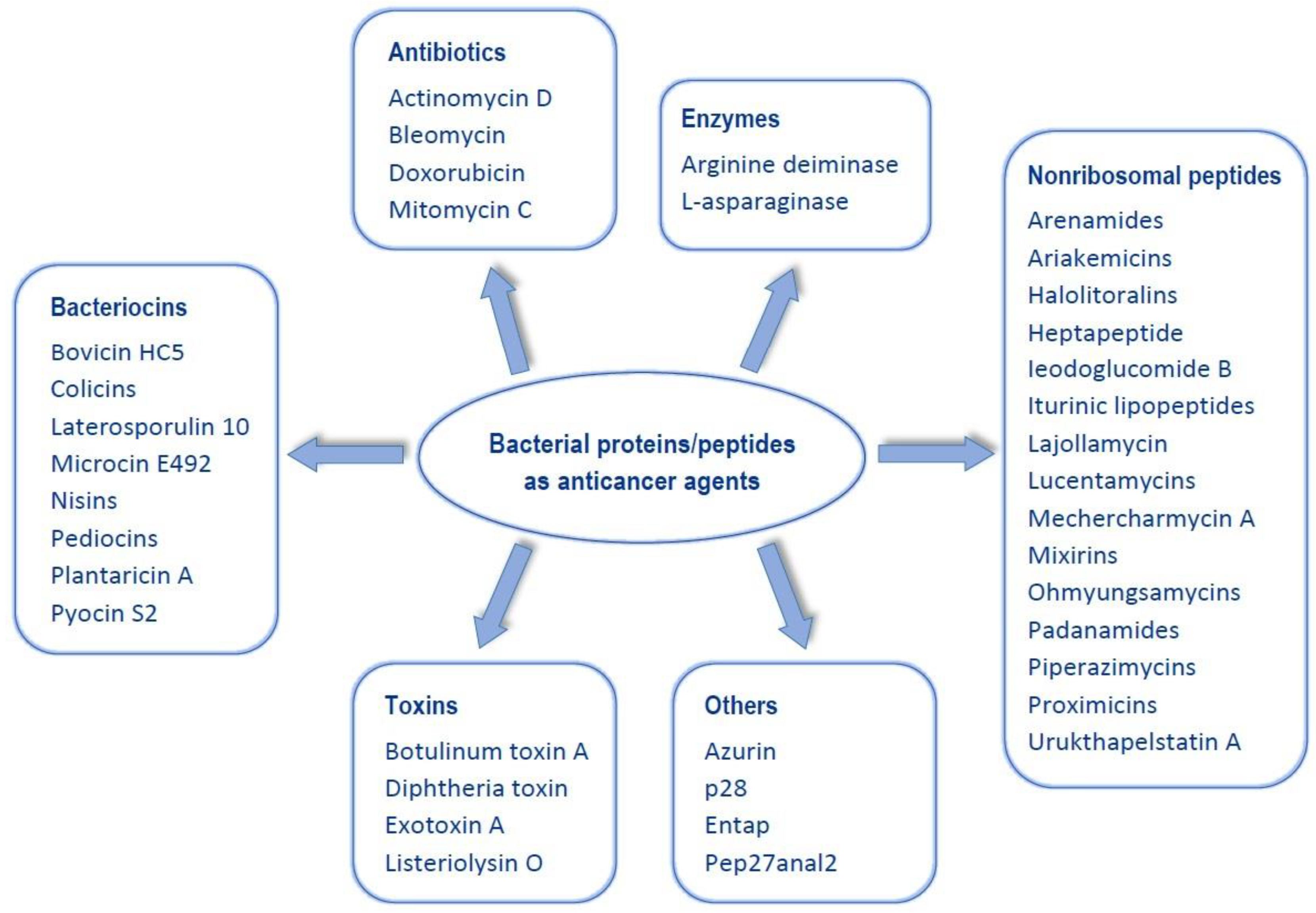
Anticancer Activity of Bacterial Proteins and Peptides



Abstract
:1. Introduction
2. Antibiotics
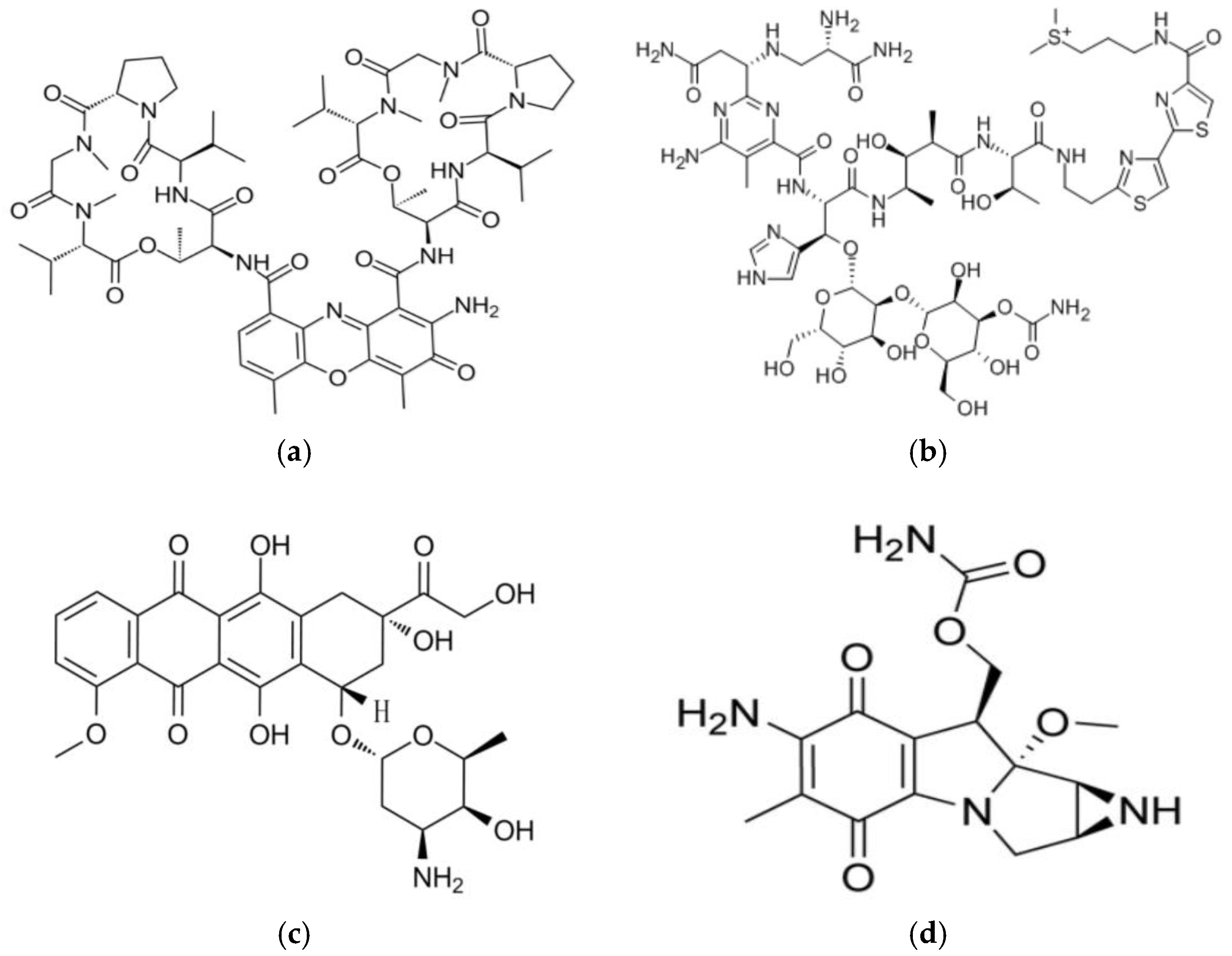
2.1. Actinomycin D
2.2. Bleomycin
2.3. Doxorubicin
2.4. Mitomycin C
3. Bacteriocins
3.1. Bovicin HC5
3.2. Colicins
3.3. Laterosporulin 10
3.4. Microcin E492
3.5. Nisins
3.6. Pediocins
3.7. Plantaricin A
3.8. Pyocins
4. Enzymes
4.1. Arginine Deiminase
4.2. l-asparaginase
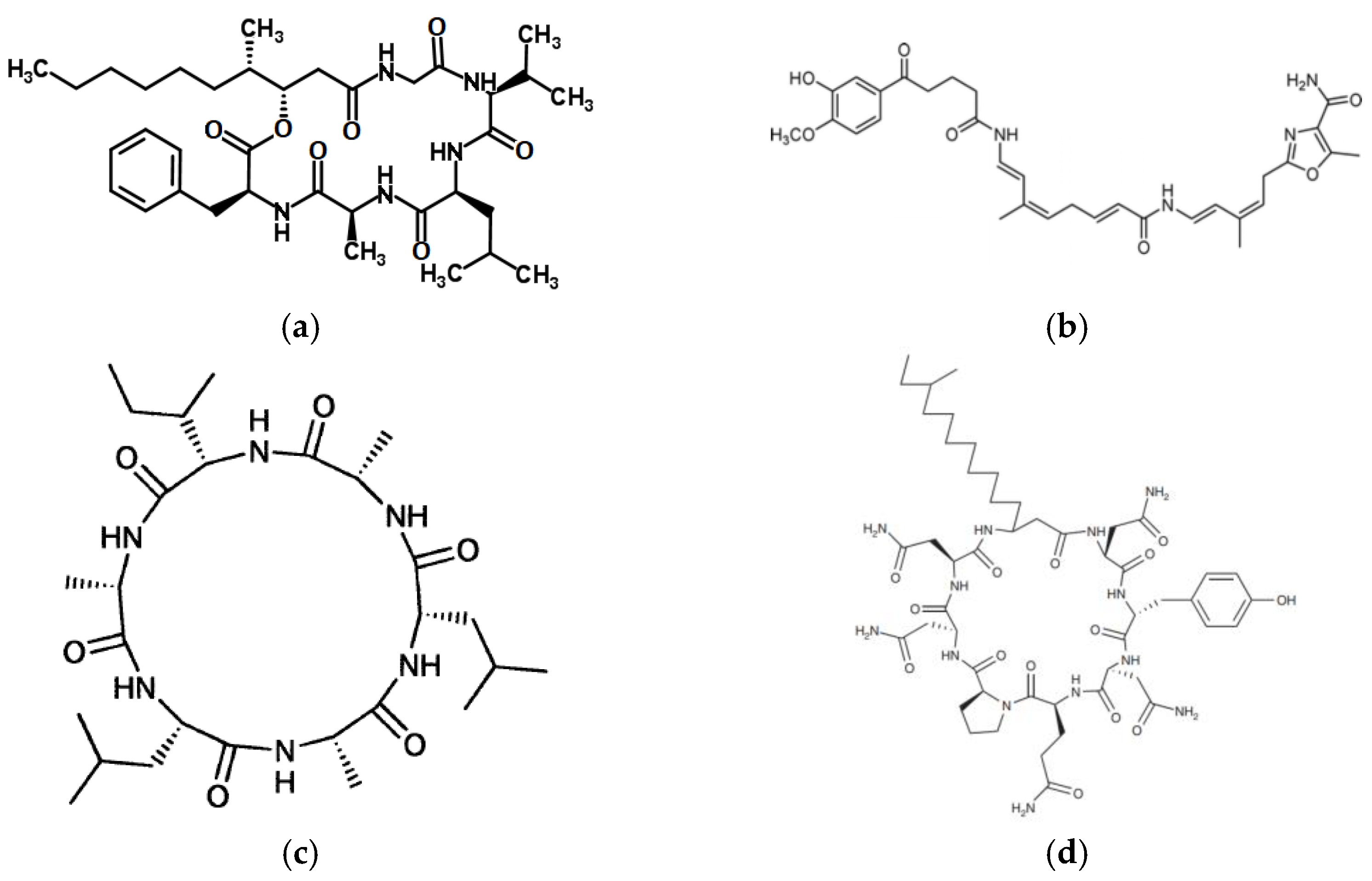
5. Nonribosomal Peptides (NRPs)
5.1. Arenamides
5.2. Ariakemicins
5.3. Halolitoralins
5.4. Heptapeptide from Paenibacillus profundus
5.5. Ieodoglucomides
5.6. Iturinic Lipopeptides
5.7. Lajollamycin
5.8. Lucentamycins
5.9. Mechercharmycins
5.10. Mixirins
5.11. Ohmyungsamycins
5.12. Padanamides
