A Chinese Variant Marek’s Disease Virus Strain with Divergence between Virulence and Vaccine Resistance

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Clinical Samples
2.2. Viral Isolation and Identification
2.3. Screening for Causative Agents
2.4. Animal Experiments
2.5. DNA Extraction and TaqMan Real-Time Polymerase Chain Teaction (qPCR)
2.6. Statistical Analysis
3. Results
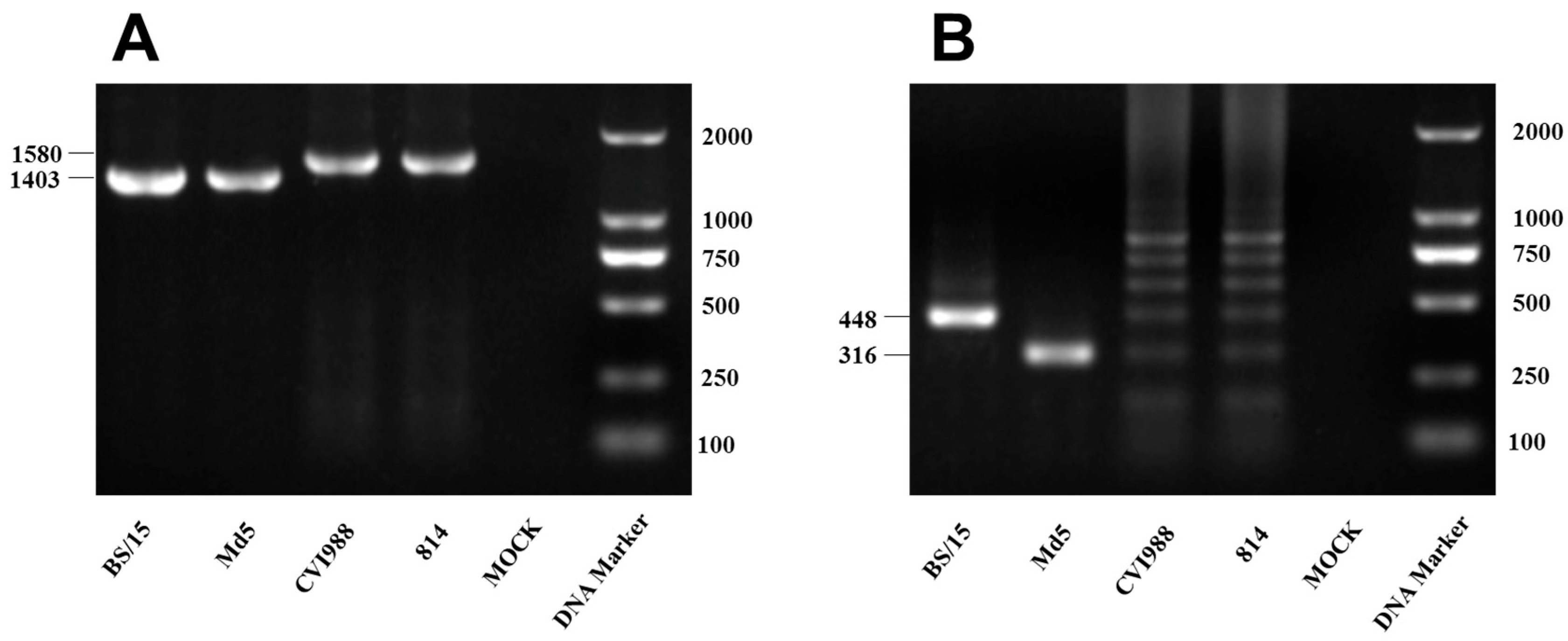
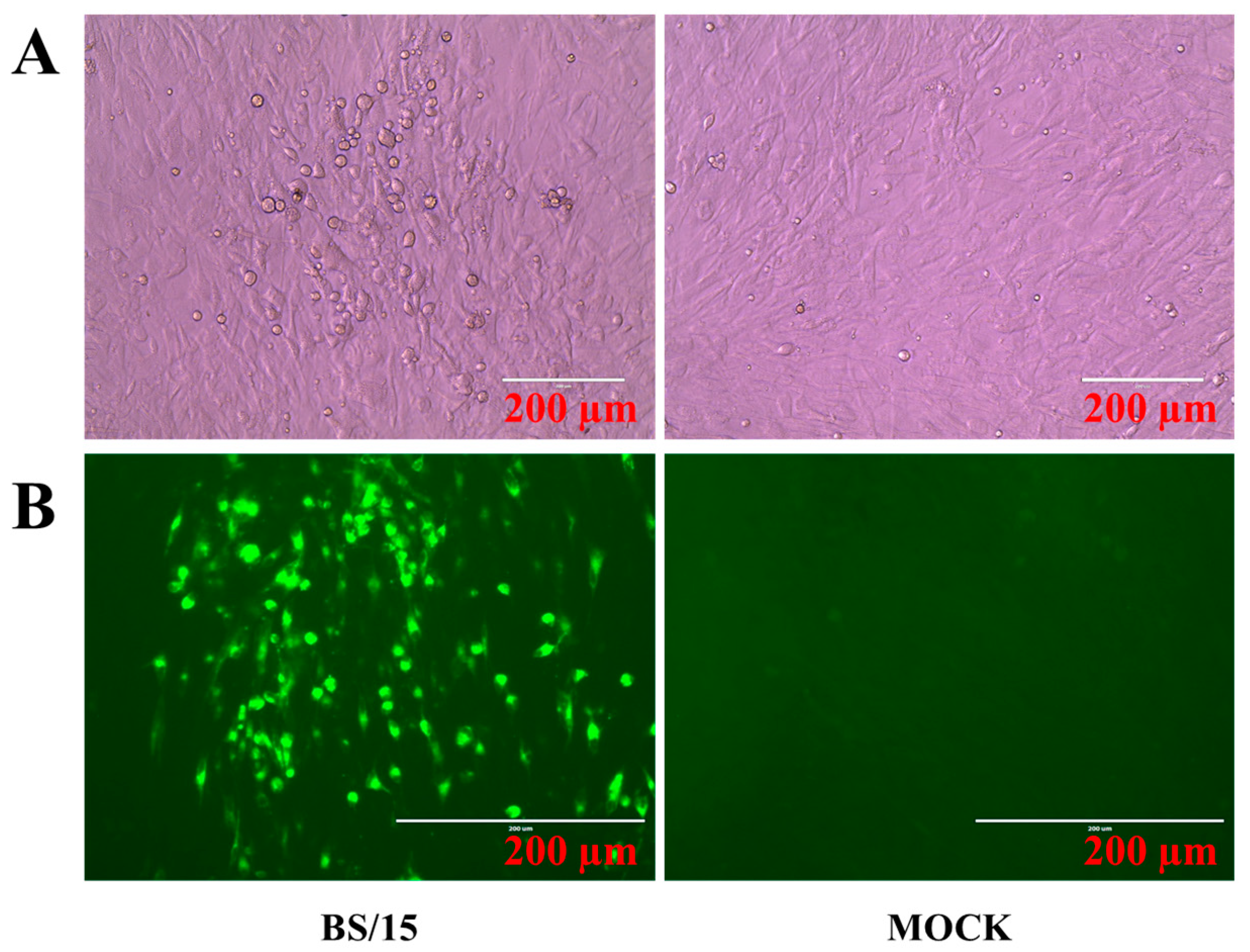
3.1. Marek’s Disease Virus (MDV) Strain BS/15 Isolated in China
3.2. Virulence Studies of MDV BS/15 and Protective Potency Evaluation of Vaccines
3.2.1. MD Incidence, Mortality, and Tumor Rates per Group
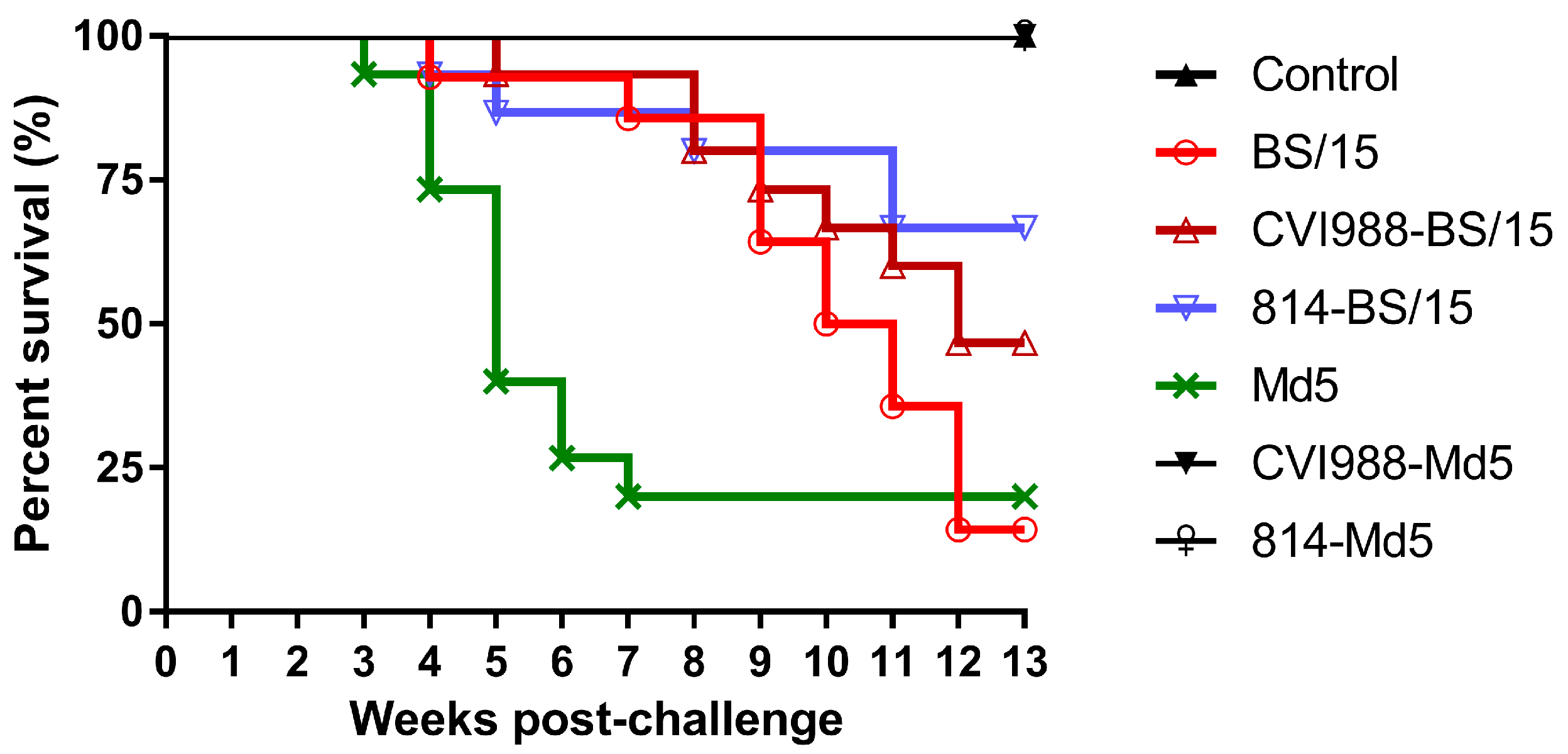
3.2.2. Survival Analysis
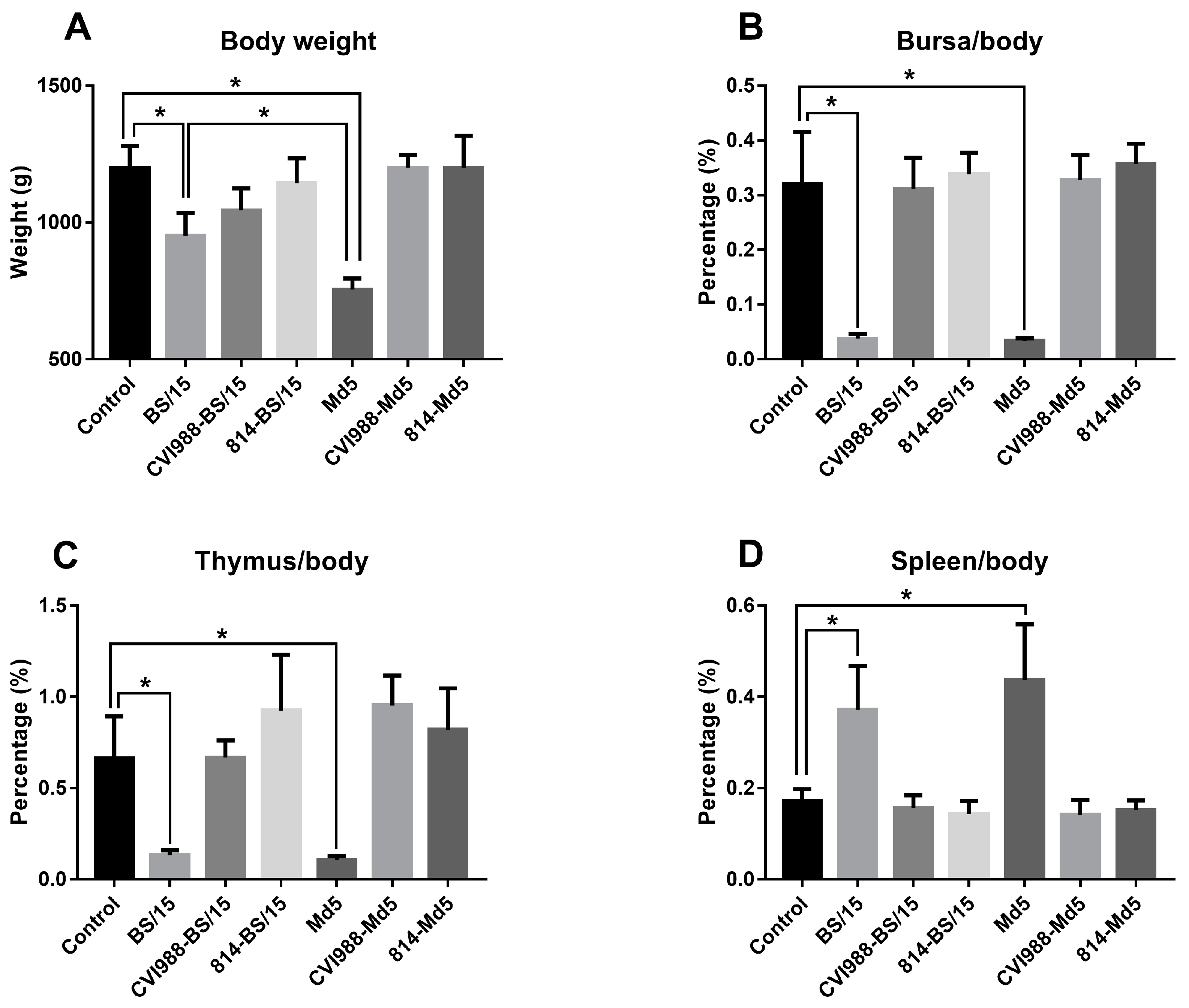
3.2.3. Developmental Disorders and Immune Organ Damages
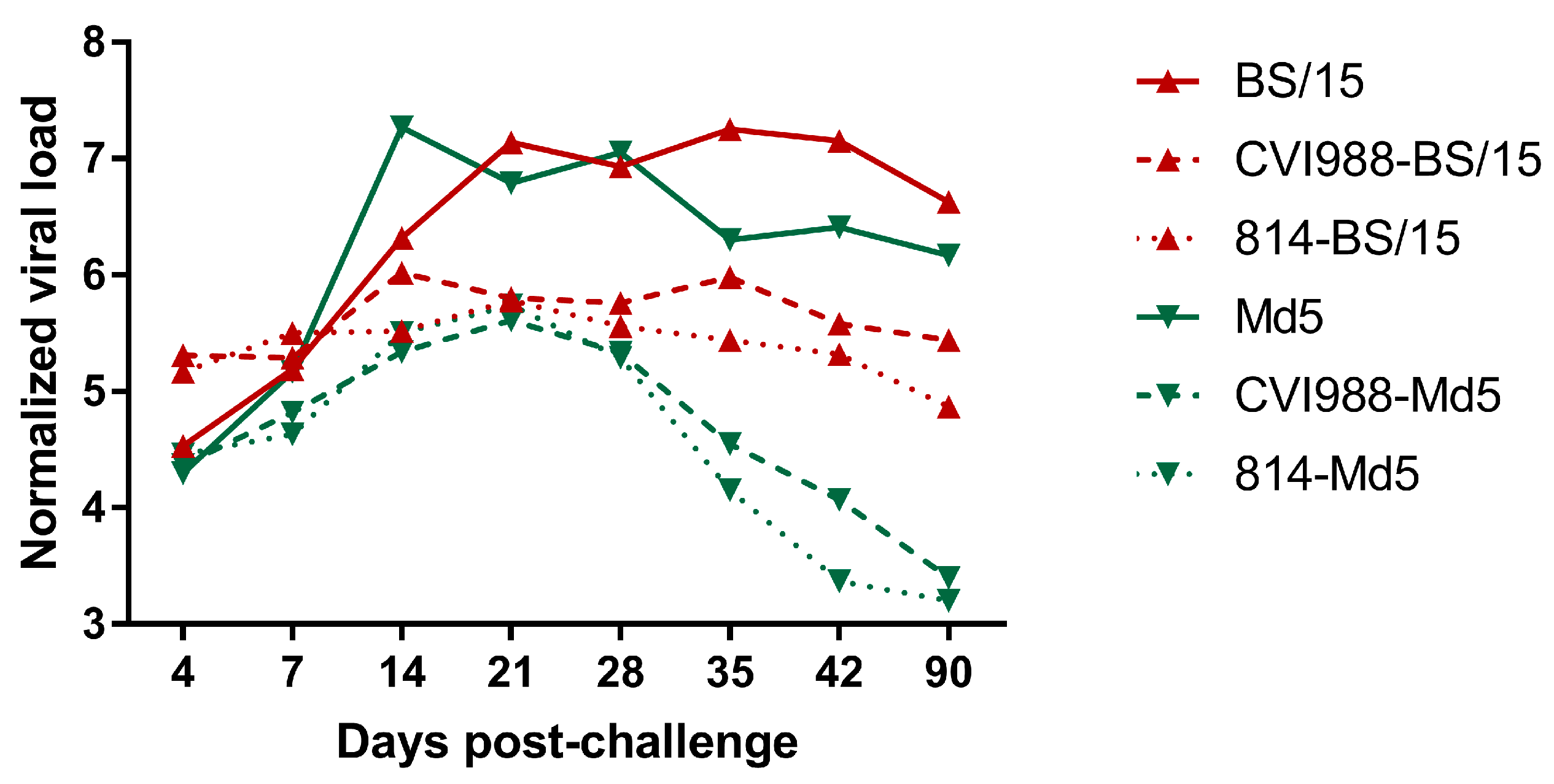
3.2.4. MDV Genome Load in Feather Pulps
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Osterrieder, N.; Kamil, J.P.; Schumacher, D.; Tischer, B.K.; Trapp, S. Marek’s disease virus: from miasma to model. Nature Rev. Microbiol. 2006, 4, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Witter, R.L. Characteristics of Marek’s disease viruses isolated from vaccinated commercial chicken flocks: association of viral pathotype with lymphoma frequency. Avian Dis. 1983, 27, 113–132. [Google Scholar] [CrossRef] [PubMed]
