Intestinal Drug Absorption Enhancement by Aloe vera Gel and Whole Leaf Extract: In Vitro Investigations into the Mechanisms of Action

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Chemical Characterisation of the A. vera Gel and the Whole-Leaf Extract
2.3. Chemical Characterisation of N-Trimethyl Chitosan Chloride
2.4. Caco-2 Cell Culturing
2.5. Cell Monolayer Integrity
2.6. In Vitro Transepithelial Electrical Resistance (TEER) Study
2.7. In Vitro Permeation Studies
2.8. Caco-2 Cell Monolayers for the Confocal Laser Scanning Microscopy (CLSM) Study
2.8.1. Fluorescence Staining
Visualisation of the Transport Pathway
Visualization of the F-Actin Filaments in the Cytoskeleton
2.8.2. Preparation of the Microscope Slides for the Confocal Laser Scanning Microscopy (CLSM)
2.8.3. Imaging with Confocal Laser Scanning Microscopy
2.9. Data Analysis
3. Results and Discussion
3.1. Characterisation of the A. vera Gel and the Whole-Leaf Extract
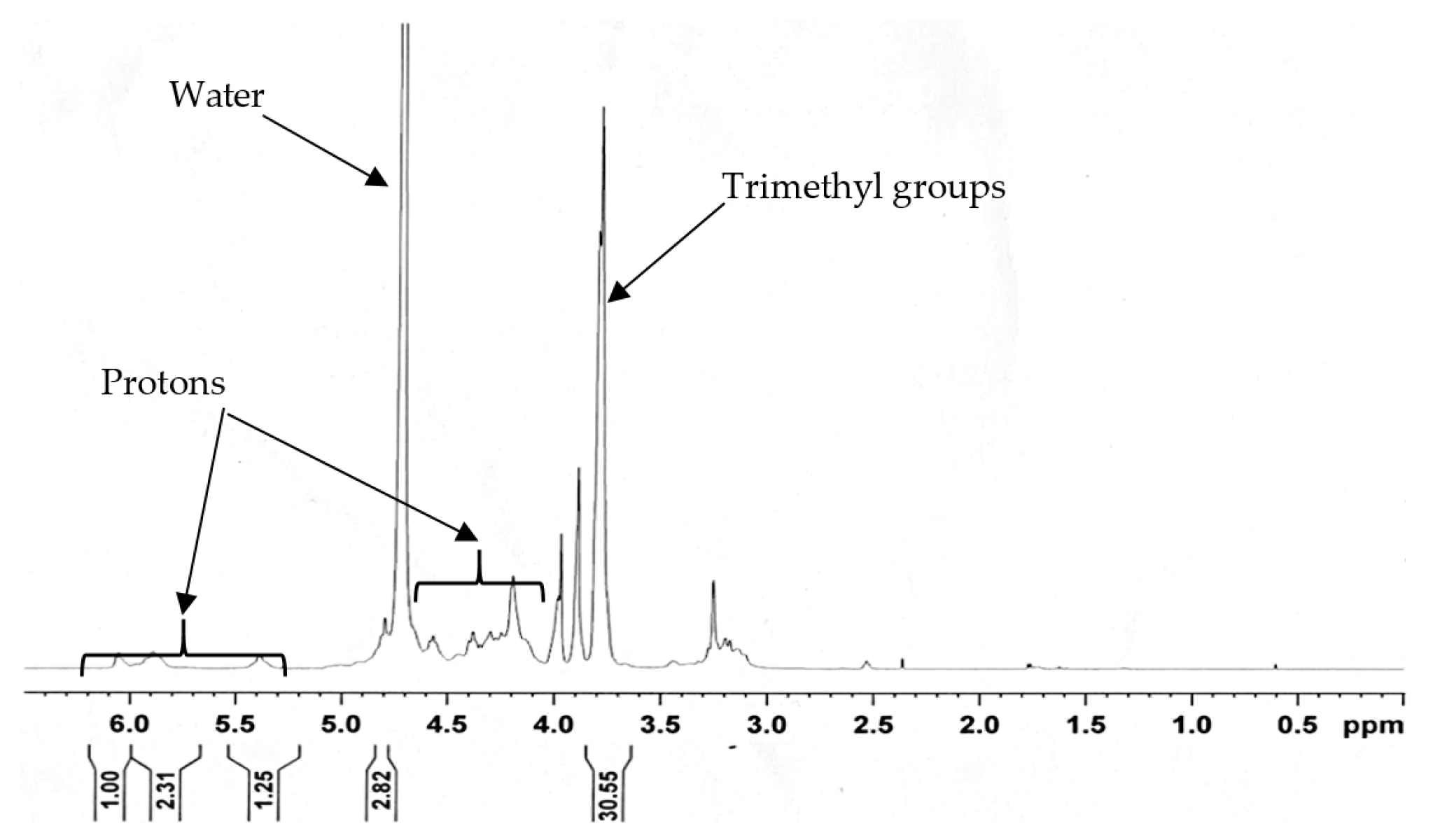
3.2. Characterization of the N-trimethyl Chitosan (TMC)
3.3. Cell Monolayer Integrity Using Lucifer Yellow
3.4. In Vitro Transepithelial Electrical Resistance (TEER) Study
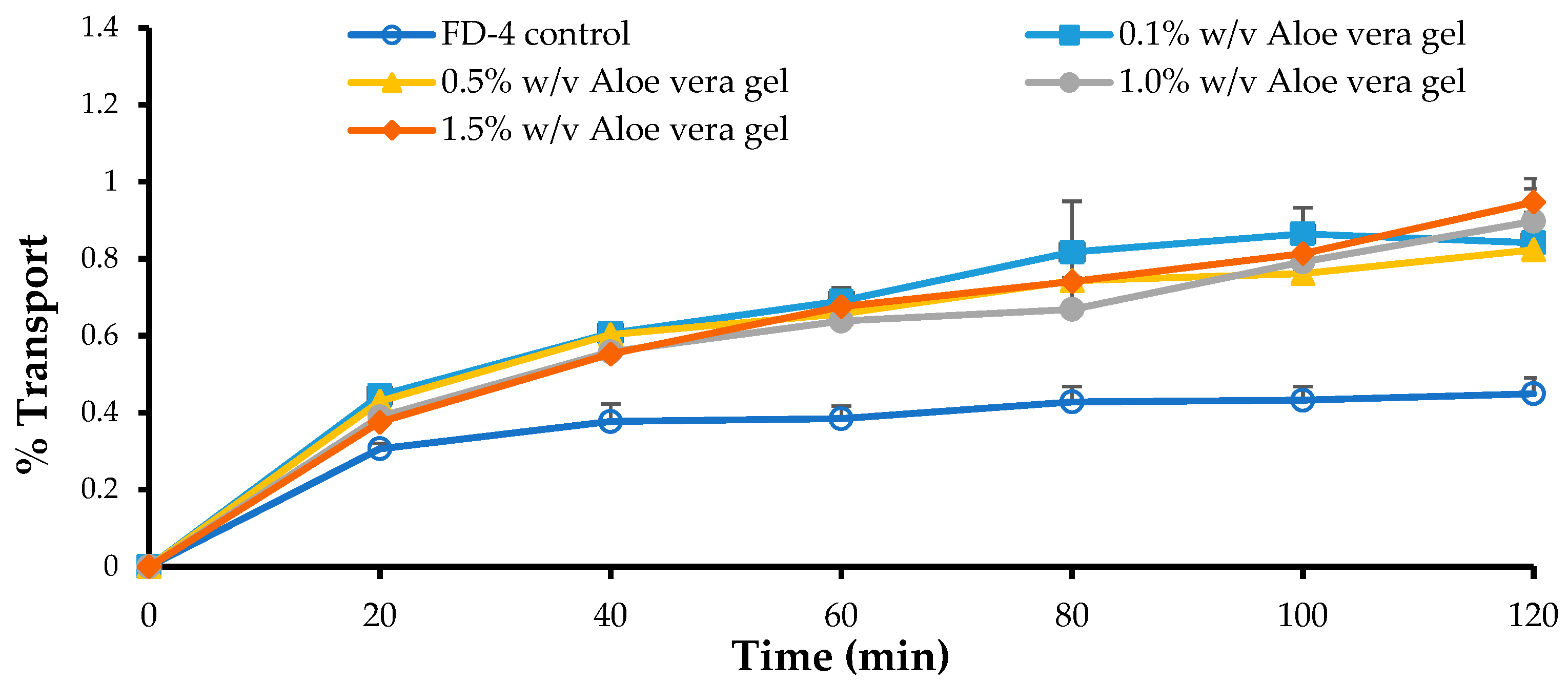
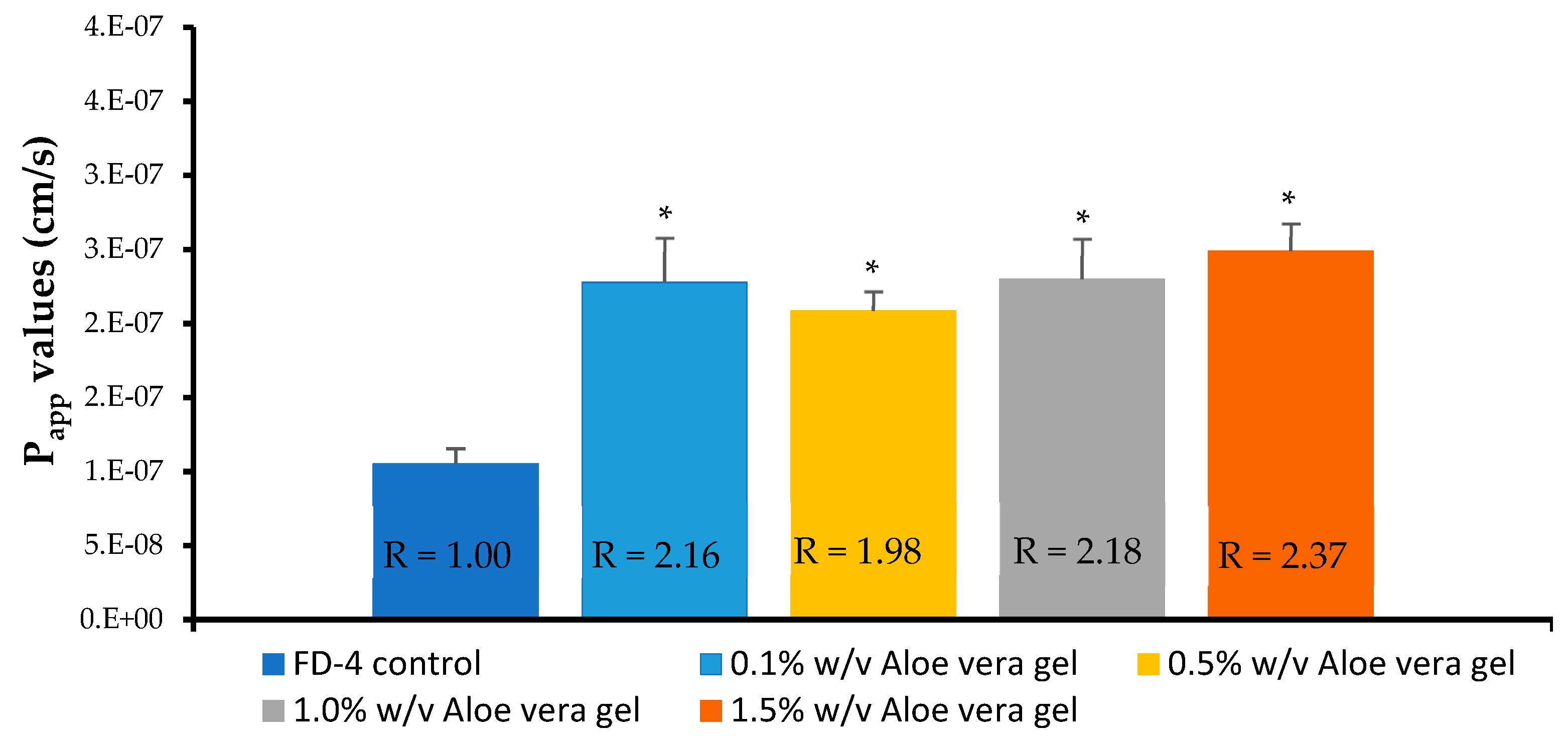
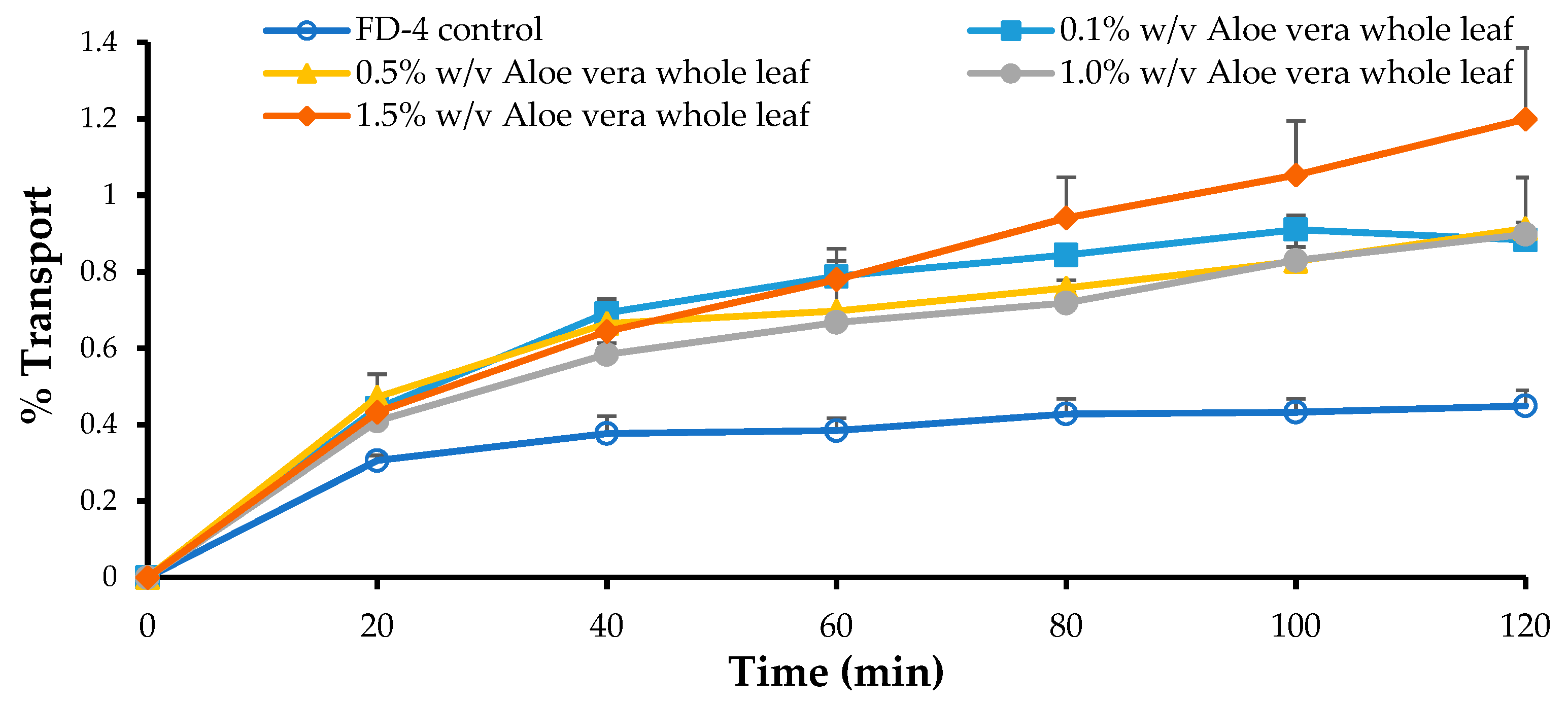
3.5. In Vitro Permeation Studies
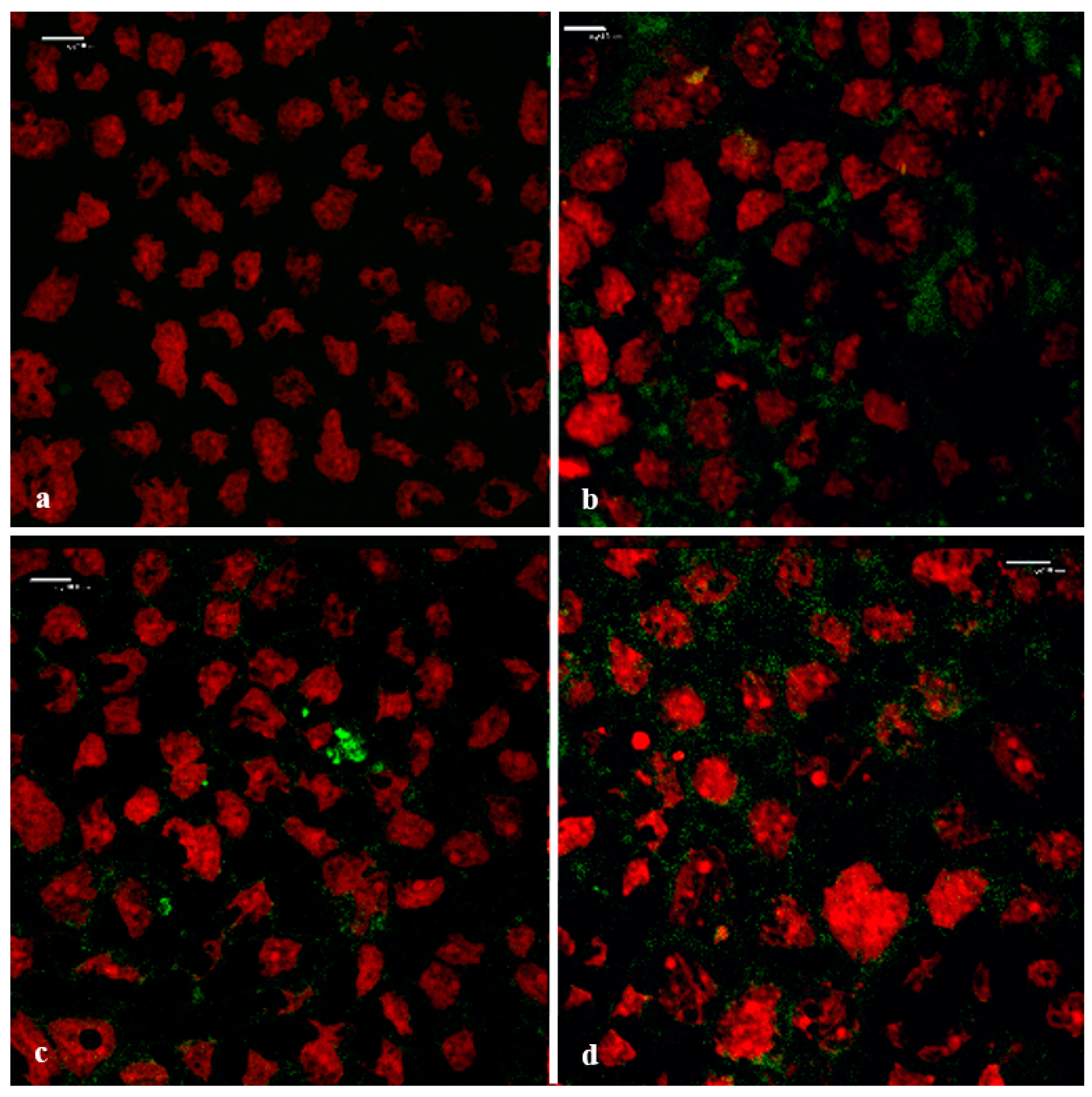
3.6. Confocal Laser Scanning Microscopy (CLSM)
3.6.1. Visualization of the Transport Pathway
3.6.2. Visualization of the F-Actin Filaments in the Cytoskeleton
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Park, K.; Kwon, I.C.; Park, K. Oral protein delivery: Current status and future prospect. React. Funct. Polym. 2011, 71, 280–287. [Google Scholar] [CrossRef]
- Griffin, B.T.; Guo, J.; Presas, E.; Donovan, M.D.; Alonso, M.J.; O’Driscoll, C.M. Pharmacokinetic, pharmacodynamic and biodistribution following oral administration of nanocarriers containing peptide and protein drugs. Adv. Drug Deliv. Rev. 2016, 106, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Muheem, A.; Shakeel, F.; Jahangir, M.A.; Anwar, M.; Mallick, N.; Jain, G.K.; Warsi, M.H.; Ahmad, F.J. A review on the strategies for the oral delivery of proteins and peptides and their clinical perspectives. Saudi Pharm. J. 2016, 24, 413–428. