Anionic Complex with Efficient Expression and Good Safety Profile for mRNA Delivery

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Preparation of Complexes
2.3. Physicochemical Properties of the Complexes
2.4. In Vitro Protein Expression
2.5. Cytotoxicity of the Complexes
2.6. Interaction of the Complexes with Erythrocytes
2.7. Animals
2.8. In Vivo Protein Expression
2.9. Statistical Analysis
3. Results
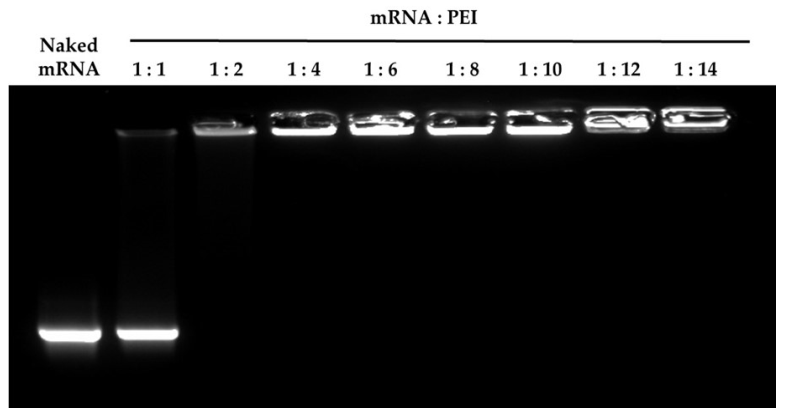
3.1. Physicochemical Properties of the mRNA/PEI Complexes
3.2. In Vitro Protein Expression of the mRNA/PEI Complexes
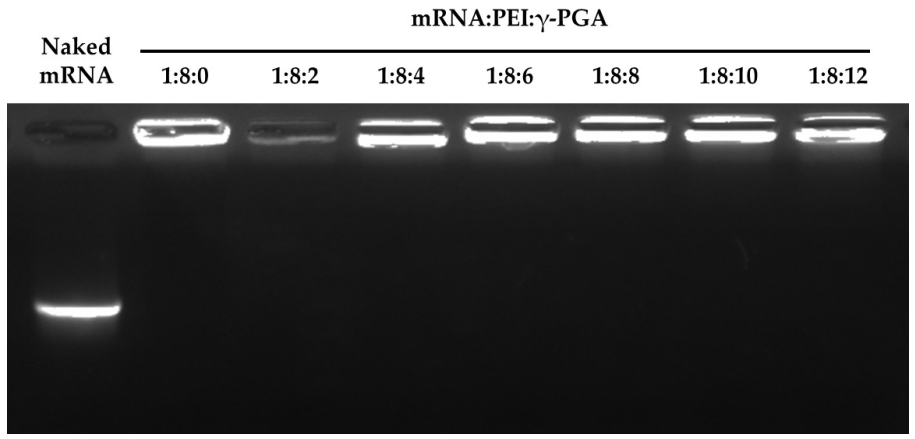
3.3. Physicochemical Properties of the mRNA/PEI8/γ-PGA Complexes
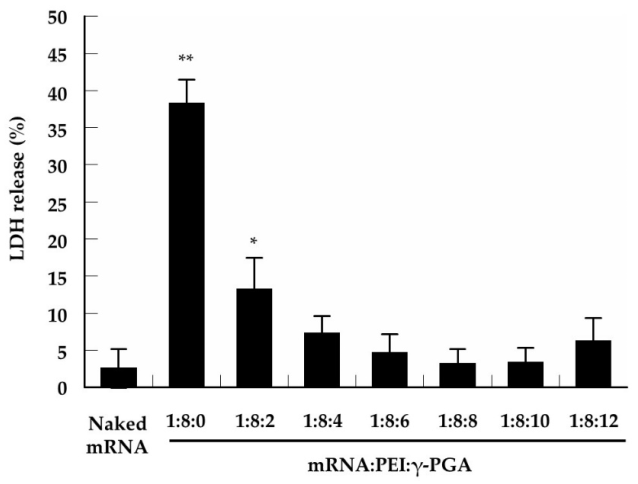
3.4. Cytotoxicity of the Complexes
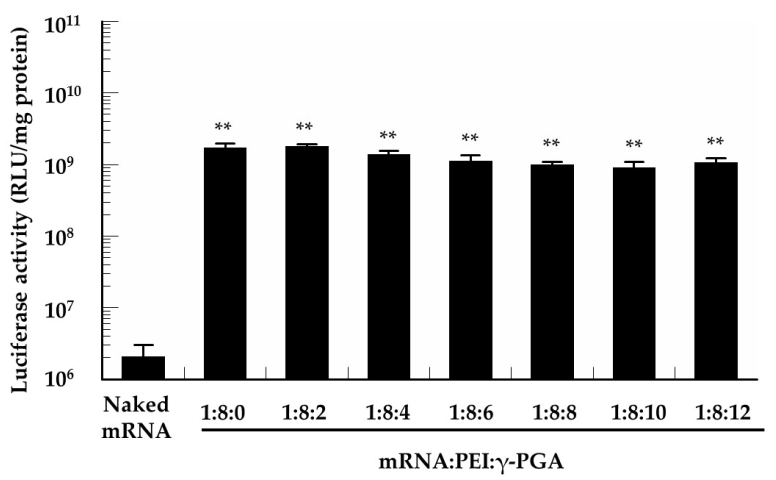
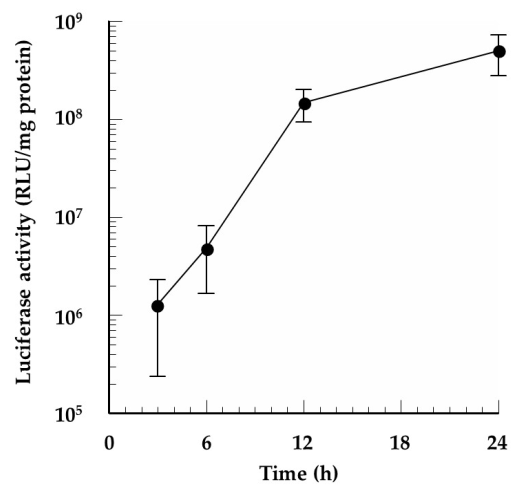
3.5. In Vitro Protein Expression of the mRNA/PEI8/γ-PGA Complexes
3.6. RNase Tolerance of the mRNA/PEI8 Complex and the mRNA/PEI8/γ-PGA12 Complex
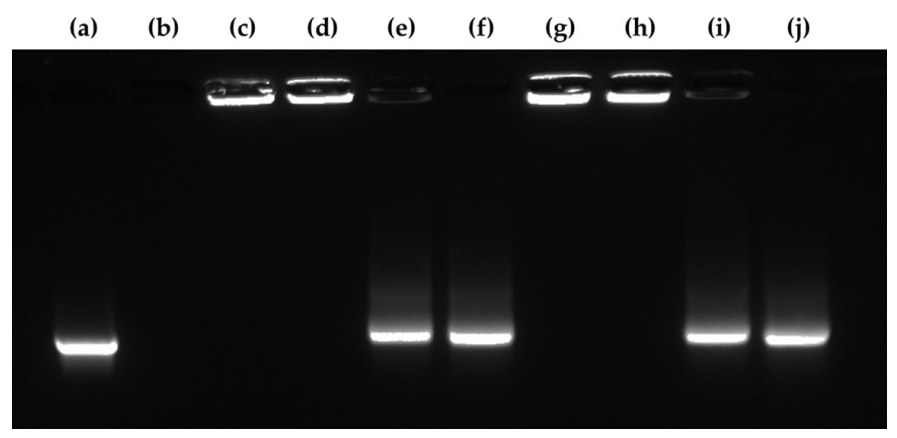
3.7. Agglutination Study of the mRNA/PEI8 Complex and mRNA/PEI8/γ-PGA12 Complex
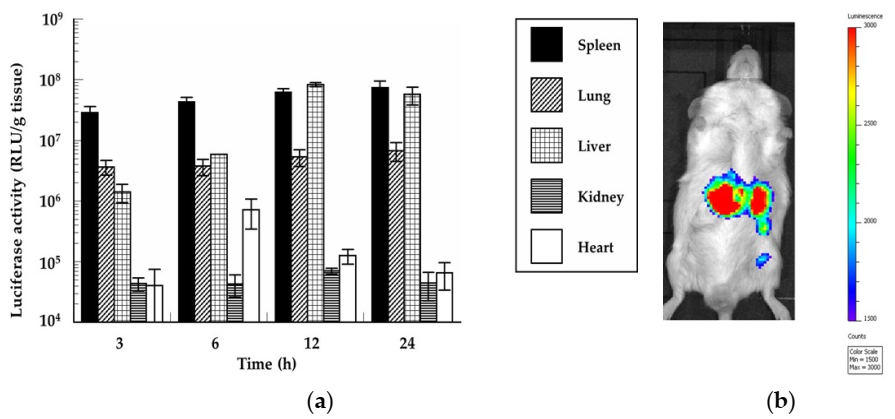
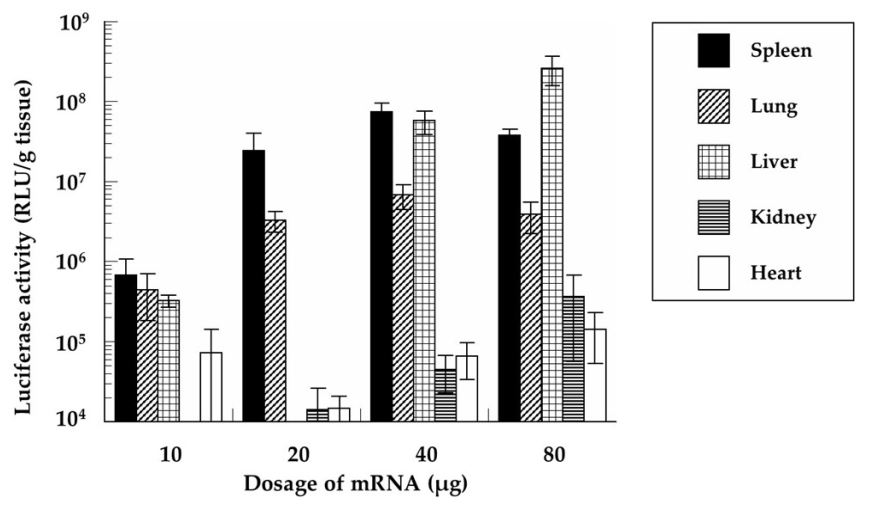
3.8. In Vivo Protein Expression of the mRNA/PEI8/γ-PGA12 Complex
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guan, S.; Rosenecker, J. Nanotechnologies in delivery of mRNA therapeutics using nonviral vector-based delivery systems. Gene Ther. 2017, 24, 133–143. [Google Scholar] [CrossRef]
- Pardi, N.; Tuyishime, S.; Muramatsu, H.; Kariko, K.; Mui, B.L.; Tam, Y.K.; Madden, T.D.; Hope, M.J.; Weissman, D. Expression kinetics of nucleoside-modified mRNA delivered in lipid nanoparticles to mice by various routes. J. Control. Release 2015, 217, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Yen, A.; Cheng, Y.; Sylvestre, M.; Gustafson, H.H.; Puri, S.; Pun, S.H. Serum Nuclease Susceptibility of mRNA Cargo in Condensed Polyplexes. Mol. Pharm. 2018, 15, 2268–2276. [Google Scholar] [CrossRef]
