic outbreak in Emilia-Romagna (Italy) during summer 2007. Parassitologia 2008, 50, 97–98. [Google Scholar] [PubMed]
- Gibney, K.B.; Fischer, M.; Prince, H.E.; Kramer, L.D.; George, K.S.; Kosoy, O.L.; Laven, J.J.; Staples, J.E. Chikungunya fever in the United States: A fifteen year review of cases. Clin. Infect. Dis. 2011, 52, e121–e126. [Google Scholar] [CrossRef] [PubMed]
- Richi Alberti, P.; Steiner, M.; Illera Martín, Ó.; Alcocer Amores, P.; Cobo Ibáñez, T.; Muñoz Fernández, S. Imported chikungunya fever in Madrid. Reumatol. Clin. 2016, 12, 226–227. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.A.; Gallian, P.; De Lamballerie, X.; Charrel, R.N. First cases of autochthonous dengue fever and chikungunya fever in France: From bad dream to reality! Clin. Microbiol. Infect. 2010, 16, 1702–1704. [Google Scholar] [CrossRef] [PubMed]
- Grandadam, M.; Caro, V.; Plumet, S.; Thiberge, J.M.; Souares, Y.; Failloux, A.B.; Tolou, H.J.; Budelot, M.; Cosserat, D.; Leparc-Goffart, I.; et al. Chikungunya virus, southeastern France. Emerg. Infect. Dis. 2011, 17, 910–913. [Google Scholar] [CrossRef] [PubMed]
- Balavoine, S.; Pircher, M.; Hoen, B.; Herrmann-Storck, C.; Najioullah, F.; Madeux, B.; Signate, A.; Valentino, R.; Lannuzel, A.; Louis, M.S.L.; et al. Guillain-Barré syndrome and chikungunya: Description of all cases diagnosed during the 2014 outbreak in the French West Indies. Am. J. Trop. Med. Hyg. 2017, 97, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Lum, F.M.; Ng, L.F.P. Cellular and molecular mechanisms of chikungunya pathogenesis. Antivir. Res. 2015, 120, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Gérardin, P.; Barau, G.; Michault, A.; Bintner, M.; Randrianaivo, H.; Choker, G.; Lenglet, Y.; Touret, Y.; Bouveret, A.; Grivard, P.; et al. Multidisciplinary prospective study of mother-to-child chikungunya virus infections on the island of La Réunion. PLoS Med. 2008, 5, 0413–0423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couderc, T.; Lecuit, M. Chikungunya virus pathogenesis: From bedside to bench. Antivir. Res. 2015, 121, 120–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Economopoulou, A.; Dominguez, M.; Helynck, B.; Sissoko, D.; Wichmann, O.; Quenel, P.; Germonneau, P.; Quatresous, I. Atypical Chikungunya virus infections: Clinical manifestations, mortality and risk factors for severe disease during the 2005–2006 outbreak on Reunion. Epidemiol. Infect. 2009, 137, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pulgarín, D.F.; Chowdhury, F.R.; Villamil-Gomez, W.E.; Rodriguez-Morales, A.J.; Bohm, G.M.; Paniz-Mondolfi, A.E. Ophthalmologic aspects of chikungunya infection. Travel Med. Infect. Dis. 2016, 14, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Chandak, N.H.; Kashyap, R.S.; Kabra, D.; Karandikar, P.; Saha, S.S.; Morey, S.H.; Purohit, H.J.; Taori, G.M.; Daginawala, H.F. Neurological complications of Chikungunya virus infection. Neurol. India 2009, 57, 177–180. [Google Scholar] [PubMed]
- Rodriguez-Morales, A.J.; Alvarez, M.F.; Bolívar-Mejía, A.; Ramirez-Vallejo, E. Cardiovascular involvement and manifestations of systemic Chikungunya virus infection: A systematic review. F1000Research 2017, 6, 390. [Google Scholar]