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Navarro, M.; Garcia, J.; Giralt, E.; Teixidó, M. Using peptides to increase transport across the intestinal barrier. Adv. Drug Deliv. Rev. 2016, 106, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Li, B.V.; Uppoor, R.S.; Mehta, M.; Yu, L.X. Bioavailability and bioequivalence. In Developing Solid Oral Dosage Forms: Pharmaceutical Theory & Practice, 2nd ed.; Qiu, Y., Chen, Y., Zhang, G.G.Z., Yu, L., Mantri, R.V., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 381–397. ISBN 978-01-2802-447-8. [Google Scholar]
- Cabrera-Pérez, M.A.; Sanz, M.B.; Sanjuan, V.M.; González-Álvarez, M.; González-Álvarez, I. Importance and applications of cell- and tissue-based in vitro models for drug permeability screening in early stages of drug development. In Concepts and Models for Drug Permeability Studies, 1st ed.; Sarmento, B., Ed.; Woodhead Publishing: Cambridge, UK, 2016; pp. 3–29. ISBN 978-00-8100-094-6. [Google Scholar]
- Rosenthal, R.; Günzel, D.; Finger, C.; Krug, S.M.; Richter, J.F.; Schulzke, J.; Fromm, M.; Amasheh, S. The effect of chitosan on transcellular and paracellular mechanisms in the intestinal epithelial barrier. Biomaterials 2012, 33, 2791–2800. [Google Scholar] [CrossRef] [PubMed]
- Artursson, P.; Palm, K.; Luthman, K. Caco-2 monolayers in experimental and theoretical predictions of drug transport. Adv. Drug Deliv. Rev. 2001, 46, 27–43. [Google Scholar] [CrossRef]
- Lin, Y.; Chen, C.; Liang, H.; Kulkarni, A.R.; Lee, P.; Chen, C.; Sung, H. Novel nanoparticles for oral insulin delivery via the paracellular pathway. Nanotechnology 2007, 18, 1–11. [Google Scholar] [CrossRef]
- Tscheik, C.; Blasig, I.E.; Winkler, L. Trends in drug delivery through tissue barriers containing tight junctions. Tissue Barriers 2013, 1, e24565. [Google Scholar] [CrossRef]