5.13. Piperazimycins
5.14. Proximicins
5.15. Urukthapelstatin A
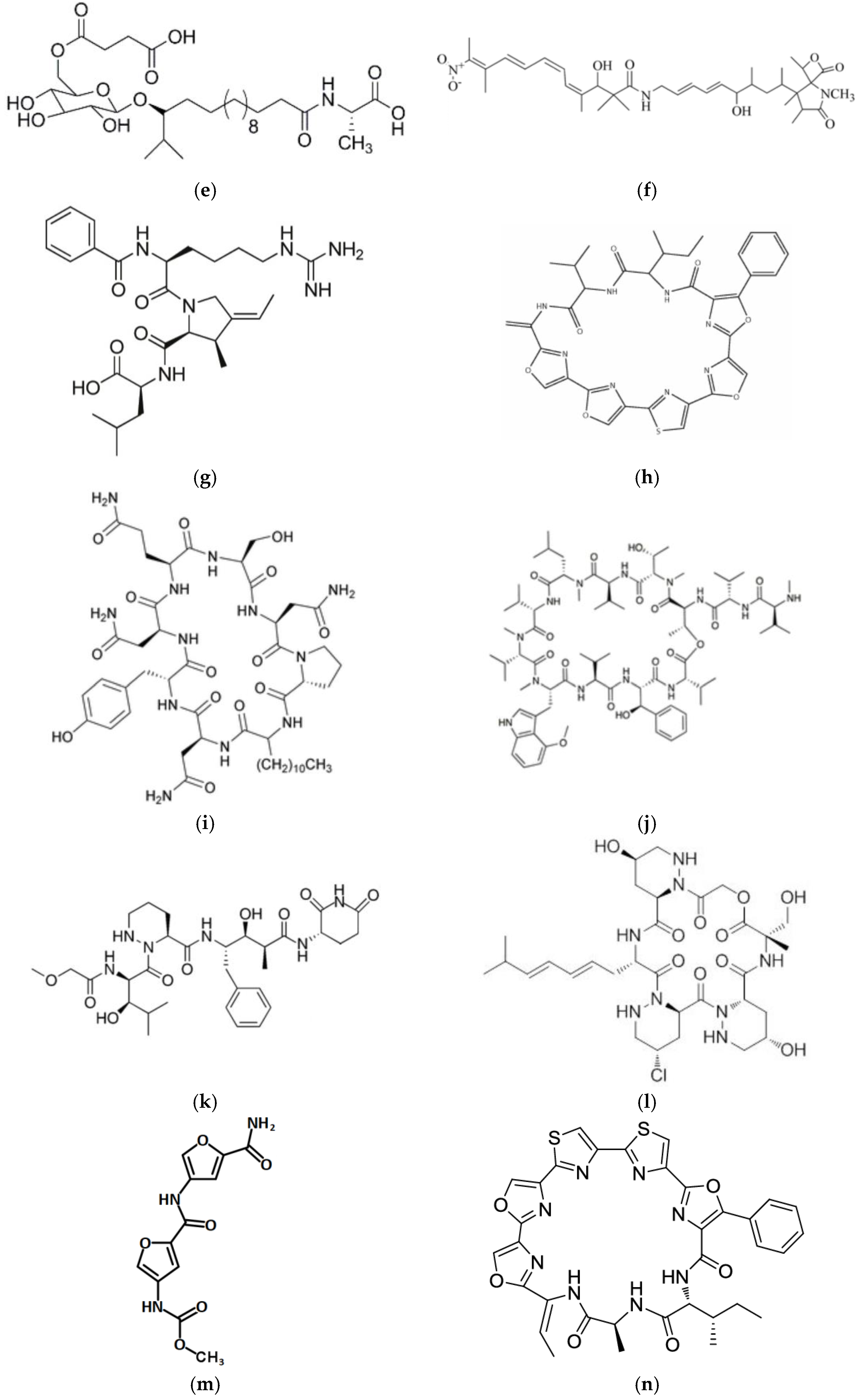
6. Toxins
6.1. Botulinum Neurotoxin Type A
6.2. Diphtheria Toxin
6.3. Exotoxin A
6.4. Listeriolysin O
7. Other Proteins/Peptides
7.1. Azurin
7.2. p28
7.3. Entap
7.4. Pep27anal2
8. Final Remarks
Author Contributions
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Ervik, M.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. GLOBOCAN 2012 v1.0, Cancer Incidence and Mortality Worldwide: IARC CancerBase No. 11; International Agency for Research on Cancer: Lyon, France, 2013. [Google Scholar]
- Worldwide Cancer Statistics. Available online: http://www.cancerresearchuk.org/health-professional/cancer-statistics/worldwide-cancer (accessed on 16 January 2018).