- Gérardin, P.; Fianu, A.; Malvy, D.; Mussard, C.; Boussaïd, K.; Rollot, O.; Michault, A.; Gaüzere, B.A.; Bréart, G.; Favier, F. Perceived morbidity and community burden after a Chikungunya outbreak: The TELECHIK survey, a population-based cohort study. BMC Med. 2011, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manimunda, S.P.; Vijayachari, P.; Uppoor, R.; Sugunan, A.P.; Singh, S.S.; Rai, S.K.; Sudeep, A.B.; Muruganandam, N.; Chaitanya, I.K.; Guruprasad, D.R. Clinical progression of chikungunya fever during acute and chronic arthritic stages and the changes in joint morphology as revealed by imaging. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 392–399. [Google Scholar] [CrossRef] [PubMed]
- De Brito, C.A.A.; von Sohsten, A.K.A.; de Sá Leitão, C.C.; de Brito, R.C.C.M.; De Azevedo Valadares, L.D.; da Fonte, C.A.M.; de Mesquita, Z.B.; Cunha, R.V.; Luz, K.; Leão, H.M.C.; et al. Pharmacologic management of pain in patients with Chikungunya: A guideline. Rev. Soc. Bras. Med. Trop. 2016, 49, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Mejía, C.R.; López-Vélez, R. Tropical arthritogenic alphaviruses. Reumatol. Clin. 2017. [Google Scholar] [CrossRef]
- Mavalankar, D.; Shastri, P.; Bandyopadhyay, T.; Parmar, J.; Ramani, K.V. Increased mortality rate associated with chikungunya epidemic, Ahmedabad, India. Emerg. Infect. Dis. 2008, 14, 412–415. [Google Scholar] [CrossRef] [PubMed]
- Erin Staples, J.; Breiman, R.F.; Powers, A.M. Chikungunya fever: An epidemiological review of a re-emerging infectious disease. Clin. Infect. Dis. 2009, 49, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Cardona-Ospina, J.A.; Henao-SanMartin, V.; Paniz-Mondolfi, A.E.; Rodríguez-Morales, A.J. Mortality and fatality due to Chikungunya virus infection in Colombia. J. Clin. Virol. 2015, 70, 14–15. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, M.; Mittal, V.; Bhattacharya, D.; Rana, U.V.S.; Lal, S. Chikungunya fever: A re-emerging viral infection. Indian J. Med. Microbiol. 2008, 26, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Mowatt, L.; Jackson, S.T. Chikungunya in the Caribbean: An Epidemic in the Making. Infect. Dis. Ther. 2014, 3, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Vazeille, M.; Moutailler, S.; Coudrier, D.; Rousseaux, C.; Khun, H.; Huerre, M.; Thiria, J.; Dehecq, J.S.; Fontenille, D.; Schuffenecker, I.; et al. Two Chikungunya isolates from the outbreak of La Reunion (Indian Ocean) exhibit different patterns of infection in the mosquito, Aedes albopictus. PLoS ONE 2007, 2, e1168. [Google Scholar] [CrossRef] [PubMed]
- Dotters-Katz, S.K.; Grace, M.R.; Strauss, R.A.; Chescheir, N.; Kuller, J.A. Chikungunya fever: Obstetric considerations on an emerging virus. Obstet. Gynecol. Surv. 2015, 70, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Vazeille, M.; Mousson, L.; Failloux, A.B. Failure to demonstrate experimental vertical transmission of the epidemic strain of chikungunya virus in Aedes albopictus from La Réunion Island, Indian Ocean. Mem. Inst. Oswaldo Cruz 2009, 104, 632–635. [Google Scholar] [CrossRef] [PubMed]
- Sourisseau, M.; Schilte, C.; Casartelli, N.; Trouillet, C.; Guivel-Benhassine, F.; Rudnicka, D.; Sol-Foulon, N.; Le Roux, K.; Prevost, M.; Fsihi, H.; et al. Characterization of reemerging chikungunya virus. PLoS Pathog. 2007, 3, e89. [Google Scholar] [CrossRef] [PubMed]
- Her, Z.; Malleret, B.; Chan, M.; Ong, E.K.; Wong, S.C.; Kwek, D.J.; Tolou, H.; Lin, R.T.; Tambyah, P.A.; Renia, L.; et al. Active infection of human blood monocytes by Chikungunya virus triggers an innate immune response. J. Immunol. 2010, 184, 5903–5913. [Google Scholar] [CrossRef] [PubMed]
- Kam, Y.W.; Ong, E.K.S.; Rénia, L.; Tong, J.C.; Ng, L.F.P. Immuno-biology of Chikungunya and implications for disease intervention. Microbes Infect. 2009, 11, 1186–1196. [Google Scholar] [CrossRef] [PubMed]