- Ward, P.D.; Tippin, T.K.; Thakker, D.R. Enhancing paracellular permeability by modulating epithelial tight junctions. Pharm. Sci. Technol. Today 2000, 3, 346–358. [Google Scholar] [CrossRef]
- Lemmer, H.J.R.; Hamman, J.H. Paracellular drug absorption enhancement through tight junction modulation. Expert Opin. Drug Deliv. 2013, 10, 103–114. [Google Scholar] [CrossRef]
- Matter, K.; Balda, M.S. Functional analysis of tight junctions. Methods 2003, 30, 228–234. [Google Scholar] [CrossRef]
- Sun, H.; Chow, E.C.Y.; Liu, S.; Du, Y.; Pang, K.S. The Caco-2 cell monolayer: Usefulness and limitations. Expert Opin. Drug Metab. Toxicol. 2008, 4, 395–411. [Google Scholar] [CrossRef] [PubMed]
- Maher, S.; Mrsny, R.J.; Brayden, D.J. Intestinal permeation enhancers for oral peptide delivery. Adv. Drug Deliv. Rev. 2016, 106, 277–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakkireddy, H.R.; Urmannb, M.; Besenius, M.; Werner, U.; Haack, T.; Brun, P.; Alié, J.; Illel, B.; Hortala, L.; Vogel, R.; et al. Oral delivery of diabetes peptides—Comparing standard formulations incorporating functional excipients and nanotechnologies in the translational context. Adv. Drug Deliv. Rev. 2016, 106, 196–222. [Google Scholar] [CrossRef] [PubMed]
- Renukuntla, J.; Vadlapudi, A.D.; Patel, A.; Boddu, S.H.S.; Mitra, A.K. Approaches for enhancing oral bioavailability of peptides and proteins. Int. J. Pharm. 2013, 447, 75–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahato, R.I.; Narang, A.S.; Thoma, L.; Miller, D.D. Emerging trends in oral delivery of peptide and protein drugs. Crit. Rev. Ther. Drug Carrier Syst. 2003, 20, 153–214. [Google Scholar] [CrossRef]
- Isoda, H.; Han, J.; Tominaga, M.; Maekawa, T. Effects of capsaicin on human intestinal cell line Caco-2. Cytotechnology 2001, 36, 155–161. [Google Scholar] [CrossRef]
- Kesarwani, K.; Gupta, R. Bioavailability enhancers of herbal origin: An overview. Asian Pac. J. Trop. Biomed. 2013, 3, 253–266. [Google Scholar] [CrossRef] [Green Version]