- Cancer. Available online: http://www.who.int/mediacentre/factsheets/fs297/en/ (accessed on 16 January 2018).
- Cancer Prevention and Control in the Context of an Integrated Approach. WHA70.12. Available online: http://apps.who.int/medicinedocs/documents/s23233en/s23233en.pdf (accessed on 16 January 2018).
- Langie, S.A.; Koppen, G.; Desaulniers, D.; Al-Mulla, F.; Al-Temaimi, R.; Amedei, A.; Azqueta, A.; Bisson, W.H.; Brown, D.G.; Brunborg, G.; et al. Causes of genome instability: The effect of low dose chemical exposures in modern society. Carcinogenesis 2015, 36 (Suppl. 1), S61–S88. [Google Scholar] [CrossRef] [PubMed]
- Kuno, T.; Tsukamoto, T.; Hara, A.; Tanaka, T. Cancer chemoprevention through the induction apoptosis by natural compounds. J. Biophys. Chem. 2012, 3, 156–173. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Raguz, S.; Yagüe, E. Resistance to chemotherapy: New treatments and novel insights into an old problem. Br. J. Cancer 2008, 99, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Coley, W.B. The treatment of inoperable sarcoma by bacterial toxins (the mixed toxins of the Streptococcus erysipelas and Bacillus prodigiosus). Proc. R. Soc. Med. Surg. 1909, 3, 1–48. [Google Scholar]
- Patyar, S.; Joshi, R.; Prasad Byrav, D.S.; Prakash, A.; Medhi, B.; Das, B.K. Bacteria in cancer therapy: A novel experimental strategy. J. Biomed. Sci. 2010, 17, 21. [Google Scholar] [CrossRef] [PubMed]
- Encyclopaedia Britannica. Available online: http://www.britannica.com/science/antibiotic (accessed on 20 February 2018).
- Barret, J.M.; Salles, B.; Provot, C.; Hill, B.T. Evaluation of DNA repair inhibition by antitumor or antibiotic drugs using a chemiluminescence microplate assay. Carcinogenesis 1997, 18, 2441–2445. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Pan, J.S.; Chang, W.C.; Hung, J.S.; Mao, S.J. The molecular mechanism of actinomycin D in preventing neointimal formation in rat carotid arteries after balloon injury. J. Biomed. Sci. 2005, 12, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Koba, M.; Konopa, J. Actinomycin D and its mechanisms of action. Postepy Hig. Med. Dosw. 2005, 59, 290–298. (In Polish) [Google Scholar]
- Cosmegen® for Injection. Available online: http://www.accessdata.fda.gov/drugsatfda_docs/label/2009/050682s025lbl.pdf (accessed on 20 February 2018).
- Segerman, Z.J.; Roy, B.; Hecht, S.M. Characterization of bleomycin-mediated cleavage of a hairpin DNA library. Biochemistry 2013, 52. [Google Scholar] [CrossRef] [PubMed]
- Bayer, R.A.; Gaynor, E.R.; Fisher, R.I. Bleomycin in non-Hodgkin’s lymphoma. Semin. Oncol. 1992, 19 (Suppl. 5), 46–53. [Google Scholar] [PubMed]
- Chen, J.K.; Yang, D.; Shen, B.; Murray, V. Bleomycin analogues preferentially cleave at the transcription start sites of actively transcribed genes in human cells. Int. J. Biochem. Cell Biol. 2017, 85, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Bleomycin. Genomics of Drug Sensitivity in Cancer. Available online: http://www.cancerrxgene.org/translation/Drug/190 (accessed on 14 April 2018).
- Abraham, S.A.; Waterhouse, D.N.; Mayer, L.D.; Cullis, P.R.; Madden, T.D.; Bally, M.B. The liposomal formulation of doxorubicin. Methods Enzymol. 2005, 391, 71–96. [Google Scholar] [CrossRef] [PubMed]
- Botlagunta, M.; Kollapalli, B.; Kakarla, L.; Gajarla, S.P.; Gade, S.P.; Dadi, C.L.; Penumadu, A.; Javeed, S. In vitro anti-cancer activity of doxorubicin against human RNA helicase, DDX3. Bioinformation 2016, 12, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Vittorio, O.; Le Grand, M.; Makharza, S.A.; Curcio, M.; Tucci, P.; Iemma, F.; Nicoletta, F.P.; Hampel, S.; Cirillo, G. Doxorubicin synergism and resistance reversal in human neuroblastoma BE(2)C cell lines: An in vitro study with dextran-catechin nanohybrids. Eur. J. Pharm. Biopharm. 2018, 122, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Doxorubicin Hydrochloride for Injection, USP. Available online: http://www.accessdata.fda.gov/drugsatfda_docs/label/2010/050467s070lbl.pdf (accessed on 20 February 2018).
- Mitomycin C from Streptomyces caespitosus. Available online: http://www.sigmaaldrich.com/content/dam/sigma-aldrich/docs/Sigma/Product_Information_Sheet/2/m0503pis.pdf (accessed on 20 February 2018).
- Bradner, W.T. Mitomycin C: A clinical update. Cancer Treat. Rev. 2001, 27, 35–50. [Google Scholar] [CrossRef] [PubMed]
- Mitomycin-C. Genomics of Drug Sensitivity in Cancer. Available online: http://www.cancerrxgene.org/translation/Drug/136 (accessed on 14 April 2018).
- Latta, V.D.; Cecchettini, A.; Del Ry, S.; Morales, M.A. Bleomycin in the setting of lung fibrosis induction: From biological mechanisms to counteractions. Pharmacol. Res. 2015, 97, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Farhane, Z.; Bonnier, F.; Byrne, H.J. An in vitro study of the interaction of the chemotherapeutic drug Actinomycin D with lung cancer cell lines using Raman micro-spectroscopy. J. Biophotonics 2018, 11, e201700112. [Google Scholar] [CrossRef] [PubMed]
- D’Arpa, P.; Liu, L.F. Topoisomerase—Targeting antitumor drugs. Biochim. Biophys. Acta 1989, 989, 163–177. [Google Scholar] [CrossRef]
- Dactinomycine. Available online: http://medycyna.anauk.net/101-0-393-.Encyklopedia.Lekow.html (accessed on 20 February 2018).
- Egger, C.; Cannet, C.; Gérard, C.; Jarman, E.; Jarai, G.; Feige, A.; Suply, T.; Micard, A.; Dunbar, A.; Tigani, B.; et al. Administration of bleomycin via the oropharyngeal aspiration route leads to sustained lung fibrosis in mice and rats as quantified by UTE-MRI and histology. PLoS ONE 2013, 8, e63432. [Google Scholar] [CrossRef] [PubMed]
- Blenoxane. Available online: http://www.accessdata.fda.gov/drugsatfda_docs/label/2010/050443s036lbl.pdf (accessed on 20 February 2018).
- Bleomycin for Injection USP. Available online: http://oncozine.com/wp-content/uploads/2017/07/Bleomycin-for-Injection-USP.pdf (accessed on 20 February 2018).
- Thorn, C.F.; Oshiro, C.; Marsh, S.; Hernandez-Boussard, T.; McLeod, H.; Klein, T.E.; Altman, R.B. Doxorubicin pathways: Pharmacodynamics and adverse effects. Pharmacogenet. Genom. 2011, 21, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Cagel, M.; Grotz, E.; Bernabeu, E.; Moretton, M.A.; Chiappetta, D.A. Doxorubicin: Nanotechnological overviews from bench to bedside. Drug Discov. Today 2017, 22, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Preet, S.; Bharati, S.; Panjeta, A.; Tewari, R.; Rishi, P. Effect of nisin and doxorubicin on DMBA-induced skin carcinogenesis—A possible adjunct therapy. Tumour Biol. 2015, 36, 8301–8308. [Google Scholar] [CrossRef] [PubMed]
- Doksorubicyna (Opis Profesjonalny). Available online: http://bazalekow.mp.pl/leki/doctor_subst.html?id=247 (accessed on 20 February 2018).
- Verweij, J.; Pinedo, H.M. Mitomycin C: Mechanism of action, usefulness and limitations. Anti-Cancer Drugs 1990, 1, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Mitomycin. Available online: http://bazalekow.mp.pl/leki/szukaj.html?item_name=mitomycin (accessed on 20 February 2018).
