
Effects of Polyphenols on P-Glycoprotein (ABCB1) Activity

 ,
,  ,
,  , ,
, , 
 and
and 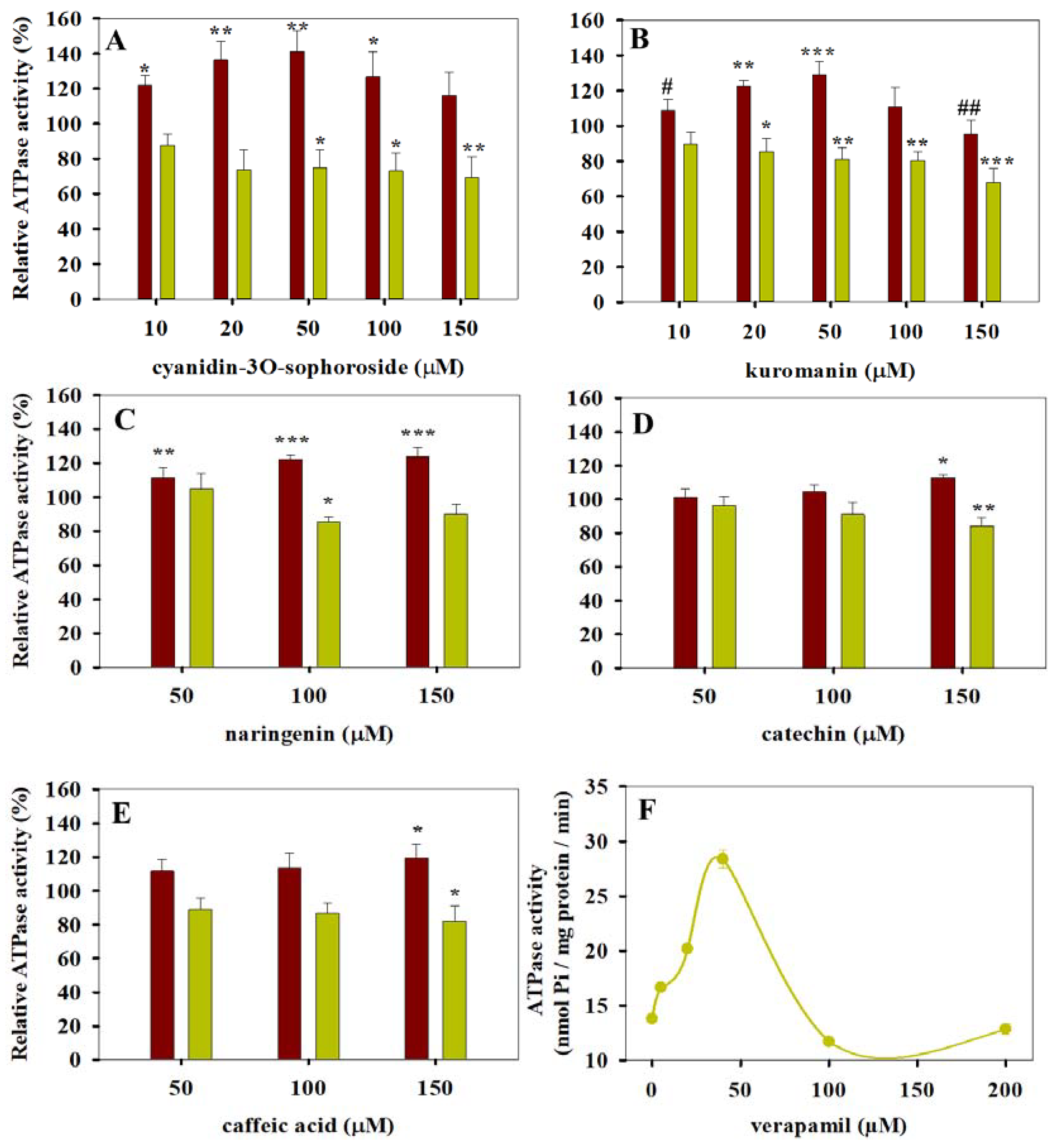{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Lines
2.3. Membrane Preparations for ATPase Activity Measurements
2.4. ATPase Activity Measurements
2.5. Calcein and Daunorubicin Accumulation Tests
2.6. UIC2 Reactivity Test
2.7. Flow Cytometry
2.8. Measurement of Membrane Fluidity
2.9. Statistical Analysis
3. Results
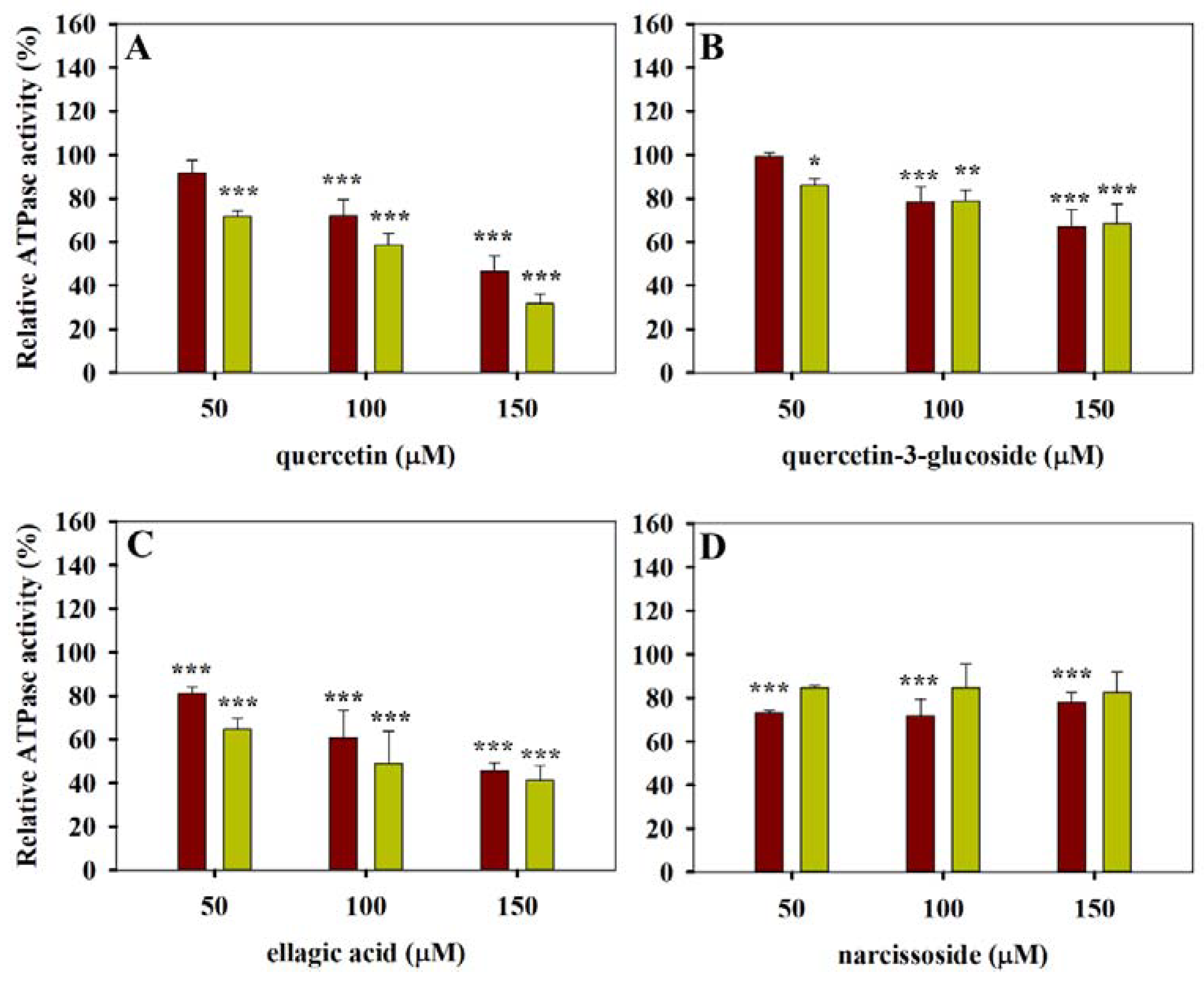
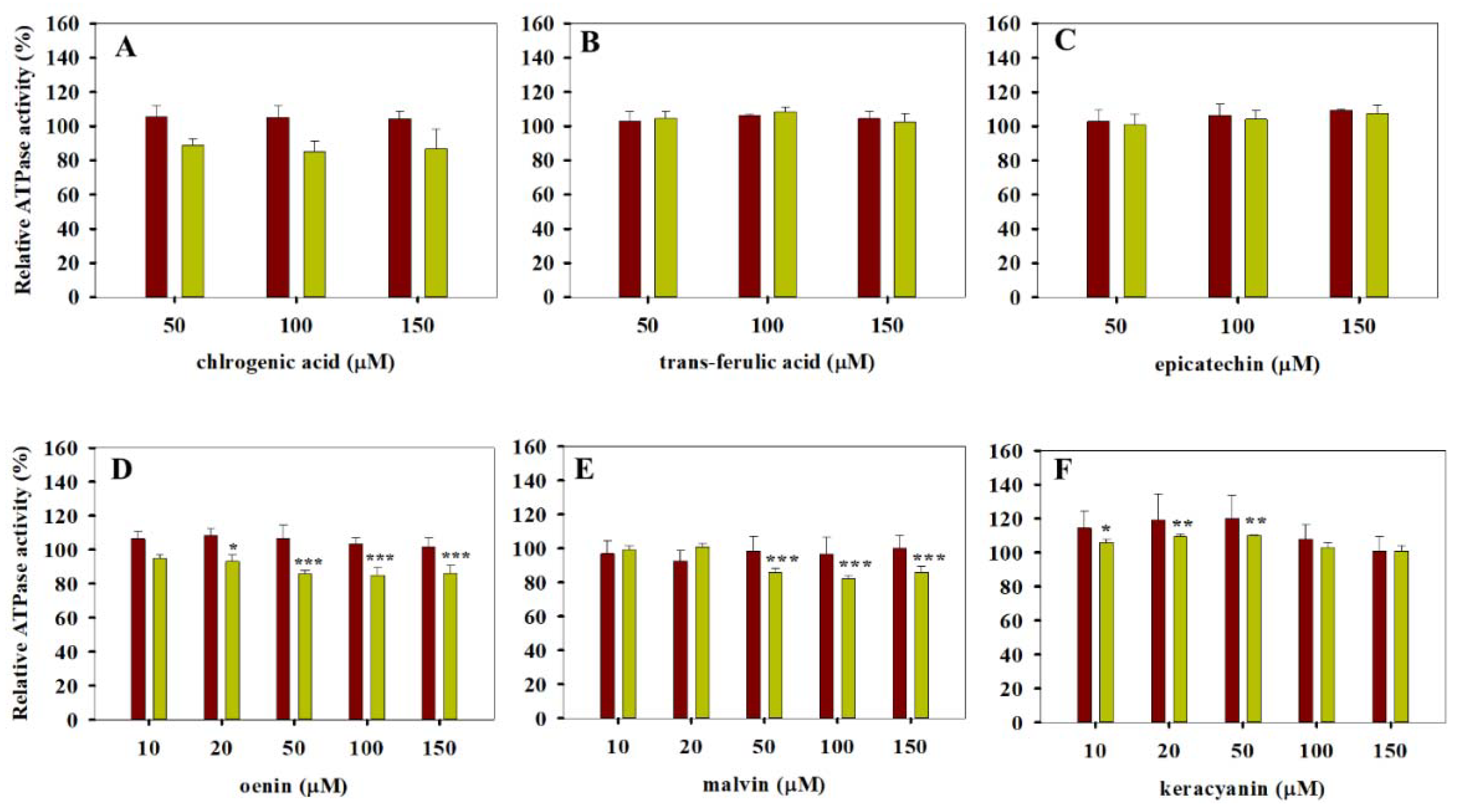
3.1. ATPase Activity Measurements
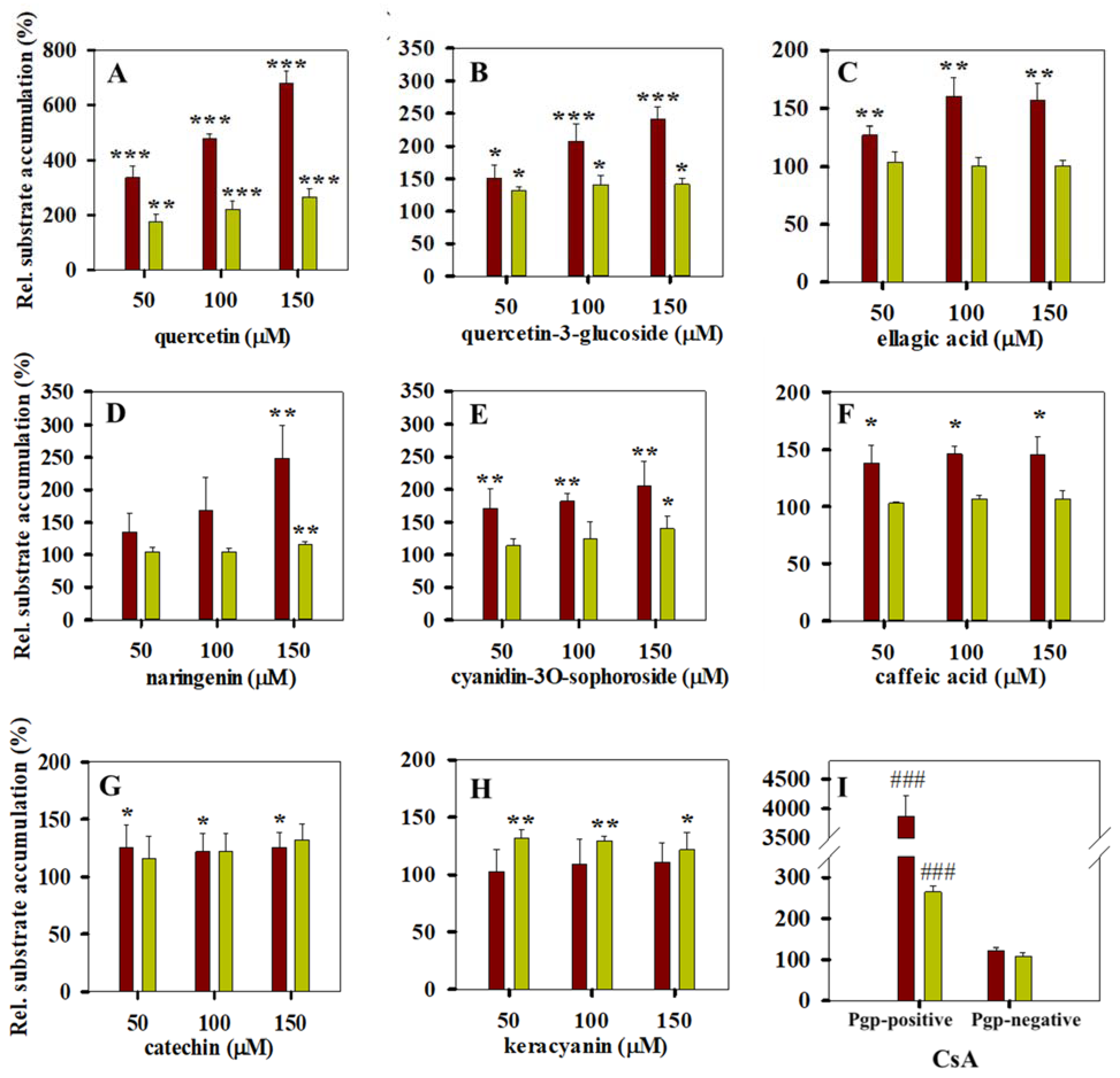
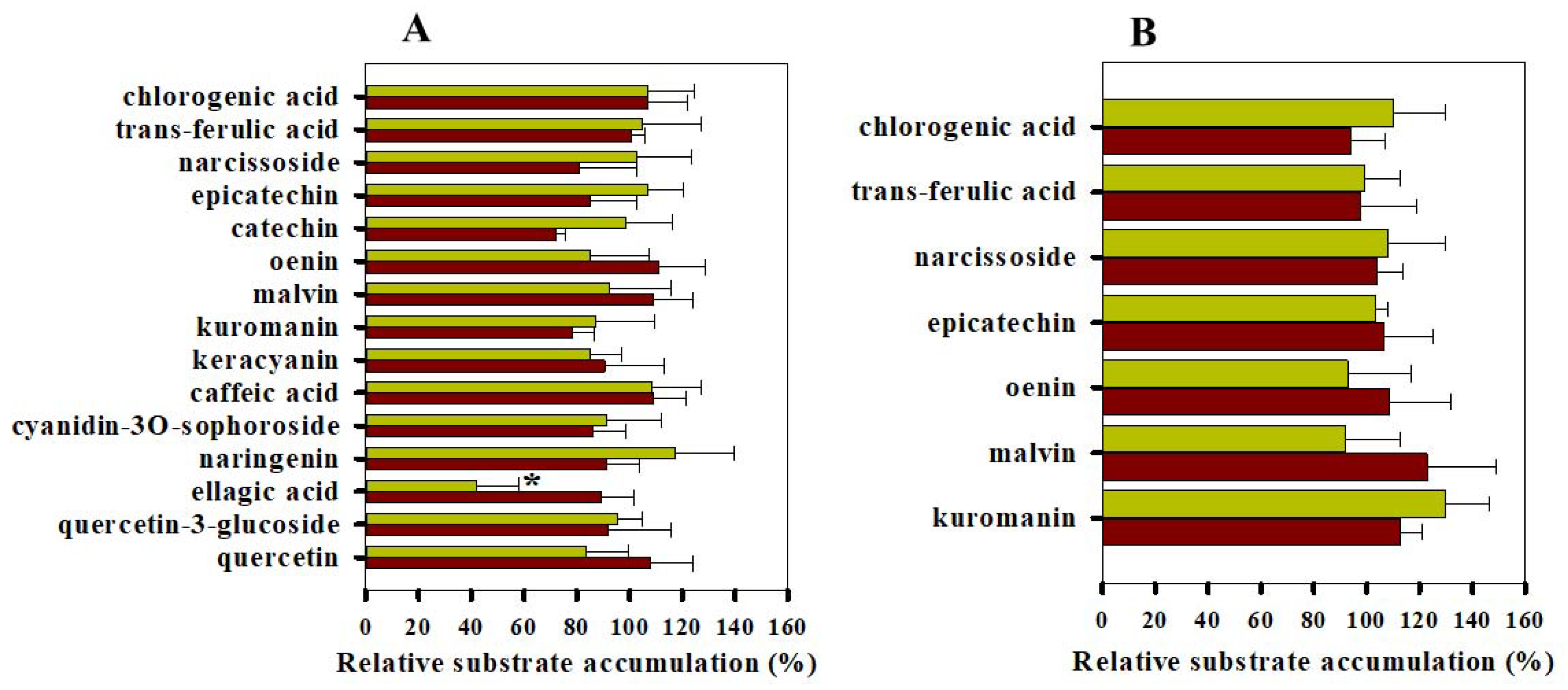
3.2. Calcein and Daunorubicin Accumulation Experiments
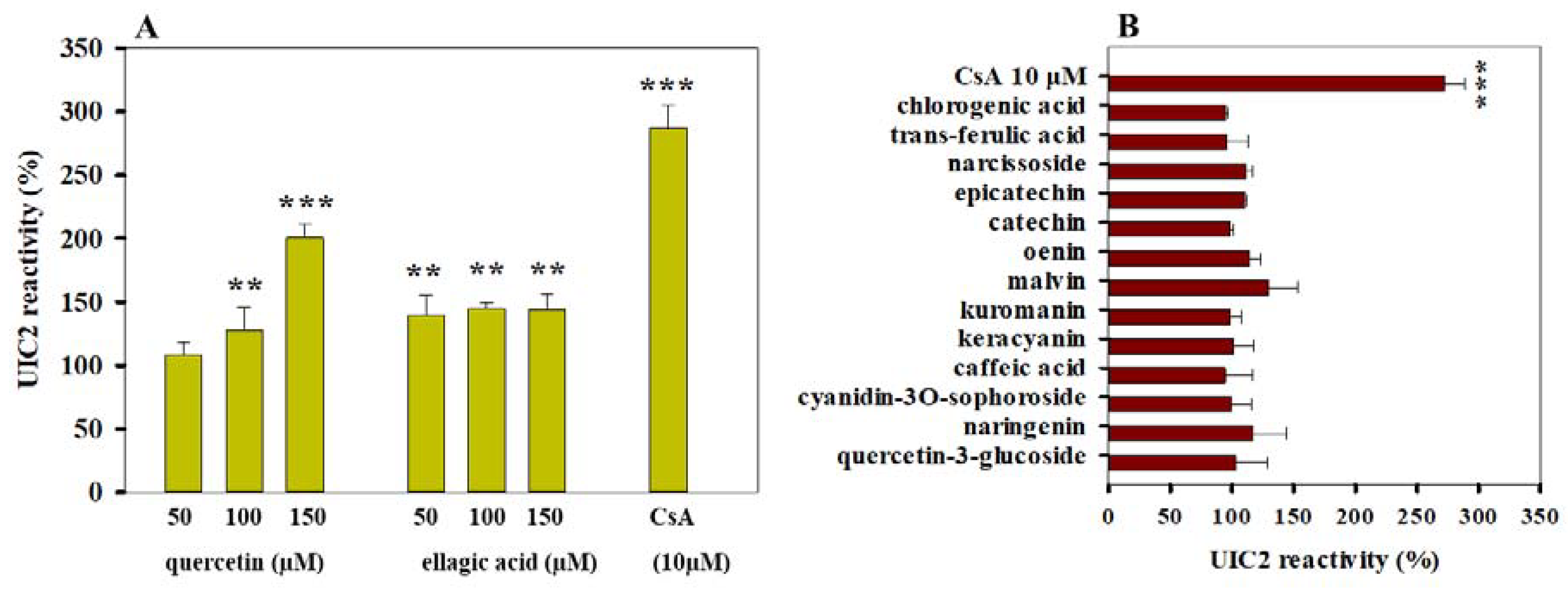
3.3. Effects of Polyphenols on the Conformation of Pgp
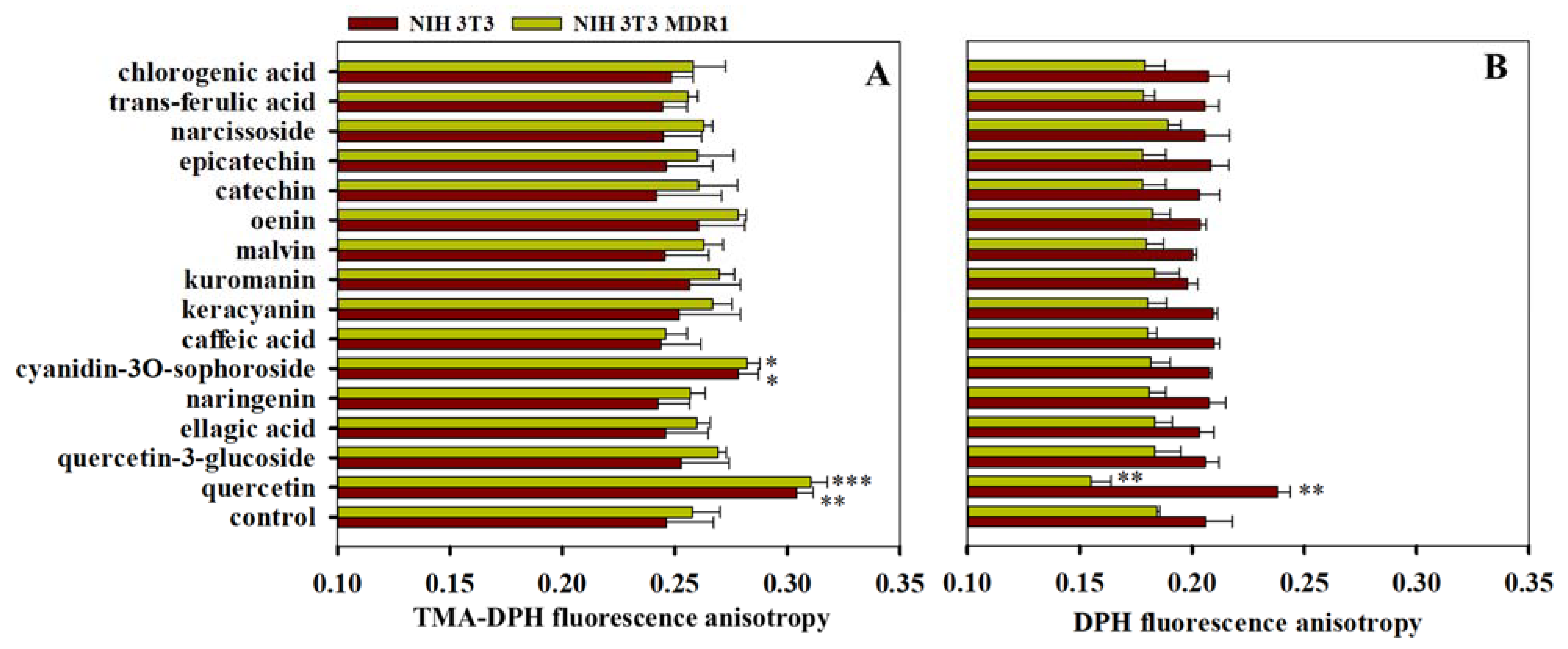
3.4. Effects of Polyphenols on Membrane Fluidity
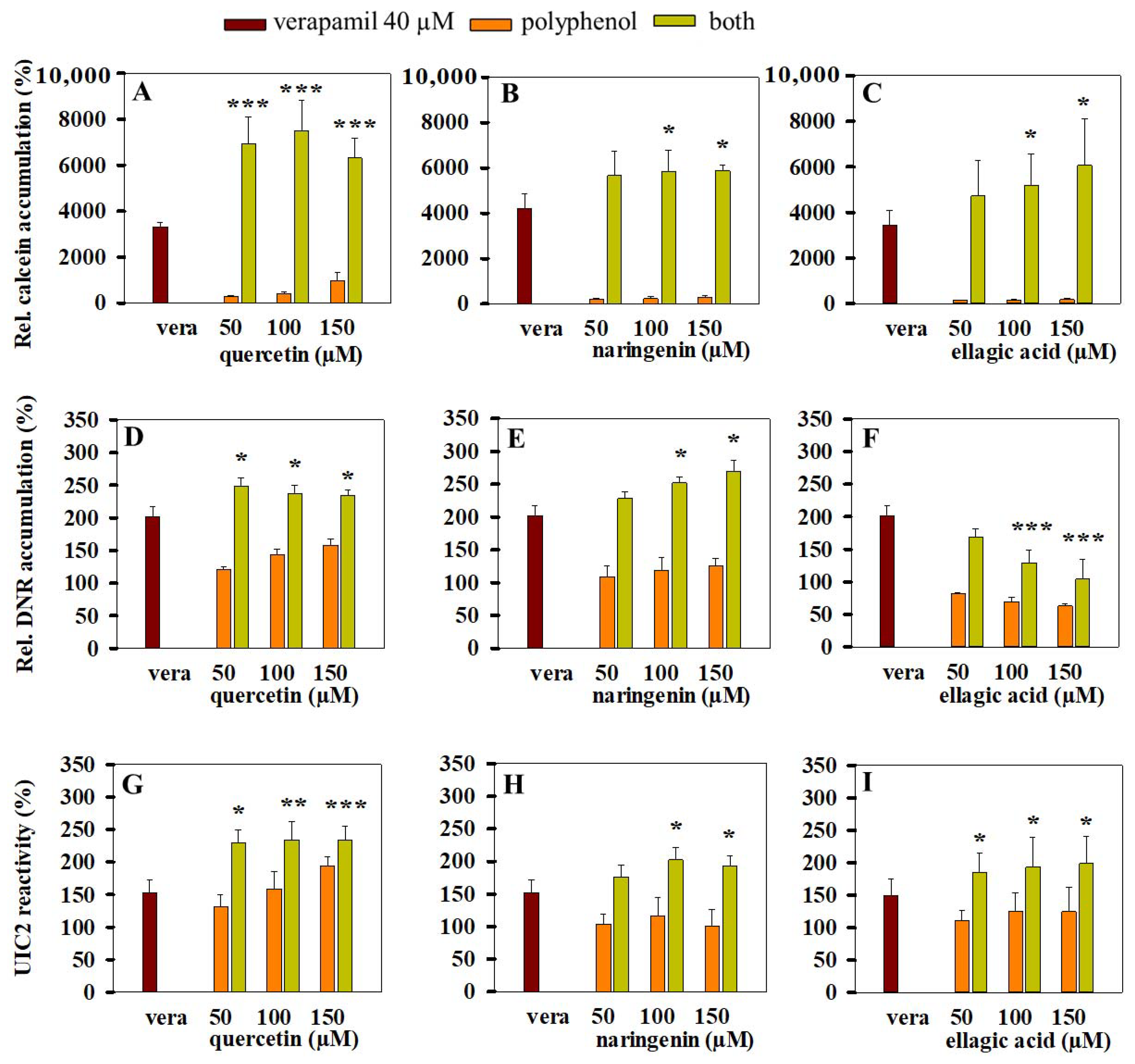
3.5. Combined Effects of Polyphenols and Verapamil on Pgp Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sarkadi, B.; Homolya, L.; Szakacs, G.; Varadi, A. Human multidrug resistance ABCB and ABCG transporters: Participation in a chemoimmunity defense system. Phys. Rev. 2006, 86, 1179–1236. [Google Scholar] [CrossRef] [PubMed]
- Goda, K.; Bacso, Z.; Szabo, G. Multidrug resistance through the spectacle of P-glycoprotein. Curr. Cancer Drug Targets 2009, 9, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Sarkadi, B.; Szakacs, G. Understanding transport through pharmacological barriers—Are we there yet? Nat. Rev. Drug Discov. 2010, 9, 897–898. [Google Scholar] [CrossRef] [Green Version]
- Szakacs, G.; Varadi, A.; Ozvegy-Laczka, C.; Sarkadi, B. The role of ABC transporters in drug absorption, distribution, metabolism, excretion and toxicity (ADME–Tox). Drug Discov. Today 2008, 13, 379–393. [Google Scholar] [CrossRef]
- Dean, M.; Fojo, T.; Bates, S. Tumour stem cells and drug resistance. Nat. Rev. Cancer 2005, 5, 275–284. [Google Scholar] [CrossRef]