- Salama, N.N.; Eddington, N.D.; Fasano, A. Tight junction modulation and its relationship to drug delivery. Adv. Drug Deliv. Rev. 2006, 58, 15–28. [Google Scholar] [CrossRef]
- Tatiraju, D.V.; Bagade, V.B.; Karambelkar, P.J.; Jadhav, V.M.; Kadam, V. Natural bioenhancers: An overview. J. Pharmacogn. Phytochem. 2013, 2, 55–60. [Google Scholar]
- Werle, M.; Bernkop-Schnürch, A. Thiolated chitosans: Useful excipients for oral drug delivery. J. Pharm. Pharmacol. 2008, 60, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Vllasaliu, D.; Casettari, L.; Fowler, R.; Exposito-Harris, R.; Garnett, M.; Illum, L.; Stolnik, S. Absorption-promoting effects of chitosan in airway and intestinal cell lines: A comparative study. Int. J. Pharm. 2012, 430, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, M.D.; Beland, F.A. An evaluation of the biological and toxicological properties of Aloe barbadensis (Miller), Aloe vera. J. Environ. Sci. Health C 2006, 24, 103–154. [Google Scholar] [CrossRef] [PubMed]
- Sahu, P.K.; Giri, D.D.; Singh, R.; Pandey, P.; Gupta, S.; Shrivastava, A.K.; Kumar, A.; Pandey, K.D. Therapeutic and medicinal uses of Aloe vera: A review. Pharmacol. Pharm. 2013, 4, 599–610. [Google Scholar] [CrossRef]
- Eshun, K.; He, Q. Aloe vera: A valuable ingredient for the food, pharmaceutical and cosmetic industries—A review. Crit. Rev. Food Sci. Nutr. 2004, 44, 91–96. [Google Scholar] [CrossRef]
- Hamman, J.H. Composition and applications of Aloe vera leaf gel. Molecules 2008, 13, 1599–1616. [Google Scholar] [CrossRef]
- Sánchez-Machado, D.I.; López-Cervantes, J.; Sendón, R.; Sanches-Silva, A. Aloe vera: Ancient knowledge with new frontiers. Trends Food Sci. Technol. 2017, 61, 94–102. [Google Scholar] [CrossRef]
- Mascolo, N.; Izzo, A.A.; Borrelli, F.; Capasso, R.; Di Carlo, G.; Sautebin, L.; Capasso, F. Healing powers of aloes. In Aloes: The genus Aloe, 1st ed.; Reynolds, T., Ed.; CRC Press: Washington, DC, USA, 2004; pp. 209–238. ISBN 978-04-1530-672-0. [Google Scholar]
- Vinson, J.A.; Al Kharrat, H.; Andreoli, L. Effect of Aloe vera preparations on the human bioavailability of vitamins C and E. Phytomedicine 2005, 12, 760–765. [Google Scholar] [CrossRef]