- Karpiński, T.M.; Szkaradkiewicz, A.K. Bacteriocins. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldra, F., Eds.; Elsevier, Academic Press: New York, NY, USA, 2016; Volume 1, pp. 312–319. [Google Scholar]
- Kaur, S.; Kaur, S. Bacteriocins as potential anticancer agents. Front. Pharmacol. 2015, 6, 272. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.M.; Pati, B.R.; Chakraborty, R.; Franco, O.L. New insights into the bioactivity of peptides from probiotics. Front. Biosci. 2016, 8, 450–459. [Google Scholar] [CrossRef]
- Drider, D.; Bendali, F.; Naghmouchi, K.; Chikindas, M.L. Bacteriocins: Not only antibacterial agents. Probiotics Antimicrob. Proteins 2016, 8, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Sieiro, P.; Montalbán-López, M.; Mu, D.; Kuipers, O.P. Bacteriocins of lactic acid bacteria: Extending the family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef] [PubMed]
- Gomes, K.M.; Duarte, R.S.; de Freire Bastos, M.D. Lantibiotics produced by Actinobacteria and their potential applications (a review). Microbiology 2017, 163, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Karpiński, T.M.; Szkaradkiewicz, A.K. Characteristic of bacteriocines and their application. Pol. J. Microbiol. 2013, 62, 223–235. [Google Scholar] [PubMed]
- Paiva, A.D.; de Oliveira, M.D.; de Paula, S.O.; Baracat-Pereira, M.C.; Breukink, E.; Mantovani, H.C. Toxicity of bovicin HC5 against mammalian cell lines and the role of cholesterol in bacteriocin activity. Microbiology 2012, 158, 2851–2858. [Google Scholar] [CrossRef] [PubMed]
- Chumchalova, J.; Smarda, J. Human tumor cells are selectively inhibited by colicins. Folia Microbiol. 2003, 48, 111–115. [Google Scholar] [CrossRef]
- Baindara, P.; Gautam, A.; Raghava, G.P.S.; Korpole, S. Anticancer properties of a defensin like class IId bacteriocin Laterosporulin10. Sci. Rep. 2017, 7, 46541. [Google Scholar] [CrossRef] [PubMed]
- Hetz, C.; Bono, M.R.; Barros, L.F.; Lagos, R. Microcin E492, a channel-forming bacteriocin from Klebsiella pneumoniae, induces apoptosis in some human cell lines. Proc. Natl. Acad. Sci. USA 2002, 99, 2696–2701. [Google Scholar] [CrossRef] [PubMed]
- Lagos, R.; Tello, M.; Mercado, G.; García, V.; Monasterio, O. Antibacterial and antitumorigenic properties of microcin E492, a pore-forming bacteriocin. Curr. Pharm. Biotechnol. 2009, 10, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Begde, D.; Bundale, S.; Mashitha, P.; Rudra, J.; Nashikkar, N.; Upadhyay, A. Immunomodulatory efficacy of nisin—A bacterial lantibiotic peptide. J. Pept. Sci. 2011, 17, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Joo, N.E.; Ritchie, K.; Kamarajan, P.; Miao, D.; Kapila, Y.L. Nisin, an apoptogenic bacteriocin and food preservative, attenuates HNSCC tumorigenesis via CHAC1. Cancer Med. 2012, 1, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Kamarajan, P.; Hayami, T.; Matte, B.; Liu, Y.; Danciu, T.; Ramamoorthy, A.; Worden, F.; Kapila, S.; Kapila, Y. Nisin ZP a bacteriocin and food preservative, inhibits head and neck cancer tumorigenesis and prolongs survival. PLoS ONE 2015, 10, e0131008. [Google Scholar] [CrossRef] [PubMed]
- Balgir, P.P.; Bhatia, P.; Kaur, B. Sequence analysis and homology based modeling to assess structure-function relationship of pediocin CP2 of Pediococcus acidilactici MTCC 5101. Ind. J. Biotechnol. 2010, 9, 431–434. [Google Scholar]
- Kumar, B.; Balgir, P.P.; Kaur, B.; Mittu, B.; Chauhan, A. In vitro cytotoxicity of native and rec-pediocin CP2 against cancer cell lines: A comparative study. Pharm. Anal. Acta 2012, 3, 1000183. [Google Scholar] [CrossRef]
- Villarante, K.I.; Elegado, F.B.; Iwatani, S.; Zendo, T.; Sonomoto, K.; de Guzman, E.E. Purification, characterization and in vitro cytotoxicity of the bacteriocin from Pediococcus acidilactici K2a2-3 against human colon adenocarcinoma (HT29) and human cervical carcinoma (HeLa) cells. World J. Microbiol. Biotechnol. 2011, 27, 975–980. [Google Scholar] [CrossRef]
- Zhao, H.; Sood, R.; Jutila, A.; Bose, S.; Fimland, G.; Nissen-Meyer, J.; Kinnunen, P.K. Interaction of the antimicrobial peptide pheromone plantaricin A with model membranes: Implications for a novel mechanism of action. Biochim. Biophys. Acta 2006, 1758, 1461–1474. [Google Scholar] [CrossRef] [PubMed]
- Abdi-Ali, A.; Worobec, E.A.; Deezagi, A.; Malekzadeh, F. Cytotoxic effects of pyocin S2 produced by Pseudomonas aeruginosa on the growth of three human cell lines. Can. J. Microbiol. 2004, 50, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Saito, H. Cytotoxicity of pyocin S2 to tumor and normal cells and its interaction with cell surfaces. Biochim. Biophys. Acta 1980, 633, 77–86. [Google Scholar] [CrossRef]
- Bactibase. Available online: http://bactibase.hammamilab.org (accessed on 12 February 2018).
- Uniprot. Available online: http://www.uniprot.org (accessed on 12 February 2018).
- Swiss-Model. Available online: http://swissmodel.expasy.org (accessed on 12 February 2018).
- Kiefer, F.; Arnold, K.; Künzli, M.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository and associated resources. Nucleic Acids Res. 2009, 37, D387–D392. [Google Scholar] [CrossRef] [PubMed]
- Smarda, J.; Fialova, M.; Smarda, J., Jr. Cytotoxic effects of colicins E1 and E3 on v-myb-transformed chicken monoblasts. Folia Biol. 2001, 47, 11–13. [Google Scholar]
- Michel-Briand, Y.; Baysse, C. The pyocins of Pseudomonas aeruginosa. Biochimie 2002, 84, 499–510. [Google Scholar] [CrossRef]
- Ni, Y.; Schwaneberg, U.; Sun, Z.H. Arginine deiminase, a potential anti-tumor drug. Cancer Lett. 2008, 261, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Glazer, E.S.; Piccirillo, M.; Albino, V.; Di Giacomo, R.; Palaia, R.; Mastro, A.A.; Beneduce, G.; Castello, G.; De Rosa, V.; Petrillo, A.; et al. Phase II study of pegylated arginine deiminase for nonresectable and metastatic hepatocellular carcinoma. J. Clin. Oncol. 2010, 28, 2220–2226. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.H.; Coates, J.M.; Bowles, T.L.; McNerney, G.P.; Sutcliffe, J.; Jung, J.U.; Gandour-Edwards, R.; Chuang, F.Y.S.; Bold, R.J.; Kung, H.-J. Arginine deiminase as a novel therapy for prostate cancer induces autophagy and caspase-independent apoptosis. Cancer Res. 2009, 69, 2. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, T.; Strauss, M.; Hering, S.; Redanz, U.; William, D.; Rosche, Y.; Classen, C.F.; Kreikemeyer, B.; Linnebacher, M.; Maletzki, C. Arginine deprivation by arginine deiminase of Streptococcus pyogenes controls primary glioblastoma growth in vitro and in vivo. Cancer Biol. Ther. 2015, 16, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Kaur, B.; Kaur, R. Purification of a dimeric arginine deiminase from Enterococcus faecium GR7 and study of its anti-cancerous activity. Protein Expr. Purif. 2016, 125, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Pritsa, A.A.; Papazisis, K.T.; Kortsaris, A.H.; Geromichalos, G.D.; Kyriakidis. Antitumor activity of l-asparaginase from Thermus thermophilus. Anticancer Drugs 2001, 12, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Avramis, V.I.; Sencer, S.; Periclou, A.P.; Sather, H.; Bostrom, B.C.; Cohen, L.J.; Ettinger, A.G.; Ettinger, L.J.; Franklin, J.; Gaynon, P.S.; et al. A randomized comparison of native Escherichia coli asparaginase and polyethylene glycol conjugated asparaginase for treatment of children with newly diagnosed standard-risk acute lymphoblastic leukemia: A Children’s Cancer Group study. Blood 2002, 99, 1986–1994. [Google Scholar] [CrossRef] [PubMed]