- Lou, H.; Dean, M. Targeted therapy for cancer stem cells: The patched pathway and ABC transporters. Oncogene 2007, 26, 1357–1360. [Google Scholar] [CrossRef] [Green Version]
- Aller, S.G.; Yu, J.; Ward, A.; Weng, Y.; Chittaboina, S.; Zhuo, R.; Harrell, P.M.; Trinh, Y.T.; Zhang, Q.; Urbatsch, I.L.; et al. Structure of P-glycoprotein reveals a molecular basis for poly-specific drug binding. Science 2009, 323, 1718–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, I.; Blight, M.A. ABC-ATPases, adaptable energy generators fuelling transmembrane movement of a variety of molecules in organisms from bacteria to humans. J. Mol. Biol. 1999, 293, 381–399. [Google Scholar] [CrossRef]
- Seeger, M.A.; van Veen, H.W. Molecular basis of multidrug transport by ABC transporters. Biochim. Biophys. Acta-Proteins Proteom. 2009, 1794, 725–737. [Google Scholar] [CrossRef]
- Barsony, O.; Szaloki, G.; Turk, D.; Tarapcsak, S.; Gutay-Toth, Z.; Bacso, Z.; Holb, I.J.; Szekvolgyi, L.; Szabo, G.; Csanady, L.; et al. A single active catalytic site is sufficient to promote transport in P-glycoprotein. Sci. Rep. 2016, 6, 24810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, A.; Kowal, J.; Broude, E.; Roninson, I.; Locher, K.P. Structural insight into substrate and inhibitor discrimination by human P-glycoprotein. Science 2019, 363, 753–756. [Google Scholar] [CrossRef] [Green Version]
- Schinkel, A.H.; Borst, P.; Smit, J.J.M.; Wagenaar, E.; Dollé, M.; Arceci, R.J.; Baas, F.; Tsuruo, T.; Mechetner, E.B.; Roninson, I.B. Binding properties of monoclonal antibodies recognizing external epitopes of the human MDR1 P-glycoprotein. Int. J. Cancer 1993, 55, 478–484. [Google Scholar] [CrossRef]
- Vahedi, S.; Lusvarghi, S.; Pluchino, K.; Shafrir, Y.; Durell, S.R.; Gottesman, M.M.; Ambudkar, S.V. Mapping discontinuous epitopes for MRK-16, UIC2 and 4E3 antibodies to extracellular loops 1 and 4 of human P-glycoprotein. Sci. Rep. 2018, 8, 12716. [Google Scholar] [CrossRef] [PubMed]