- Chen, W.; Lu, Z.; Viljoen, A.; Hamman, J. Intestinal drug transport enhancement by Aloe vera. Planta Med. 2009, 75, 587–595. [Google Scholar] [CrossRef]
- Beneke, C.; Viljoen, A.; Hamman, J.H. In vitro drug absorption enhancement effects of Aloe vera and Aloe ferox. Sci. Pharm. 2012, 80, 475–486. [Google Scholar] [CrossRef]
- Beneke, C.; Viljoen, A.; Hamman, J.H. Modulation of drug efflux by aloe materials: An in vitro investigation across rat intestinal tissue. Pharmacogn. Mag. 2013, 9, 44–48. [Google Scholar] [CrossRef]
- Lebitsa, T.; Viljoen, A.; Lu, Z.; Hamman, J. In vitro drug permeation enhancement potential of Aloe gel materials. Curr. Drug Deliv. 2012, 9, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Cole, L.; Heard, C. Skin permeation enhancement potential of Aloe vera and a proposed mechanism of action based upon size exclusion and pull effect. Int. J. Pharm. 2007, 333, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Fox, L.; Gerber, M.; Du Preez, J.L.; Du Plessis, J.; Hamman, J.H. Skin permeation enhancement effects of the gel and whole leaf materials of Aloe vera, Aloe marlothii and Aloe ferox. J. Pharm. Pharmacol. 2014, 67, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Ojewole, E.; Mackraj, I.; Akhundov, K.; Hamman, J.; Viljoen, A.; Olivier, E.; Wesley-Smith, J.; Govender, T. Investigating the effect of Aloe vera gel on the buccal permeability of didanosine. Planta Med. 2012, 78, 354–361. [Google Scholar] [CrossRef]
- Djuv, A.; Nilsen, O.G. Caco-2 cell methodology and inhibition of the P-glycoprotein transport of digoxin by Aloe vera juice. Phytother. Res. 2008, 22, 1623–1628. [Google Scholar] [CrossRef]
- Jiao, P.; Jia, Q.; Randel, G.; Diehl, B.; Weaver, S.; Milligan, G. Quantitative 1H-NMR Spectrometry Method for Quality Control of Aloe vera Products. J. AOAC Int. 2010, 93, 842–848. [Google Scholar]
- Sieval, A.B.; Thanou, M.; Kotzé, A.F.; Verhoef, J.C.; Brussee, J.; Junginger, H.E. Preparation and NMR characterization of highly substituted N-trimethyl chitosan chloride. Carbohydr. Polym. 1998, 36, 157–165. [Google Scholar] [CrossRef]