- Witter, R.L. Increased virulence of Marek’s disease virus field isolates. Avian Dis. 1997, 41, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, W.; Purchase, H.G.; Burmester, B.R. Protection against Marek’s disease by vaccination with a herpesvirus of turkeys. Avian Dis. 1970, 14, 413–429. [Google Scholar] [CrossRef] [PubMed]
- Rispens, B.H.; Van, V.H.; Mastenbroek, N.; Maas, H.J.; Schat, K.A. Control of Marek’s disease in the Netherlands. I. Isolation of an avirulent Marek’s disease virus (strain CVI988) and its use in laboratory vaccination trials. Avian Dis. 1972, 16, 108–125. [Google Scholar] [CrossRef] [PubMed]
- Witter, R.L.; Lee, L.F. Polyvalent Marek’s disease vaccines: Safety, efficacy and protective synergism in chickens with maternal antibodies. Avian Pathol. 1984, 13, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Witter, R.L. Protective Efficacy of Marek’s Disease Vaccines. Avian Dis. 1993, 37, 57–90. [Google Scholar]
- Nair, V.; Schudel, A.; Lombard, M. Successful control of Marek’s disease by vaccination. Dev. Biol. 2004, 119, 147–154. [Google Scholar]
- Atkins, K.E.; Woolhouse, M.E.J. Vaccination and reduced cohort duration can drive virulence evolution: Marek’s disease virus and industrialized agriculture. Evolution 2013, 67, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Gandon, S.; Mackinnon, M.; Nee, S.; Read, A. Imperfect