- Schilte, C.; Buckwalter, M.R.; Laird, M.E.; Diamond, M.S.; Schwartz, O.; Albert, M.L. Cutting edge: Independent roles for IRF-3 and IRF-7 in hematopoietic and nonhematopoietic cells during host response to Chikungunya infection. J. Immunol. 2012, 188, 2967–2971. [Google Scholar] [CrossRef] [PubMed]
- White, L.K.; Sali, T.; Alvarado, D.; Gatti, E.; Pierre, P.; Streblow, D.; DeFilippis, V.R. Chikungunya virus induces IPS-1-dependent innate immune activation and protein kinase R-independent translational shutoff. J. Virol. 2011, 85, 606–620. [Google Scholar] [CrossRef] [PubMed]
- Pardigon, N. The biology of chikungunya: A brief review of what we still do not know. Pathol. Biol. 2009, 57, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Wauquier, N.; Becquart, P.; Nkoghe, D.; Padilla, C.; Ndjoyi-Mbiguino, A.; Leroy, E.M. The acute phase of Chikungunya virus infection in humans is associated with strong innate immunity and T CD8 cell activation. J. Infect. Dis. 2011, 204, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Mahauad-Fernandez, W.D.; Jones, P.H.; Okeoma, C.M. Critical role for bone marrow stromal antigen 2 in acute Chikungunya virus infection. J. Gen. Virol. 2014, 95, 2450–2461. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Foo, S.S.; Sims, N.A.; Herrero, L.J.; Walsh, N.C.; Mahalingam, S. Arthritogenic alphaviruses: New insights into arthritis and bone pathology. Trends Microbiol. 2015, 23, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Das, T.; Hoarau, J.J.; Bandjee, M.C.J.; Marianne, M.; Gasque, P. Multifaceted innate immune responses engaged by astrocytes, microglia and resident dendritic cells against Chikungunya neuroinfection. J. Gen. Virol. 2015, 96, 294–310. [Google Scholar] [CrossRef] [PubMed]
- Van Duijl-Richter, M.K.S.; Hoornweg, T.E.; Rodenhuis-Zybert, I.A.; Smit, J.M. Early events in chikungunya virus infection—From virus cell binding to membrane fusion. Viruses 2015, 7, 3647–3674. [Google Scholar] [CrossRef] [PubMed]
- Goupil, B.A.; Mores, C.N. A review of chikungunya virus-induced arthralgia: Clinical manifestations, therapeutics, and pathogenesis. Open Rheumatol. J. 2016, 10, 129–140. [Google Scholar] [PubMed]
- Wichit, S.; Hamel, R.; Bernard, E.; Talignani, L.; Diop, F.; Ferraris, P.; Liegeois, F.; Ekchariyawat, P.; Luplertlop, N.; Surasombatpattana, P.; et al. Imipramine Inhibits Chikungunya Virus Replication in Human Skin Fibroblasts through Interference with Intracellular Cholesterol Trafficking. Sci. Rep. 2017, 7, 3145. [Google Scholar] [CrossRef] [PubMed]
- Bouquillard, E.; Combe, B. A report of 21 cases of rheumatoid arthritis following Chikungunya fever. A mean follow-up of two years. Jt. Bone Spine 2009, 76, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Foissac, M.; Javelle, E.; Ray, S.; Guérin, B.; Simon, F. Post-chikungunya rheumatoid arthritis, saint martin. Emerg. Infect. Dis. 2015, 21, 530–532. [Google Scholar] [CrossRef] [PubMed]
- Chopra, A.; Anuradha, V.; Lagoo-Joshi, V.; Kunjir, V.; Salvi, S.; Saluja, M. Chikungunya virus aches and pains: An emerging challenge. Arthritis Rheum. 2008, 58, 2921–2922. [Google Scholar] [CrossRef] [PubMed]
- Hussain, K.M.; Lee, R.C.H.; Ng, M.M.L.; Chu, J.J.H. Establishment of a novel primary human skeletal myoblast cellular model for chikungunya virus infection and pathogenesis. Sci. Rep. 2016, 6, 21406. [Google Scholar] [CrossRef] [PubMed]
- Parashar, D.; Paingankar, M.S.; Kumar, S.; Gokhale, M.D.; Sudeep, A.B.; Shinde, S.B.; Arankalle, V.A. Administration of E2 and NS1 siRNAs inhibit chikungunya virus replication in vitro and protects mice infected with the virus. PLoS Negl. Trop. Dis. 2013, 7, e2405. [Google Scholar] [CrossRef] [PubMed]