- Hamman, J.H.; Stander, M.; Kotzé, A.F. Effect of the degree of quaternisation of N-trimethyl chitosan chloride of absorption enhancement: In vivo evaluation in rat nasal epithelia. Int. J. Pharm. 2002, 232, 235–242. [Google Scholar] [CrossRef]
- Calatayud, M.; Devesa, V.; Montoro, R.; Vélez, D. In vitro study of intestinal transport of arsenite, monomethylarsonous acid, and dimethylarsinous acid by Caco-2 cell line. Toxicol. Lett. 2011, 204, 127–133. [Google Scholar] [CrossRef]
- Bhushani, J.A.; Karthik, P.; Anandharamakrishnan, C. Nanoemulsion based delivery system for improved bioaccessibility and Caco-2 cell monolayer permeability of green tea extract. Food Hydrocoll. 2016, 56, 372–382. [Google Scholar] [CrossRef]
- Alqahtani, S.; Mohamed, L.A.; Kaddoumi, A. Experimental models for predicting drug absorption and metabolism. Expert Opin. Drug Metab. Toxicol. 2013, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, T.; Malan, M.M.; Lemmer, H.J.R.; Gouws, C.; Aucamp, M.E.; Breytenbach, W.J.; Hamman, J.H. Combining chemical permeation enhancers for synergistic effects. Eur. J. Drug Metab. Pharmacokinet. 2016, 41, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Pick, D.; Degen, C.; Leiterer, M.; Jahreis, G.; Einax, J.W. Transport of selenium species in Caco-2 cells: Analytical approach employing the Ussing chamber technique and HPLC-ICP-MS. Microchem. J. 2013, 110, 8–14. [Google Scholar] [CrossRef]
- Wahlang, B.; Pawar, Y.B.; Bansal, A.K. Identification of permeability-related hurdles in oral delivery of curcumin using the Caco-2 cell model. Eur. J. Pharm. Biopharm. 2011, 77, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, P.; Picchini, U.; Beck, B.; van Gelder, J.; Delbar, N.; DaGaetano, A. A general approach to the apparent permeability index. J. Pharmacokinet. Pharmacodyn. 