- Mechetner, E.B.; Schott, B.; Morse, B.S.; Stein, W.D.; Druley, T.; Davis, K.A.; Tsuruo, T.; Roninson, I.B. P-glycoprotein function involves conformational transitions detectable by differential immunoreactivity. Proc. Natl. Acad. Sci. USA 1997, 94, 12908–12913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.Q.; Wang, L.; Lei, Y.; Hu, T.; Zhang, F.L.; Cho, C.H.; To, K.K.W. Reversal of P-gp and BCRP-mediated MDR by tariquidar derivatives. Eur. J. Med. Chem. 2015, 101, 560–572. [Google Scholar] [CrossRef]
- Nagy, H.; Goda, K.; Fenyvesi, F.; Bacso, Z.; Szilasi, M.; Kappelmayer, J.; Lustyik, G.; Cianfriglia, M.; Szabo, G. Distinct groups of multidrug resistance modulating agents are distinguished by competition of P-glycoprotein-specific antibodies. Biochem. Biophys. Res. Commun. 2004, 315, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Sharom, F.J. The P-glycoprotein multidrug transporter: Interactions with membrane lipids, and their modulation of activity. Biochem. Soc. Trans. 1997, 25, 1088–1096. [Google Scholar] [CrossRef]
- Sharom, F.J. Complex interplay between the P-glycoprotein multidrug efflux pump and the membrane: Its role in modulating protein function. Front. Oncol. 2014, 4, 41. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.A.; Grayer, R.J. Anthocyanins and other flavonoids. Nat. Prod. Rep. 2004, 21, 539–573. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotech. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharm. Therap. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Kellis, J.T.; Vickery, L.E. Inhibition of human estrogen synthetase (aromatase) by flavones. Science 1984, 225, 1032–1034. [Google Scholar] [CrossRef] [PubMed]
- Hertog, M.G.L.; Feskens, E.J.M.; Kromhout, D.; Hertog, M.G.L.; Hollman, P.C.H.; Hertog, M.G.L.; Katan, M.B. Dietary antioxidant flavonoids and risk of coronary heart disease: The Zutphen Elderly Study. Lancet 1993, 342, 1007–1011. [Google Scholar] [CrossRef]