vaccination: some epidemiological and evolutionary consequences. Proc. R. Soc. Lond. [Biol] 2003, 270, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Read, A.F.; Baigent, S.J.; Powers, C.; Kgosana, L.B.; Blackwell, L.; Smith, L.P.; Kennedy, D.A.; Walkdenbrown, S.W.; Nair, V.K. Imperfect Vaccination Can Enhance the Transmission of Highly Virulent Pathogens. PLoS Biol. 2015, 13, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Ewald, P.W. Imperfect Vaccines and the Evolution of Pathogen Virulence. Nature 2001, 414, 751–756. [Google Scholar]
- Nair, V. Evolution of Marek’s disease – A paradigm for incessant race between the pathogen and the host. Vet. J. 2005, 170, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.P.; Li, Z.J.; Bao, K.Y.; Lv, H.C.; Gao, Y.L.; Gao, H.L.; Qi, X.L.; Cui, H.Y.; Wang, Y.Q.; Ren, X.G. Pathogenic characteristics of Marek’s disease virus field strains prevalent in China and the effectiveness of existing vaccines against them. Vet. Microbiol. 2015, 177, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.C.; Zhang, Y.P.; Sun, G.R.; Gao, Y.L.; Li, Z.J.; Zheng, H.W.; Bao, K.Y.; Wang, X.M.; Liu, C.J. Assessments of a PCR method for detection and identification of virulent Marek’s disease virus and vaccine strain for Marek’s disease diagnosis. Chin. J. Prev. Vet. Med. 2016, 38, 567–571. [Google Scholar]