2008, 35, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.H.; Frank, D.; Costantino, H.R. Discovery of tight junction modulators: Significance for drug development and delivery. Drug Discov. Today 2008, 13, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Kotzé, A.F.; Leußen, H.L.; De Leeuw, B.J.; De Boer, B.G.; Verhoef, J.C.; Junginger, H.E. Comparison of the effect of different chitosan salts and N-trimethyl chitosan chloride on the permeability of intestinal epithelial cells (Caco-2). J. Control Release 1998, 51, 35–46. [Google Scholar] [CrossRef]
- Abcam. Immunocytochemistry and Immunofluorescence Protocol: Procedure for Staining of Cell Cultures Using Immunofluorescence. Available online: http://www.abcam.com/protocols/immunocytochemistry-immunofluorescence-protocol (accessed on 9 November 2017).
- Wu, S.; Don, T.; Lin, C.; Mi, F. Delivery of berberine using chitosan/fucoidan-taurine conjugate nanoparticles for treatment of defective intestinal epithelial tight junction barrier. Mar. Drugs 2014, 12, 5677–5697. [Google Scholar] [CrossRef]
- Abcam. Protocol Booklet: CytoPainter Phalloidin-iFlour 488 Reagent: Instructions for Use for Staining F-Actin in Adherent or Suspension Cells. Available online: http://www.abcam.com/ps/products/176/ab176753/documents/ab176753%20CytoPainter%20Phalloidin-iFluor%20488%20Reagent%20protocol%20v3%20(website).pdf (accessed on 9 November 2017).
- Dorkoosh, F.A.; Broekhuizen, C.A.N.; Borchard, G.; Rafiee-Tehrani, M.; Verhoef, J.C.; Junginger, H.E. Transport of octreotide and evaluation of mechanism of action of opening the paracellular tight junction using superporous hydrogel polymers in Caco-2 cell monolayers. J. Pharm. Sci. 2004, 93, 743–752. [Google Scholar] [CrossRef]
- Hsu, L.; Ho, Y.; Chuang, E.; Chen, C.; Juang, J.; Su, F.; Hwang, S.; Sung, H. Effects of pH on molecular mechanisms of chitosan-integrin interactions and resulting tight-junction disruptions. Biomaterials 2013, 34, 784–793. [Google Scholar] [CrossRef] [PubMed]
- Abcam. Product Datasheet: Fluoroshield Mounting Medium with Propidium Iodide ab104129. Available online: http://www.abcam.com/fluoroshield-mounting-medium-with-propidium-iodide-20ml-ab104129.html (accessed on 9 November 2017).