- Panosyan, E.H.; Wang, Y.; Xia, P.; Lee, W.N.; Pak, Y.; Laks, D.R.; Lin, H.J.; Moore, T.B.; Cloughesy, T.F.; Kornblum, H.I.; et al. Asparagine depletion potentiates the cytotoxic effect of chemotherapy against brain tumors. Mol. Cancer Res. 2014, 12, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Jaccard, A.; Gachard, N.; Marin, B.; Rogez, S.; Audrain, M.; Suarez, F.; Tilly, H.; Morschhauser, F.; Thieblemont, C.; Ysebaert, L.; et al. Efficacy of l-asparaginase with methotrexate and dexamethasone (AspaMetDex regimen) in patients with refractory or relapsing extranodal NK/T-cell lymphoma, a phase 2 study. Blood 2011, 117, 1834–1839. [Google Scholar] [CrossRef] [PubMed]
- Covini, D.; Tardito, S.; Bussolati, O.; Chiarelli, L.R.; Pasquetto, M.V.; Digilio, R.; Valentini, G.; Scotti, C. Expanding targets for a metabolic therapy of cancer: l-asparaginase. Recent Pat. Anti-Cancer Drug Discov. 2012, 7, 4–13. [Google Scholar] [CrossRef]
- Yu, M.; Henning, R.; Walker, A.; Kim, G.; Perroy, A.; Alessandro, R.; Virador, V.; Kohn, E.C. l-asparaginase inhibits invasive and angiogenic activity and induces autophagy in ovarian cancer. J. Cell. Mol. Med. 2012, 16, 2369–2378. [Google Scholar] [CrossRef] [PubMed]
- Oza, V.P.; Parmar, P.P.; Kumar, S.; Subramanian, R.B. Anticancer properties of highly purified l-asparaginase from Withania somnifera L. against acute lymphoblastic leukemia. Appl. Biochem. Biotechnol. 2010, 160, 1833–1840. [Google Scholar] [CrossRef] [PubMed]
- Meghavarnam, A.K.; Salah, M.; Sreepriya, M.; Janakiraman, S. Growth inhibitory and proapoptotic effects of l-asparaginase from Fusarium culmorum ASP-87 on human leukemia cells (Jurkat). Fundam. Clin. Pharmacol. 2017, 31, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Arjun, J.K.; Aneesh, B.P.; Kavitha, T.; Harikrishnan, K. Characterization of a novel asparaginase from soil metagenomic libraries generated from forest soil. Biotechnol. Lett. 2018, 40, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Takaku, H.; Takase, M.; Abe, S.; Hayashi, H.; Miyazaki, K. In vivo anti-tumor activity of arginine deiminase purified from Mycoplasma arginini. Int. J. Cancer 1992, 51, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Holtsberg, F.W.; Ensor, C.M.; Steiner, M.R.; Bomalaski, J.S.; Clark, M.A. Poly(ethylene glycol) (PEG) conjugated arginine deiminase: Effects of PEG formulations on its pharmacological properties. J. Controll. Release 2002, 80, 259–271. [Google Scholar] [CrossRef]
- Pieters, R.; Hunger, S.P.; Boos, J.; Rizzari, C.; Silverman, L.; Baruchel, A.; Goekbuget, N.; Schrappe, M.; Pui, C.H. l-asparaginase treatment in acute lymphoblastic leukemia: A focus on Erwinia asparaginase. Cancer 2011, 117, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.; Acharya, D.; Adholeya, A.; Barrow, C.J.; Deshmukh, S.K. Nonribosomal peptides from marine microbes and their antimicrobial and anticancer potential. Front. Pharmacol. 2017, 8, 828. [Google Scholar] [CrossRef] [PubMed]
- Sieber, S.A.; Marahiel, M.A. Learning from nature’s drug factories: Nonribosomal synthesis of macrocyclic peptides. J. Bacteriol. 2003, 185, 7036–7043. [Google Scholar] [CrossRef] [PubMed]
- Asolkar, R.N.; Freel, K.C.; Jensen, P.R.; Fenical, W.; Kondratyuk, T.P.; Park, E.-J.; Pezzuto, J.M. Arenamides A-C, cytotoxic NFκB inhibitors from the marine actinomycete Salinispora arenicola. J. Nat. Prod. 2009, 72, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Oku, N.; Adachi, K.; Matsuda, S.; Kasai, H.; Takatsuki, A.; Shizuri, Y. Ariakemicins A and B, novel polyketide-peptide antibiotics from a marine gliding bacterium of the genus Rapidithrix. Org. Lett. 2008, 10, 2481–2484. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Tan, R.-X.; Wang, Q.; Huang, W.-Y.; Yin, Y.-X. Antifungal cyclopeptides from Halobacillus litoralis YS3106 of marine origin. Tetrahedron Lett. 2002, 43, 6545–6548. [Google Scholar] [CrossRef]
- Kalinovskaya, N.I.; Romanenko, L.A.; Kalinovsky, A.I.; Dmitrenok, P.S.; Dyshlovoy, S.A. A new antimicrobial and anticancer peptide producing by the marine deep sediment strain “Paenibacillus profundus” sp. nov. Sl 79. Nat. Prod. Commun. 2013, 8, 381–384. [Google Scholar] [PubMed]
- Tareq, F.S.; Kim, J.H.; Lee, M.A.; Lee, H.-S.; Lee, Y.-J.; Lee, J.S.; Shin, H.J. Ieodoglucomides A and B from a marine-derived bacterium Bacillus licheniformis. Org. Lett. 2012, 14, 1464–1467. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Wang, N.; Hu, J.; Wang, S. Isolation and characterization of a new iturinic lipopeptide, mojavensin A produced by a marine-derived bacterium Bacillus mojavensis B0621A. J. Antibiot. 2012, 65, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Rama, M.R.; Teisan, S.; White, D.J.; Nicholson, B.; Grodberg, J.; Neuteboom, S.T.C.; Lam, K.S.; Mosca, D.A.; Lloyd, K.G.; Potts, B.C.M. Lajollamycin, a nitro-tetraene spiro-β-lactone-γ-lactam antibiotic from the marine actinomycete Streptomyces nodosus. J. Nat. Prod. 2005, 68, 240–243. [Google Scholar] [CrossRef]
- Cho, J.Y.; Williams, P.G.; Kwon, H.C.; Jensen, P.R.; Fenical, W. Lucentamycins A-D, cytotoxic peptides from the marine-derived actinomycete Nocardiopsis lucentensis. J. Nat. Prod. 2007, 70, 1321–1328. [Google Scholar] [CrossRef] [PubMed]
- Kanoh, K.; Matsuo, Y.; Adachi, K.; Imagawa, H.; Nishizawa, M.; Shizuri, Y. Mechercharmycins A and B, cytotoxic substances from marine-derived Thermoactinomyces sp. YM3-251. J. Antibiot. 2005, 58, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.L.; Hua, H.M.; Pei, Y.H.; Yao, X.S. Three new cytotoxic cyclic acylpeptides from marine Bacillus sp. Chem. Pharm. Bull. 2004, 52, 1029–1030. [Google Scholar] [CrossRef] [PubMed]
- Um, S.; Choi, T.J.; Kim, H.; Kim, B.Y.; Kim, S.-H.; Lee, S.K.; Oh, K.-B.; Shin, J.; Oh, D.-C. Ohmyungsamycins A and B: Cytotoxic and antimicrobial cyclic peptides produced by Streptomyces sp. from a volcanic island. J. Org. Chem. 2013, 78, 12321–12329. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.E.; Dalisay, D.S.; Patrick, B.O.; Matainaho, T.; Andrusiak, K.; Deshpande, R.; Myers, C.L.; Piotrowski, J.S.; Boone, C.; Yoshida, M.; et al. Padanamides A and B, highly modified linear tetrapeptides produced in culture by a Streptomyces sp. isolated from a marine sediment. Org. Lett. 2011, 13, 3936–3939. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.D.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Piperazimycins: Cytotoxic hexadepsipeptides from a marine-derived bacterium of the genus Streptomyces. J. Org. Chem. 2007, 72, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, H.-P.; Bruntner, C.; Riedlinger, J.; Bull, A.T.; Knutsen, G.; Goodfellow, M.; Jones, A.; Maldonado, L.; Pathom-aree, W.; Beil, W.; et al. Proximicin A, B and C, novel aminofuran antibiotic and anticancer compounds isolated from marine strains of the actinomycete Verrucosispora. J. Antibiot. 2008, 61, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Kanoh, K.; Yamori, T.; Kasai, H.; Katsuta, A.; Adachi, K.; Shin-Ya, K.; Shizuri, Y. Urukthapelstatin A, a novel cytotoxic substance from marine-derived Mechercharimyces asporophorigenens YM11-542. I. Fermentation, isolation and biological activities. J. Antibiot. 2007, 60, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Kanoh, K.; Imagawa, H.; Adachi, K.; Nishizawa, M.; Shizuri, Y. Urukthapelstatin A, a novel cytotoxic substance from marine-derived Mechercharimyces asporophorigenens YM11-542. II. Physico-chemical properties and structural elucidation. J. Antibiot. 2007, 60, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Henkel, J.S.; Baldwin, M.R.; Barbieri, J.T. Toxins from bacteria. EXS 2010, 100, 1–29. [Google Scholar] [PubMed]