- Gopal, S.; Manoharan, P.; Kathaperumal, K.; Chidambaram, B.; Divya, K.C. Differential detection of avian oncogenic viruses in poultry layer farms and Turkeys by use of multiplex PCR. J. Clin. Microbiol. 2012, 50, 2668–2673. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.; Li, D.; Zhu, H.; Liu, W.; Qin, L.; Liu, Z.; Wu, G.; Wang, Y.; Qi, X.; Gao, H. Development of an antigen-capture ELISA for the detection of avian leukosis virus p27 antigen. J. Virol. Methods 2012, 187, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Shi, X.; Zhang, J.; Zhao, Y.; Cui, H.; Hu, S.; Gao, H.; Cui, X.; Wang, Y.F. Identification of a conserved B-cell epitope on reticuloendotheliosis virus envelope protein by screening a phage-displayed random peptide library. PLoS ONE 2012, 7, e49842. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.T.; Gao, Y.L.; Pan, W.; Deng, X.Y.; Sun, F.F.; Kai, L.I.; Xiao-Le, Q.I.; Gao, H.L.; Liu, C.N.; Wang, X.M. Investigation of co-infection of ALV-J with REV, MDV, CAV in layer chicken flocks in some regions of China. Chin. J. Prev. Vet. Med. 2010, 32, 90–93. [Google Scholar]
- Zhang, Y.; Liu, C.J.; Qin, Y.A.; Zhang, Y.P.; Zhang, X.W.; Hao, Y.Q. Application of duplex fluorescent quantitative polymerase-chain-reaction for detecting Marek’s disease virus serotype 1. Chin. J. Prev. Vet. Med. 2007, 29, 46–51. [Google Scholar]