- Phuklia, W.; Kasisith, J.; Modhiran, N.; Rodpai, E.; Thannagith, M.; Thongsakulprasert, T.; Smith, D.R.; Ubol, S. Osteoclastogenesis induced by CHIKV-infected fibroblast-like synoviocytes: A possible interplay between synoviocytes and monocytes/macrophages in CHIKV-induced arthralgia/arthritis. Virus Res. 2013, 177, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Noret, M.; Herrero, L.; Rulli, N.; Rolph, M.; Smith, P.N.; Li, R.W.; Roques, P.; Gras, G.; Mahalingam, S. Interleukin 6, RANKL, and osteoprotegerin expression by chikungunya virus-infected human osteoblasts. J. Infect. Dis. 2012, 206, 455–457. [Google Scholar] [CrossRef] [PubMed]
- Bhakat, S.; Soliman, M.E. Chikungunya virus (CHIKV) inhibitors from natural sources: A medicinal chemistry perspective. J. Nat. Med. 2015, 69, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Rothan, H.A.; Bahrani, H.; Abdulrahman, A.Y.; Mohamed, Z.; Teoh, T.C.; Othman, S.; Rashid, N.N.; Rahman, N.A.; Yusof, R. Mefenamic acid in combination with ribavirin shows significant effects in reducing chikungunya virus infection in vitro and in vivo. Antivir. Res. 2016, 127, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Haese, N.N.; Broeckel, R.M.; Hawman, D.W.; Heise, M.T.; Morrison, T.E.; Streblow, D.N. Animal models of chikungunya virus infection and disease. J. Infect. Dis. 2016, 214, S482–S487. [Google Scholar] [CrossRef] [PubMed]
- Hugo, L.E.; Prow, N.A.; Tang, B.; Devine, G.; Suhrbier, A. Chikungunya virus transmission between Aedes albopictus and laboratory mice. Parasit. Vectors 2016, 9, 555. [Google Scholar] [CrossRef] [PubMed]
- Couderc, T.; Chrétien, F.; Schilte, C.; Disson, O.; Brigitte, M.; Guivel-Benhassine, F.; Touret, Y.; Barau, G.; Cayet, N.; Schuffenecker, I.; et al. A mouse model for Chikungunya: Young age and inefficient type-I interferon signaling are risk factors for severe disease. PLoS Pathog. 2008, 4, e29. [Google Scholar] [CrossRef] [PubMed]
- Werneke, S.W.; Schilte, C.; Rohatgi, A.; Monte, K.J.; Michault, A.; Arenzana-Seisdedos, F.; Vanlandingham, D.L.; Higgs, S.; Fontanet, A.; Albert, M.L.; et al. ISG15 is critical in the control of chikungunya virus infection independent of UbE1l mediated conjugation. PLoS Pathog. 2011, 7, e1002322. [Google Scholar] [CrossRef] [PubMed]
- Hawman, D.W.; Stoermer, K.; Montgomery, S.A.; Pal, P.; Oko, L.; Diamond, M.S.; Morrison, T.E. Chronic joint disease caused by persistent Chikungunya virus infection is controlled by the adaptive immune response. J. Virol. 2013, 87, 13878–13888. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.L.; Hodge, H.M.; Campbell, W.E., Jr. Growth of chikungunya virus in baby hamster kidney cell (BHK-21-clone 13) suspension cultures. Appl. Microbiol. 1971, 21, 338–341. [Google Scholar] [PubMed]
- Bosco-Lauth, A.M.; Han, S.; Hartwig, A.; Bowen, R.A. Development of a hamster model for chikungunya virus infection and pathogenesis. PLoS ONE 2015, 10, e0130150. [Google Scholar] [CrossRef] [PubMed]
- Gardner, M.B.; Luciw, P.A. Macaque models of human infectious disease. ILAR J. 2008, 49, 220–255. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.W. The newala epidemic: III. The virus: Isolation, pathogenic properties and relationship to the epidemic. J. Hyg. 1956, 54, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.D.; Singh, K.R. Experimental infection of Macaca radiata with Chikungunya virus and transmission of virus by mosquitoes. Indian J. Med. Res. 1968, 56, 802–811. [Google Scholar] [PubMed]