- Purkiss, J.R.; Friis, L.M.; Doward, S.; Quinn, C.P. Clostridium botulinum neurotoxins act with a wide range of potencies on SH-SY5Y human neuroblastoma cells. Neurotoxicology 2001, 22, 447–453. [Google Scholar] [CrossRef]
- Karsenty, G.; Rocha, J.; Chevalier, S.; Scarlata, E.; Andrieu, C.; Zouanat, F.Z.; Rocchi, P.; Giusiano, S.; Elzayat, E.A.; Corcos, J. Botulinum toxin type A inhibits the growth of LNCaP human prostate cancer cells in vitro and in vivo. Prostate 2009, 69, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Proietti, S.; Nardicchi, V.; Porena, M.; Giannantoni, A. Botulinum toxin type-A toxin activity on prostate cancer cell lines. Urologia 2012, 79, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Bandala, C.; Perez-Santos, J.L.; Lara-Padilla, E.; Delgado Lopez, G.; Anaya-Ruiz, M. Effect of botulinum toxin A on proliferation and apoptosis in the T47D breast cancer cell line. Asian Pac. J. Cancer Prev. 2013, 14, 891–894. [Google Scholar] [CrossRef] [PubMed]
- Holmes, R.K. Biology and molecular epidemiology of diphtheria toxin and the tox gene. J. Infect. Dis. 2000, 181 (Suppl. 1), S156–S167. [Google Scholar] [CrossRef] [PubMed]
- Martarelli, D.; Pompei, P.; Mazzoni, G. Inhibition of adrenocortical carcinoma by diphtheria toxin mutant CRM197. Chemotherapy 2009, 55, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Vallera, D.A.; Li, C.; Jin, N.; Panoskaltsis-Mortari, A.; Hall, W.A. Targeting urokinase-type plasminogen activator receptor on human glioblastoma tumors with diphtheria toxin fusion protein DTAT. J. Natl. Cancer Inst. 2002, 94, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Lutz, M.B.; Baur, A.S.; Schuler-Thurner, B.; Schuler, G. Immunogenic and tolerogenic effects of the chimeric IL-2-diphtheria toxin cytocidal agent Ontak® on CD25+ cells. Oncoimmunology 2014, 3, e28223. [Google Scholar] [CrossRef] [PubMed]
- Shafiee, F.; Rabbani, M.; Jahanian-Najafabadi, A. Production and evaluation of cytotoxic effects of DT386-BR2 fusion protein as a novel anti-cancer agent. J. Microbiol. Methods 2016, 130, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.J.; Dao, H., Jr.; Nagarajan, P.; Duvic, M. Primary cutaneous anaplastic large-cell lymphoma: Complete remission for 13 years after denileukin diftitox. JAAD Case Rep. 2017, 3, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Alderson, R.F.; Kreitman, R.J.; Chen, T.; Yeung, P.; Herbst, R.; Fox, J.A.; Pastan, I. CAT-8015: A second-generation Pseudomonas exotoxin A-based immunotherapy targeting CD22-expressing hematologic malignancies. Clin. Cancer Res. 2009, 15, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Karpiński, T.M.; Szkaradkiewicz, A.K. Anticancer peptides from bacteria. Bangladesh J. Pharmacol. 2013, 8, 343–348. [Google Scholar] [CrossRef]
- Oh, S.; Todhunter, D.A.; Panoskaltsis-Mortari, A.; Buchsbaum, D.J.; Toma, S.; Vallera, D.A. A deimmunized bispecific ligand-directed toxin that shows an impressive anti-pancreatic cancer effect in a systemic nude mouse orthotopic model. Pancreas 2012, 41, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Risberg, K.; Fodstad, O.; Andersson, Y. Synergistic anticancer effects of the 9.2.27PE immunotoxin and ABT-737 in melanoma. PLoS ONE 2011, 6, e24012. [Google Scholar] [CrossRef] [PubMed]
- Waldron, N.N.; Kaufman, D.S.; Oh, S.; Inde, Z.; Hexum, M.K.; Ohlfest, J.R.; Vallera, D.A. Targeting tumor-initiating cancer cells with dCD133KDEL shows impressive tumor reductions in a xenotransplant model of human head and neck cancer. Mol. Cancer Ther. 2011, 10, 1829–1838. [Google Scholar] [CrossRef] [PubMed]
- Kerr, D.E.; Wu, G.Y.; Wu, C.H.; Senter, P.D. Listeriolysin O potentiates immunotoxin and bleomycin cytotoxicity. Bioconjug. Chem. 1997, 8, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Provoda, C.J.; Stier, E.M.; Lee, K.D. Tumor cell killing enabled by listeriolysin O-liposome-mediated delivery of the protein toxin gelonin. J. Biol. Chem. 2003, 278, 35102–35108. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Boland, J.A.; Kuhn, M.; Berche, P.; Chakraborty, T.; Domínguez-Bernal, G.; Goebel, W.; González-Zorn, B.; Wehland, J.; Kreft, J. Listeria pathogenesis and molecular virulence determinants. Clin. Microbiol. Rev. 2001, 14, 584–640. [Google Scholar] [CrossRef] [PubMed]
- Bergelt, S.; Frost, S.; Lilie, H. Listeriolysin O as cytotoxic component of an immunotoxin. Protein Sci. 2009, 18, 1210–1220. [Google Scholar] [CrossRef] [PubMed]
- Stachowiak, R.; Lyzniak, M.; Budziszewska, B.K.; Roeske, K.; Bielecki, J.; Hoser, G.; Kawiak, J. Cytotoxicity of bacterial metabolic products, including listeriolysin O, on leukocyte targets. J. Biomed. Biotechnol. 2012, 2012, 954375. [Google Scholar] [CrossRef] [PubMed]
- Goto, M.; Yamada, T.; Kimbara, K.; Horner, J.; Newcomb, M.; Gupta, T.K.; Chakrabarty, A.M. Induction of apoptosis in macrophages by Pseudomonas aeruginosa azurin: Tumour-suppressor protein p53 and reactive oxygen species, but not redox activity, as critical elements in cytotoxicity. Mol. Microbiol. 2003, 47, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Zhou, J.; Su, Z.; Huang, Y. Bacterial cupredoxin azurin hijacks cellular signaling networks: Protein-protein interactions and cancer therapy. Protein Sci. 2017, 26, 2334–2341. [Google Scholar] [CrossRef] [PubMed]
- Punj, V.; Bhattacharyya, S.; Saint-Dic, D.; Vasu, C.; Cunningham, E.A.; Graves, J.; Yamada, T.; Constantinou, A.I.; Christov, K.; White, B.; et al. Bacterial cupredoxin azurin as an inducer of apoptosis and regression in human breast cancer. Oncogene 2004, 23, 2367–2378. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-H.; Lee, M.-H.; Cho, Y.-J.; Park, B.S.; Kim, S.; Kim, G.C. The bacterial protein azurin enhances sensitivity of oral squamous carcinoma cells to anticancer drugs. Yonsei Med. J. 2011, 52, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Goto, M.; Punj, V.; Zaborina, O.; Chen, M.L.; Kimbara, K.; Majumdar, D.; Cunningham, E.; Das Gupta, T.K.; Chakrabarty, A.M. Bacterial redox protein azurin, tumor suppressor protein p53, and regression of cancer. Proc. Natl. Acad. Sci. USA 2002, 99, 14098–14103. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, N.; Chakrabarty, A.M.; Fialho, A.M. Engineering of bacterial strains and their products for cancer therapy. Appl. Microbiol. Biotechnol. 2013, 97, 5189–5199. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Mehta, R.R.; Lekmine, F.; Christov, K.; King, M.L.; Majumdar, D.; Shilkaitis, A.; Green, A.; Bratescu, L.; Beattie, C.W.; et al. A peptide fragment of azurin induces a p53-mediated cell cycle arrest in human breast cancer cells. Mol. Cancer Ther. 2009, 8, 2947–2958. [Google Scholar] [CrossRef] [PubMed]
- Mehta, R.R.; Yamada, T.; Taylor, B.N.; Christov, K.; King, M.L.; Majumdar, D.; Lekmine, F.; Tiruppathi, C.; Shilkaitis, A.; Bratescu, L.; et al. A cell penetrating peptide derived from azurin inhibits angiogenesis and tumor growth by inhibiting phosphorylation of VEGFR-2, FAK and Akt. Angiogenesis 2011, 14, 355–369. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Das Gupta, T.K.; Beattie, C.W. p28-mediated activation of p53 in G2-M phase of the cell cycle enhances the efficacy of DNA damaging and antimitotic chemotherapy. Cancer Res. 2016, 76, 2354–2365. [Google Scholar] [CrossRef] [PubMed]
- CDG. Available online: http://www.cdgti.com/clinical-trials/ (accessed on 14 February 2018).