- Huxley, R.R.; Neil, H.A.W. The relation between dietary flavonol intake and coronary heart disease mortality: A meta-analysis of prospective cohort studies. Eur. J Clin. Nutr. 2003, 57, 904–908. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.P.; Lee, J.; Gourley, L.; Duffy, S.W.; Day, N.E.; Estève, J. Dietary effects on breast-cancer risk in Singapore. Lancet 1991, 337, 1197–1200. [Google Scholar] [CrossRef]
- Blando, F.; Gerardi, C.; Nicoletti, I. Sour cherry (Prunus cerasus L.) anthocyanins as ingredients for functional foods. J. Biomed. Biotech. 2004, 2004, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Jaitak, V. Natural products as multidrug resistance modulators in cancer. Eur. J. Med. Chem. 2019, 176, 268–291. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Zhao, S.; Fan, X.; Chen, Y.; Zou, X.; Hu, M.; Wang, B.; Jin, J.; Wang, X.; Hu, J.; et al. Inhibitory effects of flavonoids on P-glycoprotein in vitro and in vivo: Food/herb-drug interactions and structure–activity relationships. Tox. Appl. Pharm. 2019, 369, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Damar, I.; Ekşi, A. Antioxidant capacity and anthocyanin profile of sour cherry (Prunus cerasus L.) juice. Food Chem. 2012, 135, 2910–2914. [Google Scholar] [CrossRef]
- Homoki, J.R.; Nemes, A.; Fazekas, E.; Gyemant, G.; Balogh, P.; Gal, F.; Al-Asri, J.; Mortier, J.; Wolber, G.; Babinszky, L.; et al. Anthocyanin composition, antioxidant efficiency, and α-amylase inhibitor activity of different Hungarian sour cherry varieties (Prunus cerasus L.). Food Chem. 2016, 194, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Nemes, A.; Szollosi, E.; Stundl, L.; Biro, A.; Homoki, J.R.; Szarvas, M.M.; Balogh, P.; Cziaky, Z.; Remenyik, J. Determination of flavonoid and proanthocyanidin profile of Hungarian sour cherry. Molecules 2018, 23, 3278. [Google Scholar] [CrossRef] [Green Version]