- Baigent, S.J.; Petherbridge, L.J.; Howes, K.; Smith, L.P.; Currie, R.J.; Nair, V.K. Absolute quantitation of Marek’s disease virus genome copy number in chicken feather and lymphocyte samples using real-time PCR. J. Virol. Methods 2005, 123, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.M.; Burmester, B.R. Resistance to Marek’s disease at hatching in chickens vaccinated as embryos with the turkey herpesvirus. Avian Dis. 1982, 26, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Zhang, L.; Wang, J.; Chen, L.; Hu, S.; Wang, Z.; Ma, H. Isolation and analysis of a very virulent Marek’s disease virus strain in China. Virol. J. 2013, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ning, C.; Shuai, S.; Peng, S.; Zhang, Y.; Ni, H.; Cui, Z. Isolation and pathogenic analysis of virulent Marek’s disease virus field strain in China. Poult. Sci. 2016, 95, 1521–1528. [Google Scholar]
- Zhang, Y.P.; Lv, H.C.; Bao, K.Y.; Gao, Y.L.; Gao, H.L.; Qi, X.L.; Cui, H.Y.; Wang, Y.Q.; Li, K.; Gao, L. Molecular and pathogenicity characterization of Gallid herpesvirus 2 newly isolated in China from 2009 to 2013. Virus Genes 2015, 52, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Zou, H.; Shi, H.; Shao, H.; Ye, J.; Ji, M.; Wu, G.; Qin, A. Outbreak of Marek’s disease in a vaccinated broiler breeding flock during its peak egg-laying period in