- Kotzé, A.F.; Leußen, H.L.; De Leeuw, B.J.; De Boer, B.G.; Verhoef, J.C.; Junginger, H.E. N-trimethyl chitosan chloride as a potential absorption enhancer across mucosal surfaces: In vitro evaluation in intestinal epithelial cells (Caco-2). Pharm. Res. 1997, 14, 1197–1202. [Google Scholar] [CrossRef] [PubMed]
- Thanou, M.; Verhoef, J.C.; Junginger, H.E. Oral drug absorption enhancement by chitosan and its derivatives. Adv. Drug Deliv. Rev. 2001, 52, 117–126. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
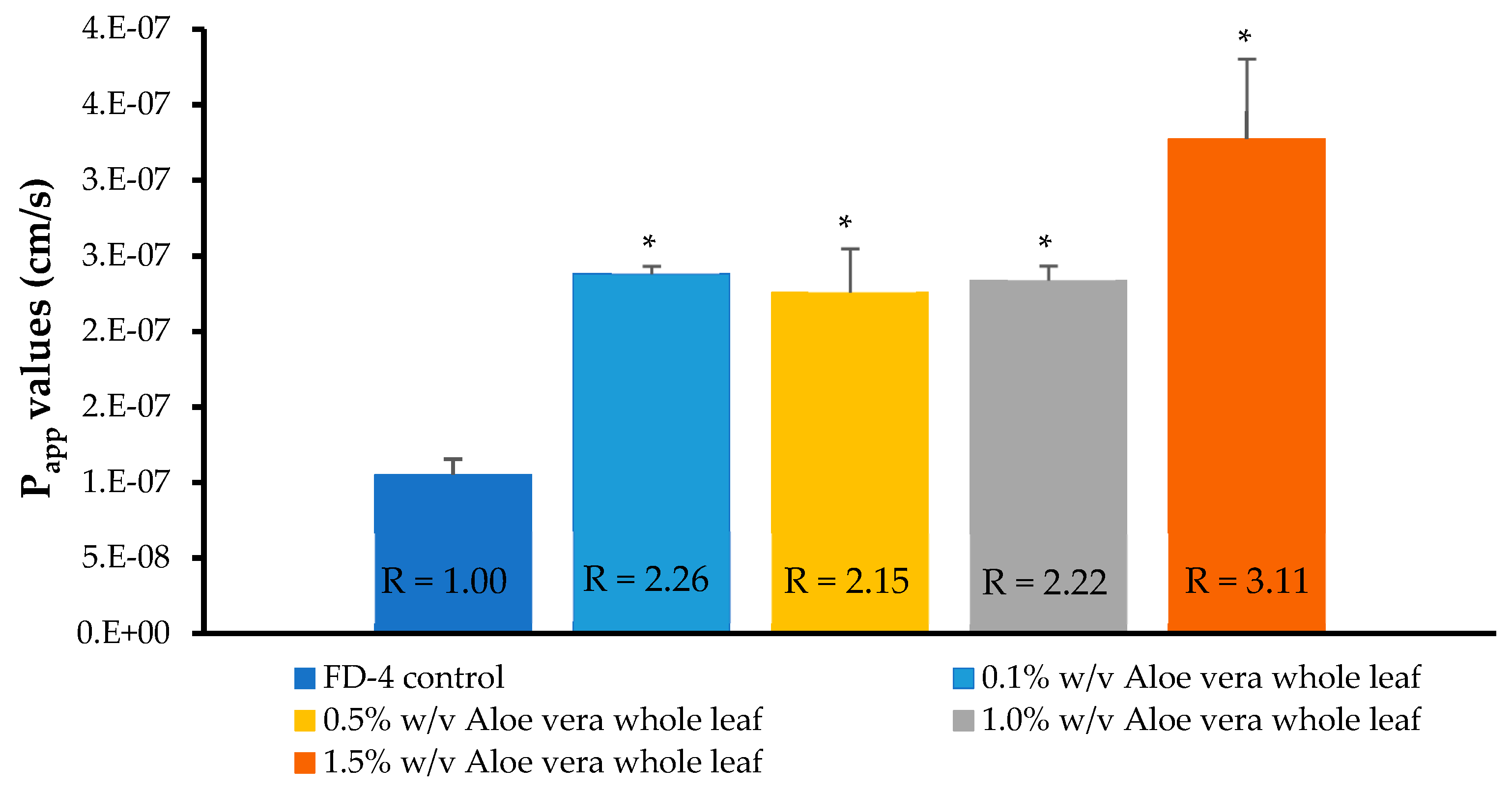{kind=link}
{kind=link}
{kind=link}
| Type of Transwell® Plate | TEER Value Measured (Ω) | TEER Value Normalized for Surface Area (Ω·cm2) |
|---|---|---|
| Transwell® 6-well plates (surface area = 4.67 cm2) [45] | 150 | 700.5 |
| Transwell® 24-well plates (surface area = 0.33 cm2) [46] | 750 | 247.5 |
| Snapwell® 6-well plates (surface area = 1.12 cm2) [47] | 179 | 200.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haasbroek, A.; Willers, C.; Glyn, M.; du Plessis, L.; Hamman, J. Intestinal Drug Absorption Enhancement by Aloe vera Gel and Whole Leaf Extract: In Vitro Investigations into the Mechanisms of Action. Pharmaceutics 2019, 11, 36. https://doi.org/10.3390/pharmaceutics11010036
Haasbroek A, Willers C, Glyn M, du Plessis L, Hamman J. Intestinal Drug Absorption Enhancement by Aloe vera Gel and Whole Leaf Extract: In Vitro Investigations into the Mechanisms of Action. Pharmaceutics. 2019; 11(1):36. https://doi.org/10.3390/pharmaceutics11010036
Chicago/Turabian StyleHaasbroek, Anja, Clarissa Willers, Matthew Glyn, Lissinda du Plessis, and Josias Hamman. 2019. "Intestinal Drug Absorption Enhancement by Aloe vera Gel and Whole Leaf Extract: In Vitro Investigations into the Mechanisms of Action" Pharmaceutics 11, no. 1: 36. https://doi.org/10.3390/pharmaceutics11010036
APA StyleHaasbroek, A., Willers, C., Glyn, M., du Plessis, L., & Hamman, J. (2019). Intestinal Drug Absorption Enhancement by Aloe vera Gel and Whole Leaf Extract: In Vitro Investigations into the Mechanisms of Action. Pharmaceutics, 11(1), 36. https://doi.org/10.3390/pharmaceutics11010036