- Bruggemann, E.P.; Currier, S.J.; Gottesman, M.M.; Pastan, I. Characterization of the azidopine and vinblastine binding site of P- glycoprotein. J. Biol. Chem. 1992, 267, 21020–21026. [Google Scholar] [CrossRef]
- Sarkadi, B.; Price, E.M.; Boucher, R.C.; Germann, U.A.; Scarborough, G.A. Expression of the human multidrug resistance cDNA in insect cells generates a high activity drug-stimulated membrane ATPase. J. Biol. Chem. 1992, 267, 4854–4858. [Google Scholar] [CrossRef]
- Krasznai, Z.T.; Trencsenyi, G.; Krasznai, Z.; Mikecz, P.; Nizsaloczki, E.; Szaloki, G.; Szabo, J.P.; Balkay, L.; Marian, T.; Goda, K. 18FDG a PET tumor diagnostic tracer is not a substrate of the ABC transporter P-glycoprotein. Eur. J. Pharm. Sci. 2014, 64, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollo, Z.; Homolya, L.; Davis, C.W.; Sarkadi, B. Calcein accumulation as a fluorometric functional assay of the multidrug transporter. BBA Biomembr. 1994, 1191, 384–388. [Google Scholar] [CrossRef]
- Goda, K.; Balkay, L.; Marian, T.; Tron, L.; Aszalos, A.; Szabo, G. Intracellular pH does not affect drug extrusion by P-glycoprotein. J. Photochem. Photobiol. B Biol. 1996, 34, 177–182. [Google Scholar] [CrossRef]
- Stott, B.M.; Vu, M.P.; McLemore, C.O.; Lund, M.S.; Gibbons, E.; Brueseke, T.J.; Wilson-Ashworth, H.A.; Bell, J.D. Use of fluorescence to determine the effects of cholesterol on lipid behavior in sphingomyelin liposomes and erythrocyte membranes. J. Lipid Res. 2008, 49, 1202–1215. [Google Scholar] [CrossRef] [Green Version]
- Eskra, J.N.; Schlicht, M.J.; Bosland, M.C. Effects of black raspberries and their ellagic acid and anthocyanin constituents on taxane chemotherapy of castration-resistant prostate cancer cells. Sci. Rep. 2019, 9, 4367. [Google Scholar] [CrossRef]