China. BMC Vet. Res. 2015, 11, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Witter, R.L.; Calnek, B.W.; Buscaglia, C.; Gimeno, I.M.; Schat, K.A. Classification of Marek’s disease viruses according to pathotype: philosophy and methodology. Avian Pathol. 2005, 34, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Walkden-Brown, S.W.; Islam, A.; Islam, A.; Burgess, S.K.; Groves, P.J.; Cooke, J. Pathotyping of Australian isolates of Marek’s disease virus in commercial broiler chickens vaccinated with herpesvirus of turkeys (HVT) or bivalent (HVT/SB1) vaccine and association with viral load in the spleen and feather dander. Aust. Vet. J. 2013, 91, 341–350. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primer Sequence | Product Size (bp) |
|---|---|---|
| meq | F: 5′-GGGAAATGACAGGTGAATTGTG-3′ | 1403/1580 a |
| R: 5′-TAAGGAAAATTTGTTACCCCAG-3′ | ||
| 132bpr | F: 5′-TGCGATGAAAGTGCTATGGAGG-3′ | 316–844 b |
| R: 5′-GAGAATCCCTATGAGAAAGCGC-3′ |
| Vaccine | Challenge | MD Incidence Diseased/Total (%) | PI | Mortality Deaths/Total (%) | Tumor Incidence a |
|---|---|---|---|---|---|
| None | None | 0/14 (0%) | - | 0/14 (0%) | 0% |
| None | Md5 | 15/15 (100%) | - | 12/15 (80.0%) | 40.0% |
| CVI988 | Md5 | 1/14 (7.1%) | 92.9 | 0/14 (0%) | 0% |
| 814 | Md5 | 0/13 (0%) | 100 | 0/14 (0%) | 0% |
| None | BS/15 | 14/14 (100%) | - | 12/14 (85.7%) | 64.3% |