- Karpiński, T.M. New Peptide (Entap) with Anti-Proliferative Activity Produced by Bacteria of Enterococcus Genus. Habilitation Thesis, Scientific Publisher of Poznań University of Medical Sciences, Poznan, Poland, 2012. (In Polish). [Google Scholar]
- Karpiński, T.M.; Szkaradkiewicz, A.; Gamian, A. New enterococcal anticancer peptide. In Proceedings of the 23rd European Congress of Clinical Microbiology and Infectious Diseases, Berlin, Germany, 27–30 April 2013. [Google Scholar]
- Sung, W.S.; Park, Y.; Choi, C.H.; Hahm, K.S.; Lee, D.G. Mode of antibacterial action of a signal peptide, Pep27 from Streptococcus pneumoniae. Biochem. Biophys. Res. Commun. 2007, 363, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Hahm, K.-S.; Park, Y.; Kim, H.Y.; Lee, W.; Lim, S.C.; Seo, Y.K.; Choi, C.H. Functional and structural characteristics of anticancer peptide Pep27 analogues. Cancer Cell Int. 2005, 5, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- I-TASSER. Available online: http://zhanglab.ccmb.med.umich.edu/I-TASSER/ (accessed on 16 February 2018).
- Pangestuti, R.; Kim, S.-K. Bioactive peptide of marine origin for the prevention and treatment of non-communicable diseases. Mar. Drugs 2017, 15, 67. [Google Scholar] [CrossRef] [PubMed]
- Vijaya, T. Anticancer, antimicrobial and antioxidant bioactive factors derived from marine fungal endophytes; a review. Indo Am. J. Pharm. Res. 2017, 7, 7313–7321. [Google Scholar]
- Guzmán-Rodríguez, J.J.; Ochoa-Zarzosa, A.; López-Gómez, R.; López-Meza, J.E. Plant antimicrobial peptides as potential anticancer agents. BioMed Res. Int. 2015, 2015, 735087. [Google Scholar] [CrossRef] [PubMed]
- Negi, B.; Kumar, D.; Rawat, D.S. Marine peptides as anticancer agents: A remedy to mankind by nature. Curr. Protein Pept. Sci. 2017, 18, 885–904. [Google Scholar] [CrossRef] [PubMed]
- Karpiński, T.M.; Szkaradkiewicz, A.K. Human defensins. Arch. Biomed. Sci. 2013, 1, 1–5. [Google Scholar]
- Deslouches, B.; Di, P.Y. Antimicrobial peptides with selective antitumor mechanisms: Prospect for anticancer applications. Oncotarget 2017, 8, 46635–46651. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Chen, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Dermaseptin-PH: A novel peptide with antimicrobial and anticancer activities from the skin secretion of the south american orange-legged leaf frog, Pithecopus (Phyllomedusa) hypochondrialis. Molecules 2017, 22, 1805. [Google Scholar] [CrossRef] [PubMed]
- Dykhuizen, D. Species numbers in bacteria. Proc. Calif. Acad. Sci. 2005, 56 (Suppl. 1), 62–71. [Google Scholar] [PubMed]
- Schloss, P.D.; Girard, R.A.; Martin, T.; Edwards, J.; Thrash, J.C. Status of the archaeal and bacterial census: An update. mBio 2016, 7, E00201-16. [Google Scholar] [CrossRef] [PubMed]
- Cheung, R.C.; Ng, T.B.; Wong, J.H. Marine peptides: Bioactivities and applications. Mar. Drugs 2015, 13, 4006–4043. [Google Scholar] [CrossRef] [PubMed]
- Daliri, E.B.-M.; Oh, D.H.; Lee, B.H. Bioactive peptides. Foods 2017, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.M.; Gwak, J.W.; Kamarajan, P.; Fenno, J.C.; Rickard, A.H.; Kapila, Y.L. Biomedical applications of nisin. J. Appl. Microbiol. 2016, 120, 1449–1465. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, M.; Moridnia, A.; Mortazavi, D.; Salehi, M.; Bagheri, M.; Sheikhi, A. Kefir: A powerful probiotics with anticancer properties. Med. Oncol. 2017, 34, 183. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Torres, V.; Encinar, J.A.; Herranz-López, M.; Pérez-Sánchez, A.; Galiano, V.; Barrajón-Catalán, E.; Micol, V. An updated review on marine anticancer compounds: The use of virtual screening for the discovery of small-molecule cancer drugs. Molecules 2017, 22, 1037. [Google Scholar] [CrossRef] [PubMed]
- Gilad, Y.; Firer, M.; Gellerman, G. Recent innovations in peptide based targeted drug delivery to cancer cells. Biomedicines 2016, 4, 11. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Protein/Peptide | Source | Biological Target: Human Cancer Cells | IC50 | References |
|---|---|---|---|---|---|
| 1 | Actinomycin D | Actinomyces antibioticus | Wilms cancer, Ewing sarcoma, neuroblastomas, trophoblastic tumours | from 0.4 nM to 0.42 µM | [12,13,14,15] |
| 2 | Bleomycin | Streptomyces verticillus | head and neck squamous cell carcinomas, Hodgkin’s disease, non-Hodgkin’s lymphoma, testicular carcinomas, ovarian cancer, malignant pleural effusion | from 25.2 nM to 2.93 mM | [12,16,17,18,19] |
| 3 | Doxorubicin | Streptomyces peucetius var. caesius | acute lymphoblastic leukaemia, acute myeloblastic leukaemia, Wilms’ tumour, neuroblastoma, soft tissue and bone sarcomas, breast carcinoma, ovarian carcinoma, transitional cell bladder carcinoma, thyroid carcinoma, gastric carcinoma, Hodgkin’s disease, malignant lymphoma, bronchogenic carcinoma, oral squamous carcinoma | from 548.2 nM to 44.7 µM | [12,20,21,22,23] |
| 4 | Mitomycin C | Streptomyces caespitosus | cancers of the head and neck, lungs, breast, cervix, bladder, colorectal and anal carcinomas, hepatic cell carcinoma, melanoma, stomach and pancreatic carcinomas | from 9.48 nM to 249 µM | [12,24,25,26] |
| No. | Protein/Peptide | Source | Biological Target: Human Cancer Cell Lines | IC50 | References |
|---|---|---|---|---|---|
| 1 | Bovicin HC5 | Streptococcus bovis HC5 | breast adenocarcinoma (MCF-7), liver hepatocellular carcinoma (HepG2) | 279.4–289.3 µM | [41,47] |
| 2 | Colicins A and E1 | Escherichia coli | breast carcinoma (MCF7, ZR75, BT549, BT474, MDA-MB-231, SKBR3, T47D), osteosarcoma (HOS), leiomyosarcoma (SKUT-1), fibrosarcoma (HS913T) | n.d. | [41,48] |
| 3 | Laterosporulin 10 | Brevibacillus sp. strain SKDU10 | cervical cancer (HeLa), embryonic kidney cancer (HEK293T), fibrosarcoma (HT1080), lung carcinoma (H1299) breast cancer (MCF-7) | n.d. | [49] |
| 4 | Microcin E492 | Klebsiella pneumoniae RYC492 | cervical adenocarcinoma (HeLa), acute T cell leukaemia (Jurkat), Burkitt’s lymphoma (Ramos), B-lymphoblastoid cells (RJ2.25) | n.d. | [50,51] |