- Romiti, N.; Tramonti, G.; Donati, A.; Chieli, E. Effects of grapefruit juice on the multidrug transporter P-glycoprotein in the human proximal tubular cell line HK-2. Life Sci. 2004, 76, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Goda, K.; Fenyvesi, F.; Bacso, Z.; Nagy, H.; Marian, T.; Megyeri, A.; Krasznai, Z.; Juhasz, I.; Vecsernyes, M.; Szabo, G. Complete inhibition of p-glycoprotein by simultaneous treatment with a distinct class of modulators and the UIC2 monoclonal antibody. J. Pharmacol. Exp. Therap. 2007, 320, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Muzi, M.; Mankoff, D.A.; Link, J.M.; Shoner, S.; Collier, A.C.; Sasongko, L.; Unadkat, J.D. Imaging of cyclosporine inhibition of P-glycoprotein activity using 11C-verapamil in the brain: Studies of healthy humans. J. Nucl. Med. 2009, 50, 1267–1275. [Google Scholar] [CrossRef] [Green Version]
- Prendergast, F.G.; Callahan, P.J.; Haugland, R.P. 1-[4-(Trimethylamino)phenyl]-6-phenylhexa-1,3,5-triene: Synthesis, fluorescence properties, and use as a fluorescence probe of lipid bilayers. Biochemistry 1981, 20, 7333–7338. [Google Scholar] [CrossRef]
- Do Canto, A.M.T.M.; Robalo, J.R.; Santos, P.D.; Carvalho, A.J.P.; Ramalho, J.P.P.; Loura, L.M.S. Diphenylhexatriene membrane probes DPH and TMA-DPH: A comparative molecular dynamics simulation study. Biochim. Biophys. Acta-Biomembr. 2016, 1858, 2647–2661. [Google Scholar] [CrossRef]
- Kessel, D. Probing membrane alterations associated with anthracycline resistance using fluorescent dyes. Biochem. Pharmacol. 1988, 37, 4253–4256. [Google Scholar] [CrossRef]
- Tarapcsak, S.; Szaloki, G.; Telbisz, A.; Gyongy, Z.; Matuz, K.; Csosz, E.; Nagy, P.; Holb, I.J.; Ruhl, R.; Nagy, L.; et al. Interactions of retinoids with the ABC transporters P-glycoprotein and breast cancer resistance protein. Sci. Rep. 2017, 7, 41376. [Google Scholar] [CrossRef] [PubMed]
- Teel, R.W. Ellagic acid binding to DNA as a possible mechanism for its antimutagenic and anticarcinogenic action. Cancer Lett. 1986, 30, 329–336. [Google Scholar] [CrossRef]
- Morris, M.E.; Zhang, S. Flavonoid-drug interactions: Effects of flavonoids on ABC transporters. Life Sci. 2006, 78, 2116–2130. [Google Scholar] [CrossRef]