- Zhao, M.; Li, M.; Zhang, Z.; Gong, T.; Sun, X.; Zhao, M.; Li, M.; Zhang, Z.; Gong, T.; Sun, X. Induction of HIV-1 gag specific immune responses by cationic micelles mediated delivery of gag mRNA. Drug Deliv. 2016, 23, 2596–2607. [Google Scholar] [CrossRef]
- McKinlay, C.J.; Benner, N.L.; Haabeth, O.A.; Waymouth, R.M.; Wender, P.A. Enhanced mRNA delivery into lymphocytes enabled by lipid-varied libraries of charge-altering releasable transporters. Proc. Natl. Acad. Sci. USA 2018, 115, E5859–E5866. [Google Scholar] [CrossRef] [Green Version]
- Uchida, S.; Kataoka, K.; Itaka, K. Screening of mRNA Chemical Modification to Maximize Protein Expression with Reduced Immunogenicity. Pharmaceutics 2015, 7, 137–151. [Google Scholar] [CrossRef] [Green Version]
- Holtkamp, S.; Kreiter, S.; Selmi, A.; Simon, P.; Koslowski, M.; Huber, C.; Türeci, Ö.; Sahin, U. Modification of antigen-encoding RNA increases stability, translational efficacy, and T-cell stimulatory capacity of dendritic cells. Blood 2006, 108, 4009–4017. [Google Scholar] [CrossRef]
- Debus, H.; Baumhof, P.; Probst, J.; Kissel, T. Delivery of messenger RNA using poly(ethylene imine)-poly(ethylene glycol)-copolymer blends for polyplex formation: Biophysical characterization and in vitro transfection properties. J. Control. Release 2010, 148, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Tagalakis, A.D.; Kenny, G.D.; Bienemann, A.S.; McCarthy, D.; Munye, M.M.; Taylor, H.; Wyatt, M.J.; Lythgoe, M.F.; White, E.A.; Hart, S.L. PEGylation improves the receptor-mediated transfection efficiency of peptide-targeted, self-assembling, anionic nanocomplexes. J. Control. Release 2014, 174, 177–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trubetskoy, V.S.; Wong, S.C.; Subbotin, V.; Budker, V.G.; Loomis, A.; Hagstrom, J.E.; Wolff, J.A. Recharging cationic DNA complexes with highly charge polyanions for in vitro and in vivo gene delivery. Gene Ther. 2003, 10, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Park, K.; Kim, J.; Kim, K.S.; Oh, E.J.; Kang, H.; Han, S.E.; Oh, Y.K.; Park, T.G.; Hahn, S.K. Hyaluronic Acid-Polyethyleneimine Conjugate for Target Specific Intracellular Delivery of siRNA. Biopolymers 2008, 89, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.F.; Hsu, H.K.; Lin, C.C.; Cheng, Y.M.; Hsu, K.H. Novel PEI/Poly-γ-Gutamic Acid Nanoparticles for High Efficient siRNA and Plasmid DNA Co-Delivery. Molecules 2017, 22, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurosaki, T.; Kitahara, T.; Fumoto, S.; Nishida, K.; Nakamura, J.; Niidome, T.; Kodama, Y.; Nakagawa, H.; To, H.; Sasaki, H. Ternary complexes of pDNA, polyethylenimine, and γ-polyglutamic acid for gene delivery systems. Biomaterials 2009, 30, 2846–2853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurosaki, T.; Kodama, Y.; Muro, T.; Higuchi, N.; Nakamura, T.; Kitahara, T.; Miyakoda, M.; Yui, K.; Sasaki, H. Secure Splenic Delivery of Plasmid DNA and Its Application to DNA Vaccine. Biol. Pharm. Bull. 2013, 36, 1800–1806. [Google Scholar] [CrossRef] [Green Version]
- Leong, S.P.L.; Cady, B.; Jablons, D.M.; Garcia-Aguilar, J.; Reintgen, D.; Jakub, J.; Pendas, S.; Duhaime, L.; Cassell, R.; Gardner, M.; et al. Clinical patterns of metastasis. Cancer Metastasis Rev. 2006, 25, 221–232. [Google Scholar] [CrossRef]
- Mocellin, S.; Hoon, D.; Ambrosi, A.; Nitti, D.; Rossi, C.R. The prognostic value of circulating tumor cells in patients with melanoma: A systematic review and meta-analysis. Clin. Cancer Res. 2006, 12, 4605–4613. [Google Scholar] [CrossRef] [Green Version]
- Francken, A.B.; Accortt, N.A.; Shaw, H.M.; Wiener, M.; Soong, S.J.; Hoekstra, H.J.; Thompson, J.F. Prognosis and determinants of outcome following locoregional or distant recurrence in patients with cutaneous melanoma. Ann. Surg. Oncol. 2008, 15, 1476–1484. [Google Scholar] [CrossRef]