- Inoue, S.; Morita, K.; Matias, R.R.; Tuplano, J.V.; Resuello, R.R.G.; Candelario, J.R.; Cruz, D.J.M.; Mapua, C.A.; Hasebe, F.; Igarashi, A.; et al. Distribution of three arbovirus antibodies among monkeys (Macaca fascicularis) in the Philippines. J. Med. Primatol. 2003, 32, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Labadie, K.; Larcher, T.; Joubert, C.; Mannioui, A.; Delache, B.; Brochard, P.; Guigand, L.; Dubreil, L.; Lebon, P.; Verrier, B.; et al. Chikungunya disease in nonhuman primates involves long-term viral persistence in macrophages. J. Clin. Investig. 2010, 120, 894–906. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.I.; Clark, D.C.; Pesavento, P.; Lerche, N.W.; Luciw, P.A.; Reisen, W.K.; Brault, A.C. Comparative pathogenesis of epidemic and enzootic Chikungunya viruses in a pregnant Rhesus macaque model. Am. J. Trop. Med. Hyg. 2010, 83, 1249–1258. [Google Scholar] [CrossRef] [PubMed]
- Messaoudi, I.; Vomaske, J.; Totonchy, T.; Kreklywich, C.N.; Haberthur, K.; Springgay, L.; Brien, J.D.; Diamond, M.S.; DeFilippis, V.R.; Streblow, D.N. Chikungunya virus infection results in higher and persistent viral replication in aged rhesus macaques due to defects in anti-viral immunity. PLoS Negl. Trop. Dis. 2013, 7, e2343. [Google Scholar] [CrossRef] [PubMed]
- Nakao, E.; Hotta, S. Immunogenicity of purified, inactivated chikungunya virus in monkeys. Bull. World Health Org. 1973, 48, 559–562. [Google Scholar] [PubMed]
- Turell, M.J.; Malinoski, F.J. Limited potential for mosquito transmission of a live attenuated chikungunya virus vaccine. Am. J. Trop. Med. Hyg. 1992, 47, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Kam, Y.W.; Lee, W.W.L.; Simarmata, D.; Le Grand, R.; Tolou, H.; Merits, A.; Roques, P.; Ng, L.F.P. Unique epitopes recognized by antibodies induced in chikungunya virus-infected non-human primates: Implications for the study of immunopathology and vaccine development. PLoS ONE 2014, 9, e95647. [Google Scholar] [CrossRef] [PubMed]
- Pal, P.; Fox, J.M.; Hawman, D.W.; Huang, Y.J.S.; Messaoudi, I.; Kreklywich, C.; Denton, M.; Legasse, A.W.; Smith, P.P.; Johnson, S.; et al. Chikungunya viruses that escape monoclonal antibody therapy are clinically attenuated, stable, and not purified in mosquitoes. J. Virol. 2014, 88, 8213–8226. [Google Scholar] [CrossRef] [PubMed]
- Mallilankaraman, K.; Shedlock, D.J.; Bao, H.; Kawalekar, O.U.; Fagone, P.; Ramanathan, A.A.; Ferraro, B.; Stabenow, J.; Vijayachari, P.; Sundaram, S.G.; et al. A DNA vaccine against Chikungunya virus is protective in mice and induces neutralizing antibodies in mice and nonhuman primates. PLoS Negl. Trop. Dis. 2011, 5, e928. [Google Scholar] [CrossRef] [PubMed]
- Gardner, J.; Anraku, I.; Le, T.T.; Larcher, T.; Major, L.; Roques, P.; Schroder, W.A.; Higgs, S.; Suhrbier, A. Chikungunya virus arthritis in adult wild-type mice. J. Virol. 2010, 84, 8021–8032. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Lecuit, M. Chikungunya virus and the global spread of a mosquito-borne disease. N. Engl. J. Med. 2015, 372, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Marcombe, S.; Farajollahi, A.; Healy, S.P.; Clark, G.G.; Fonseca, D.M. Insecticide resistance status of United States populations of Aedes albopictus and mechanisms involved. PLoS ONE 2014, 9, e101992. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Yang, J.; Luo, L.; Yang, Z.; Liu, Q. Who is vulnerable to dengue fever? A community survey of the 2014 outbreak in Guangzhou, China. Int. J. Environ. Res. Public Health 2016, 13, 712. [Google Scholar] [CrossRef] [PubMed]