- Nagy, H.; Goda, K.; Arceci, R.; Cianfriglia, M.; Mechetner, E.; Szabo, G. P-glycoprotein conformational changes detected by antibody competition. Eur. J. Biochem. 2001, 268, 2416–2420. [Google Scholar] [CrossRef] [Green Version]
- Aänismaa, P.; Gatlik-Landwojtowicz, E.; Seelig, A. P-glycoprotein senses its substrates and the lateral membrane packing density: Consequences for the catalytic cycle. Biochemistry 2008, 47, 10197–11207. [Google Scholar] [CrossRef]
- Ollila, F.; Halling, K.; Vuorela, P.; Vuorela, H.; Slotte, J.P. Characterization of flavonoid-biomembrane interactions. Arch. Biochem. Biophys. 2002, 399, 103–108. [Google Scholar] [CrossRef]
- Strugała, P.; Tronina, T.; Huszcza, E.; Gabrielska, J. Bioactivity in vitro of quercetin glycoside obtained in Beauveria bassiana culture and its interaction with liposome membranes. Molecules 2017, 22, 1520. [Google Scholar] [CrossRef] [Green Version]
- Ulrih, N.P.; Ota, A.; Šentjurc, M.; Kure, S.; Abram, V. Flavonoids and cell membrane fluidity. Food Chem. 2010, 121, 78–84. [Google Scholar] [CrossRef]
- Murota, K.; Shimizu, S.; Chujo, H.; Moon, J.H.; Terao, J. Efficiency of absorption and metabolic conversion of quercetin and its glucosides in human intestinal cell line Caco-2. Arch. Biochem. Biophys. 2000, 384, 391–397. [Google Scholar] [CrossRef]
- De Granada-Flor, A.; Sousa, C.; Filipe, H.A.L.; Santos, M.S.C.S.; De Almeida, R.F.M. Quercetin dual interaction at the membrane level. Chem. Comm. 2019, 55, 1750–1753. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Egido, E.; Li-Blatter, X.; Müller, R.; Merino, G.; Bernèche, S.; Seelig, A. Allocrite sensing and binding by the breast cancer resistance protein (ABCG2) and P-glycoprotein (ABCB1). Biochemistry 2015, 54, 6195–6206. [Google Scholar] [CrossRef]
- Sharom, F.J.; Yu, X.; Doige, C.A. Functional reconstitution of drug transport and ATPase activity in proteoliposomes containing partially purified P-glycoprotein. J. Biol. Chem. 1993, 268, 24197–24202. [Google Scholar] [CrossRef]