- Shuaibu, M.N.; Cherif, M.S.; Kurosaki, T.; Helegbe, G.K.; Kikuchi, M.; Yanagi, T.; Sasaki, H.; Hirayama, K. Effect of nanoparticle coating on the immunogenicity of plasmid DNA vaccine encoding P. yoelii MSP-1 C-terminal. Vaccine 2011, 29, 3239–3247. [Google Scholar] [CrossRef]
- Kodama, Y.; Nakamura, T.; Kurosaki, T.; Egashira, K.; Mine, T.; Nakagawa, H.; Muro, T.; Kitahara, T.; Higuchi, N.; Sasaki, H. Biodegradable nanoparticles composed of dendrigraft poly-l-lysine for gene delivery. Eur. J. Pharm. Biopharm. 2014, 87, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Wang, H.; Pang, G.; He, T.; Yu, P.; Cheng, G.; Zhang, Y.; Chang, J. A Logic AND-Gated Sonogene Nanosystem for Precisely Regulating the Apoptosis of Tumor Cells. ACS Appl. Mater. Interfaces 2020, 12, 56692–56700. [Google Scholar] [CrossRef]
- Kang, J.H.; Toita, R.; Niidome, T.; Katayama, Y. Effective delivery of DNA into tumor cells and tissues by electroporation of polymer-DNA complex. Cancer Lett. 2008, 265, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.C.; Lee, M.; Bae, Y.H. Polymeric gene carriers. Crit. Rev. Eukaryot. Gene Expr. 2005, 15, 317–342. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.Y.; Pelet, J.M.; Putnam, D. Polymer systems for gene delivery-Past, present, and future. Prog. Polym. Sci. 2007, 32, 799–837. [Google Scholar] [CrossRef]
- Xiong, M.P.; Laird Forrest, M.; Ton, G.; Zhao, A.; Davies, N.M.; Kwon, G.S. Poly(aspartate-g-PEI800), a polyethylenimine analogue of low toxicity and high transfection efficiency for gene delivery. Biomaterials 2007, 28, 4889–4900. [Google Scholar] [CrossRef] [PubMed]
- Boussif, O.; LezoualC’H, F.; Zanta, M.A.; Mergny, M.D.; Scherman, D.; Demeneix, B.; Behr, J.P. A versatile vector for gene and oligonucleotide transfer into cells in culture and in vivo: Polyethylenimine. Proc. Natl. Acad. Sci. USA 1995, 92, 7297–7301. [Google Scholar] [CrossRef] [Green Version]
- Zuber, G.; Dauty, E.; Nothisen, M.; Belguise, P.; Behr, J.P. Towards synthetic viruses. Adv. Drug Deliv. Rev. 2001, 52, 245–253. [Google Scholar] [CrossRef]