- Cardona-Ospina, J.A.; Villamil-Gómez, W.E.; Jimenez-Canizales, C.E.; Castañeda-Hernández, D.M.; Rodríguez-Morales, A.J. Estimating the burden of disease and the economic cost attributable to chikungunya, Colombia, 2014. Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Chakradhar, S. Disease in three dimensions: Tissue engineering takes on infectious disease. Nat. Med. 2017, 3, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Mills, M.; Estes, M.K. Physiologically relevant human tissue models for infectious diseases. Drug Discov. Today 2016, 21, 1540–1552. [Google Scholar] [CrossRef] [PubMed]
- Chau, D.Y.; Johnson, C.; MacNeil, S.; Haycock, J.W.; Ghaemmaghami, A.M. The development of a 3D immunocompetent model of human skin. Biofabrication 2013, 5, 035011. [Google Scholar] [CrossRef] [PubMed]
- Kuhbacher, A.; Henkel, H.; Stevens, P.; Grumaz, C.; Finkelmeier, D.; Burger-Kentischer, A.; Sohn, K.; Rupp, S. Central Role for Dermal Fibroblasts in Skin Model Protection against Candida albicans. J. Infect. Dis. 2017, 215, 1742–1752. [Google Scholar] [CrossRef] [PubMed]
- Bartfeld, S.; Clevers, H. Stem cell-derived organoids and their application for medical research and patient treatment. J. Mol. Med. 2017, 95, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Dang, J.; Tiwari, S.K.; Lichinchi, G.; Qin, Y.; Patil, V.S.; Eroshkin, A.M.; Rana, T.M. Zika Virus Depletes Neural Progenitors in Human Cerebral Organoids through Activation of the Innate Immune Receptor TLR3. Cell Stem Cell 2016, 19, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Nguyen, H.N.; Song, M.M.; Hadiono, C.; Ogden, S.C.; Hammack, C.; Yao, B.; Hamersky, G.R.; Jacob, F.; Zhong, C.; et al. Brain-Region-Specific Organoids Using Mini-bioreactors for Modeling ZIKV Exposure. Cell 2016, 165, 1238–1254. [Google Scholar] [CrossRef] [PubMed]
- Goupil, B.A.; McNulty, M.A.; Martin, M.J.; McCracken, M.K.; Christofferson, R.C.; Mores, C.N. Novel lesions of bones and joints associated with chikungunya virus infection in two mouse models of disease: New insights into disease pathogenesis. PLoS ONE 2016, 11, e0155243. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Liu, L.; Shi, Y.; Qiu, J.; Fang, W.; Rong, M.; Guo, Z.; Gao, W. Characterization of silk fibroin/chitosan 3D porous scaffold and in vitro cytology. PLoS ONE 2015, 10, e0128658. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Location | Year/Duration | Affected |
|---|---|---|
| Lamu Island, Kenya | 2004 | 13,500 |
| La Réunion | 2005–2006 | 255,000 |
| India | 2005 | 1,380,000 |
| Mauritius | 2006 | 13,500 |
| Gabon | 2007 | 20,000 |
| Thailand | 2008–2009 | 49,000 |
| Republic of Congo | 2011 | 8000 |
| Martinique-Guadeloupe | 2014 | 308,000 |
| French Polynesia | 2014–2015 | 66,000 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganesan, V.K.; Duan, B.; Reid, S.P. Chikungunya Virus: Pathophysiology, Mechanism, and Modeling. Viruses 2017, 9, 368. https://doi.org/10.3390/v9120368
Ganesan VK, Duan B, Reid SP. Chikungunya Virus: Pathophysiology, Mechanism, and Modeling. Viruses. 2017; 9(12):368. https://doi.org/10.3390/v9120368
Chicago/Turabian StyleGanesan, Vaishnavi K., Bin Duan, and St Patrick Reid. 2017. "Chikungunya Virus: Pathophysiology, Mechanism, and Modeling" Viruses 9, no. 12: 368. https://doi.org/10.3390/v9120368
APA StyleGanesan, V. K., Duan, B., & Reid, S. P. (2017). Chikungunya Virus: Pathophysiology, Mechanism, and Modeling. Viruses, 9(12), 368. https://doi.org/10.3390/v9120368