- Bohn, K.; Lange, A.; Chmielewski, J.; Hrycyna, C.A. Dual modulation of human P-glycoprotein and ABCG2 with prodrug dimers of the atypical antipsychotic agent paliperidone in a model of the blood-brain barrier. Mol. Pharm. 2017, 14, 1107–1119. [Google Scholar] [CrossRef]
- Namanja-Magliano, H.A.; Bohn, K.; Agrawal, N.; Willoughby, M.E.; Hrycyna, C.A.; Chmielewski, J. Dual inhibitors of the human blood-brain barrier drug efflux transporters P-glycoprotein and ABCG2 based on the antiviral azidothymidine. Bioorg. Med. Chem. 2017, 25, 5128–5132. [Google Scholar] [CrossRef]
- Chen, H.J.; Chung, Y.L.; Li, C.Y.; Chang, Y.T.; Wang, C.C.N.; Lee, H.Y.; Lin, H.Y.; Hung, C.C. Taxifolin resensitizes multidrug resistance cancer cells via uncompetitive inhibition of P-glycoprotein function. Molecules 2018, 23, 3055. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, K.; Tarapcsák, S.; Gyöngy, Z.; Ritter, Z.; Batta, G.; Bosire, R.; Remenyik, J.; Goda, K. Effects of Polyphenols on P-Glycoprotein (ABCB1) Activity. Pharmaceutics 2021, 13, 2062. https://doi.org/10.3390/pharmaceutics13122062
Singh K, Tarapcsák S, Gyöngy Z, Ritter Z, Batta G, Bosire R, Remenyik J, Goda K. Effects of Polyphenols on P-Glycoprotein (ABCB1) Activity. Pharmaceutics. 2021; 13(12):2062. https://doi.org/10.3390/pharmaceutics13122062
Chicago/Turabian StyleSingh, Kuljeet, Szabolcs Tarapcsák, Zsuzsanna Gyöngy, Zsuzsanna Ritter, Gyula Batta, Rosevalentine Bosire, Judit Remenyik, and Katalin Goda. 2021. "Effects of Polyphenols on P-Glycoprotein (ABCB1) Activity" Pharmaceutics 13, no. 12: 2062. https://doi.org/10.3390/pharmaceutics13122062
APA StyleSingh, K., Tarapcsák, S., Gyöngy, Z., Ritter, Z., Batta, G., Bosire, R., Remenyik, J., & Goda, K. (2021). Effects of Polyphenols on P-Glycoprotein (ABCB1) Activity. Pharmaceutics, 13(12), 2062. https://doi.org/10.3390/pharmaceutics13122062