- Pandey, A.P.; Sawant, K.K. Polyethylenimine: A versatile, multifunctional non-viral vector for nucleic acid delivery. Mater. Sci. Eng. C 2016, 68, 904–918. [Google Scholar] [CrossRef]
- Kopatz, I.; Remy, J.S.; Behr, J.P. A model for non-viral gene delivery: Through syndecan adhesion molecules and powered by actin. J. Gene Med. 2004, 6, 769–776. [Google Scholar] [CrossRef] [Green Version]
- Jere, D.; Jiang, H.L.; Arote, R.; Kim, Y.K.; Choi, Y.J.; Cho, M.H.; Akaike, T.; Cho, C.S. Degradable polyethylenimines as DNA and small interfering RNA carriers. Expert Opin. Drug Deliv. 2009, 6, 827–834. [Google Scholar] [CrossRef]
- Moghimi, S.M.; Symonds, P.; Murray, J.C.; Hunter, A.C.; Debska, G.; Szewczyk, A. A Two-Stage Poly(ethylenimine)-Mediated Cytotoxicity: Implications for Gene Transfer/Therapy. Mol. Ther. 2005, 11, 990–995. [Google Scholar] [CrossRef]
- Martin, S.J.; Finucane, D.M.; Amarante-Mendes, G.P.; O’Brien, G.A.; Green, D.R. Phosphatidylserine externalization during CD95-induced apoptosis of cells and cytoplasts requires ICE/CED-3 protease activity. J. Biol. Chem. 1996, 271, 28753–28756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waring, P.; Lambert, D.; Sjaarda, A.; Hurne, A.; Beaver, J. Increased cell surface exposure of phosphatidylserine on propidium iodide negative thymocytes undergoing death by necrosis. Cell Death Differ. 1999, 6, 624–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonneau, M.J.; Poulin, R. Spermine oxidation leads to necrosis with plasma membrane phosphatidylserine redistribution in mouse leukemia cells. Exp. Cell Res. 2000, 259, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.A.; Poniris, M.H.; Wilson, M.R. pDMAEMA is internalised by endocytosis but does not physically disrupt endosomes. J. Control. Release 2004, 96, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Goula, D.; Benoist, C.; Mantero, S.; Merlo, G.; Levi, G.; Demeneix, B.A. Polyethylenimine-based intravenous delivery of transgenes to mouse lung. Gene Ther. 1998, 5, 1291–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mRNA:PEI | Particle Size (nm) | ζ-Potential (mV) |