| 5 | Nisin A | Lactococcus lactis | head and neck squamous cell carcinoma (UM-SCC-17B, UM-SCC-14A, HSC-3), breast adenocarcinoma (MCF-7), liver hepatocellular carcinoma (HepG2), acute T cell leukaemia (Jurkat) | 105.5–225 µM | [40,47,52,53] |
| 6 | Nisin ZP | Lactococcus lactis | head and neck squamous cell carcinoma (UM-SCC-17B, HSC-3) | n.d. | [54] |
| 7 | Pediocin CP2 | Pediococcus acidilactici MTCC 5101 | mammary gland adenocarcinoma (MCF-7), hepatocarcinoma (Hep G2), cervical adenocarcinoma (HeLa) | n.d. | [55,56] |
| 8 | Pediocin K2a2-3 | Pediococcus acidilactici K2a2-3 | colon adenocarcinoma (HT29) | n.d. | [57] |
| 9 | Plantaricin A | Lactobacillus plantarum C11 | T cell leukaemia (Jurkat) | n.d. | [58] |
| 10 | Pyocin S2 | Pseudomonas aeruginosa 42A | hepatocellular carcinoma (HepG2), multiple myeloma (Im9), cervical adenocarcinoma (HeLa), embryonal ovary carcinoma (AS-II) | n.d. | [59,60] |
| No. | Protein/Peptide | Source | Biological Target: Human Cancer Cells/Cell Lines | IC50 | References |
|---|---|---|---|---|---|
| 1 | Arginine deiminase | Mycoplasma hominis, M. arginini | hepatocellular carcinoma (HCC), prostate cancer (CWR22Rv1), glioblastoma (HROG02, HROG05, HROG10, HROG17) | 1.95 µg/mL | [67,68,69,70,71] |
| 2 | l-asparaginase | Escherichia coli, Erwinia sp. | paediatric medulloblastoma (DAOY), glioblastomas (GBM-ES, U87), acute lymphoblastic leukaemia (ALL, HL60, MOLT-3, MOLT-4), myeloblastic leukaemia, acute T cell leukaemia (Jurkat), Hodgkin and non-Hodgkin lymphomas, myelosarcoma, multiple myeloma, extranodal NK/T cell lymphoma, ovarian carcinomas | 0.39–90 µg/mL | [72,73,74,75,76,77,78,79,80] |
| No. | Protein/Peptide | Source | Biological Target: Cancer Cells | IC50 | References |
|---|---|---|---|---|---|
| 1 | Arenamides A, B | Salinispora arenicola | human colon carcinoma (HCT-116) | 1.7–3.7 µM | [86] |
| 2 | Ariakemicins A, B | Rapidithrix sp. | human lung cancer (A549) | 25.4–42.3 µM | [87] |
| 3 | Halolitoralins A–C | Halobacillus litoralis YS3106 | human gastric tumour (BGC) | n.d. | [88] |
| 4 | Heptapeptide | Paenibacillus profundus | human melanoma (SK-MEL-28) | 3.07 µM | [89] |
| 5 | Ieodoglucomide B | Bacillus licheniformis | human lung cancer, human stomach cancer | n.d. | [90] |
| 6 | Iso-C16 fengycin B, anteiso-C17 fengycin B, mojavensin A | Bacillus mojavensis B0621A | human leukaemia (HL-60) | 1.6–100 mM | [91] |
| 7 | Lajollamycin | Streptomyces nodosus NPS007994 | mouse melanoma (B16-F10) | n.d. | [92] |
| 8 | Lucentamycins A, B | Nocardiopsis lucentensis CNR-712 | human colon carcinoma cells (HCT-116) | 0.2–11 µM | [93] |
| 9 | Mechercharmycin A | Thermoactinomyces sp. YM3-251 | human lung cancer cells (A549), human leukaemia (Jurkat) | 400–460 µM | [94] |
| 10 | Mixirins A–C | Bacillus sp. | human colon tumour (HCT-116) | 0.65–1.6 µM | [95] |
| 11 | Ohmyungsamycins A and B | Streptomyces sp. | diverse cancer cells | from 359 nM to 16.8 µM | [96] |
| 12 | Padanamide A, B | Streptomyces sp. | human leukaemia (Jurkat) | 30.9–90.7 µM | [97] |
| 13 | Piperazimycins A–C | Streptomyces sp. | multiple tumour cell lines | n.d. | [98] |
| 14 | Proximicins A–C | Verrucosispora sp. MG-37 and AB-18-032 | human gastric adenocarcinoma (AGS), human hepatocellular carcinoma (HepG2), human breast carcinoma (MCF 7) | n.d. | [99] |
| 15 | Urukthapelstatin A | Mechercharimyces asporophorigenens YM11-542 | human lung cancers (A549, DMS114, NCIH460), ovarian cancers (OVCAR-3, OVCAR-4, OVCAR-5, OVCAR-8, SK-OV3), breast cancer (MCF-7), colon cancer (HCT-116) | 12 nM | [100,101] |
| No. | Protein/Peptide | Source | Biological Target: Human Cancer Cell Lines | IC50 | References |
|---|---|---|---|---|---|
| 1 | Botulinum neurotoxin type A | Clostridium botulinum | prostate cancer (PC-3, LNCaP), breast cancer (T47D), neuroblastoma (SH-SY5Y) | 0.54–300 nM | [103,104,105,106] |
| 2 | Diphtheria toxin | Corynebacterium diphtheriae | adrenocortical carcinoma (H295R), glioblastomas (U118MG, U373MG, U87MG), cutaneous T cell lymphomas (CTCL), breast carcinoma (MCF 7), cervical adenocarcinoma (HeLa) | 0.55–2.08 µg/mL | [107,108,109,110,111,112] |
| 3 | Exotoxin A | Pseudomonas aeruginosa | pancreatic cancer (PaCa-2), melanomas (FEMX, Melmet-1, Melmet-5, Melmet-44, MelRM, MM200), head and neck squamous carcinomas, Burkitt’s lymphoma (Daudi, CA46), leukemias (EHEB, MEC1) | 0.3–8.6 ng/mL | [113,114,115,116,117] |
| 4 | Listeriolysin O | Listeria monocytogenes | breast carcinomas (MCF7, SKBR-3), leukemia T-lymphocytes (Jurkat) | from 50 pM to 0.1 nM, in conjugates | [118,119,120,121,122] |
| No. | Protein/Peptide | Source | Biological Target: Human Cancer Cells/Cell Lines | IC50 | References |
|---|---|---|---|---|---|
| 1 | Azurin | Pseudomonas aeruginosa | breast cancer (MCF-7, MDA-MB-157), oral squamous carcinoma (YD-9), melanoma (UISO-Mel-2) | 32–53 µM | [123,124,125,126,127] |
| 2 | p28 | Pseudomonas aeruginosa | breast cancer (MCF-7, ZR-75-1, T47D), glioblastoma (U87, LN229), melanoma (Mel-29), brain tumors | n.d. | [128,129,130,131,132] |
| 3 | Entap | Enterococcus sp. | gastric adenocarcinoma (AGS), uterine cervix adenocarcinoma (HeLa), mammary gland adenocarcinoma (MDA-MB-231), prostate carcinoma (22Rv1), colorectal adenocarcinoma (HT-29) | n.d. | [114,133,134] |
| 4 | Pep27anal2 | Streptococcus pneumoniae | leukemia (AML-2, HL-60, Jurkat), gastric cancer (SNU-601), breast cancer (MCF-7) | 10–29 µM | [135,136] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karpiński, T.M.; Adamczak, A. Anticancer Activity of Bacterial Proteins and Peptides. Pharmaceutics 2018, 10, 54. https://doi.org/10.3390/pharmaceutics10020054
Karpiński TM, Adamczak A. Anticancer Activity of Bacterial Proteins and Peptides. Pharmaceutics. 2018; 10(2):54. https://doi.org/10.3390/pharmaceutics10020054
Chicago/Turabian StyleKarpiński, Tomasz M., and Artur Adamczak. 2018. "Anticancer Activity of Bacterial Proteins and Peptides" Pharmaceutics 10, no. 2: 54. https://doi.org/10.3390/pharmaceutics10020054
APA StyleKarpiński, T. M., & Adamczak, A. (2018). Anticancer Activity of Bacterial Proteins and Peptides. Pharmaceutics, 10(2), 54. https://doi.org/10.3390/pharmaceutics10020054