| CVI988 | BS/15 | 10/15 (66.7%) | 33.3 | 8/15 (53.3%) | 13.3% |
| 814 | BS/15 | 5/15 (33.3%) | 66.7 | 5/15 (33.3%) | 20.0% |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, G.-r.; Zhang, Y.-p.; Lv, H.-c.; Zhou, L.-y.; Cui, H.-y.; Gao, Y.-l.; Qi, X.-l.; Wang, Y.-q.; Li, K.; Gao, L.; et al. A Chinese Variant Marek’s Disease Virus Strain with Divergence between Virulence and Vaccine Resistance. Viruses 2017, 9, 71. https://doi.org/10.3390/v9040071
Sun G-r, Zhang Y-p, Lv H-c, Zhou L-y, Cui H-y, Gao Y-l, Qi X-l, Wang Y-q, Li K, Gao L, et al. A Chinese Variant Marek’s Disease Virus Strain with Divergence between Virulence and Vaccine Resistance. Viruses. 2017; 9(4):71. https://doi.org/10.3390/v9040071
Chicago/Turabian StyleSun, Guo-rong, Yan-ping Zhang, Hong-chao Lv, Lin-yi Zhou, Hong-yu Cui, Yu-long Gao, Xiao-le Qi, Yong-qiang Wang, Kai Li, Li Gao, and et al. 2017. "A Chinese Variant Marek’s Disease Virus Strain with Divergence between Virulence and Vaccine Resistance" Viruses 9, no. 4: 71. https://doi.org/10.3390/v9040071
APA StyleSun, G. -r., Zhang, Y. -p., Lv, H. -c., Zhou, L. -y., Cui, H. -y., Gao, Y. -l., Qi, X. -l., Wang, Y. -q., Li, K., Gao, L., Pan, Q., Wang, X. -m., & Liu, C. -j. (2017). A Chinese Variant Marek’s Disease Virus Strain with Divergence between Virulence and Vaccine Resistance. Viruses, 9(4), 71. https://doi.org/10.3390/v9040071