|---|---|---|
| 1:1 | 39.4 ± 17.1 | −8.9 ± 0.6 |
| 1:2 | Aggregated | −0.2 ± 0.2 |
| 1:4 | 84.9 ± 10.6 | 36.1 ± 0.5 |
| 1:6 | 40.7 ± 4.2 | 42.9 ± 0.3 |
| 1:8 | 46.3 ± 6.4 | 47.4 ± 1.4 |
| 1:10 | 38.9 ± 10.1 | 51.2 ± 1.4 |
| 1:12 | 42.9 ± 21.4 | 54.4 ± 0.1 |
| mRNA:PEI:γ-PGA | Particle Size (nm) | ζ-Potential (mV) |
|---|---|---|
| 1:8:0 | 46.3 ± 6.4 | 47.4 ± 1.4 |
| 1:8:2 | 45.1 ± 2.6 | 36.2 ± 0.3 |
| 1:8:4 | Aggregated | 17.0 ± 0.7 |
| 1:8:6 | 80.4 ± 8.0 | −10.0 ± 0.1 |
| 1:8:8 | 44.7 ± 7.9 | −27.2 ± 3.0 |
| 1:8:10 | 46.5 ± 12.4 | −31 ± 0.3 |
| 1:8:12 | 52.0 ± 7.3 | −35.5 ± 1.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamada, E.; Kurosaki, T.; Hashizume, J.; Harasawa, H.; Nakagawa, H.; Nakamura, T.; Kodama, Y.; Sasaki, H. Anionic Complex with Efficient Expression and Good Safety Profile for mRNA Delivery. Pharmaceutics 2021, 13, 126. https://doi.org/10.3390/pharmaceutics13010126
Hamada E, Kurosaki T, Hashizume J, Harasawa H, Nakagawa H, Nakamura T, Kodama Y, Sasaki H. Anionic Complex with Efficient Expression and Good Safety Profile for mRNA Delivery. Pharmaceutics. 2021; 13(1):126. https://doi.org/10.3390/pharmaceutics13010126
Chicago/Turabian StyleHamada, Eri, Tomoaki Kurosaki, Junya Hashizume, Hitomi Harasawa, Hiroo Nakagawa, Tadahiro Nakamura, Yukinobu Kodama, and Hitoshi Sasaki. 2021. "Anionic Complex with Efficient Expression and Good Safety Profile for mRNA Delivery" Pharmaceutics 13, no. 1: 126. https://doi.org/10.3390/pharmaceutics13010126
APA StyleHamada, E., Kurosaki, T., Hashizume, J., Harasawa, H., Nakagawa, H., Nakamura, T., Kodama, Y., & Sasaki, H. (2021). Anionic Complex with Efficient Expression and Good Safety Profile for mRNA Delivery. Pharmaceutics, 13(1), 126. https://doi.org/10.3390/pharmaceutics13010126