
TLR Agonists as Vaccine Adjuvants Targeting Cancer and Infectious Diseases


Abstract
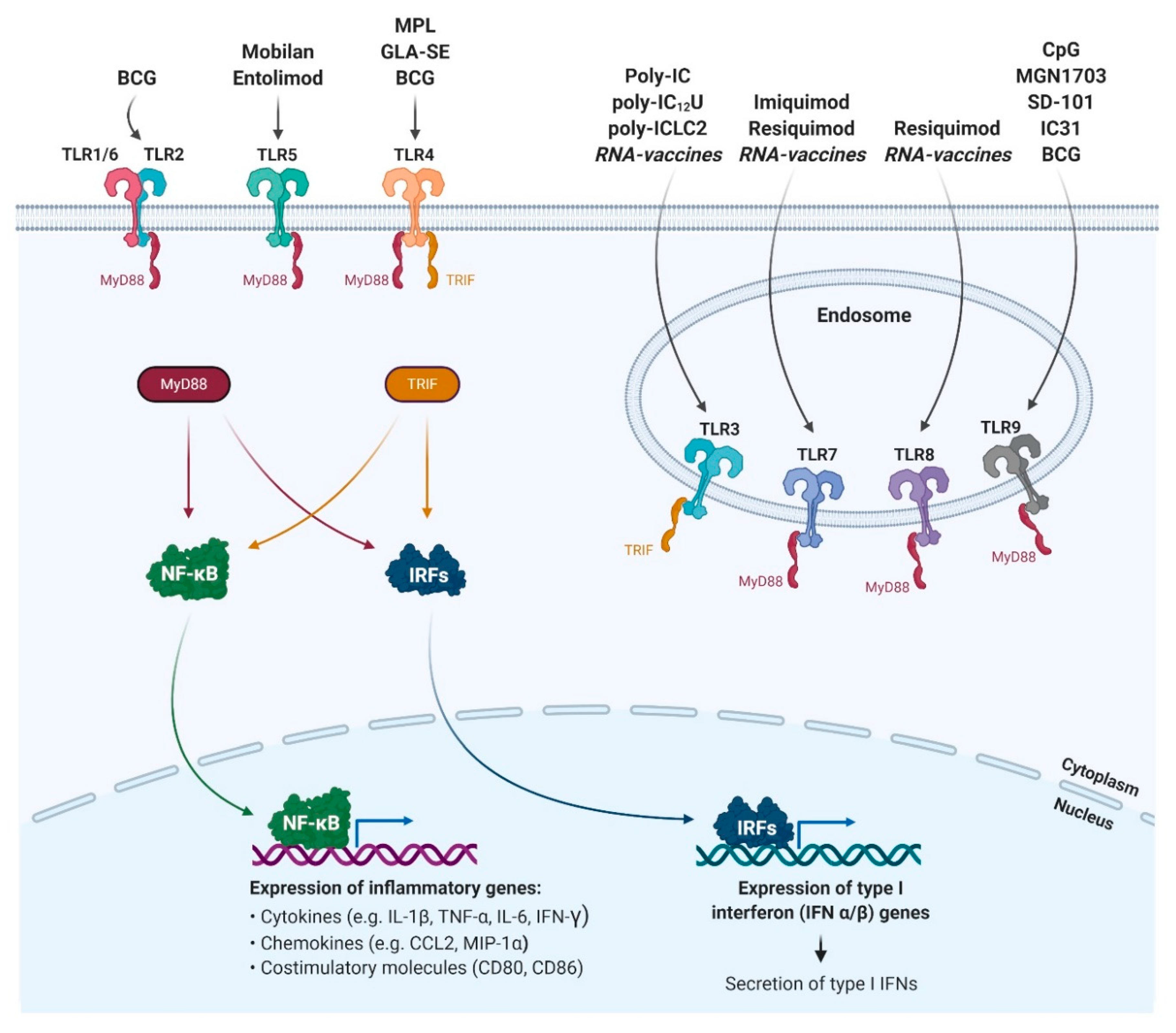
:1. Historical Perspective
2. TLR3 Agonists
Poly-IC and Derivates
3. TLR4 Agonists
3.1. Monophosphoryl Lipid (MPL)
3.2. Glucopyranosyl Lipid A in a Stable Emulsion (GLA-SE)
4. TLR5 Agonists
4.1. Mobilan
4.2. Entolimod
5. TLR7 Agonists
5.1. Resiquimod
5.2. Imiquimod
6. TLR9 Agonists
6.1. CpG
6.2. MGN1703
6.3. SD-101
6.4. IC31
7. Vaccines that Work as TLR Agonists
7.1. BCG Vaccine
7.2. RNA-Vaccines
8. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Luke, A.J.O.N.; Douglas, G.; Andrew, G.B. The history of Toll-like receptors—Redefining innate immunity. Nat. Rev. Immunol. 2013, 13, 453. [Google Scholar] [CrossRef]
- Janeway, C.A. Pillars article: Approaching the asymptote? Evolution and revolution in immunology. Cold spring harb symp quant biol. 1989. 54: 1–13. J. Immunol. 2013, 191, 4475. [Google Scholar] [PubMed]
- Poltorak, A.; He, X.; Smirnova, I.; Liu, M.-Y.; Van Huffel, C.; Du, X.; Birdwell, D.; Alejos, E.; Silva, M.; Galanos, C.; et al. Defective LPS Signaling in C3H/HeJ and C57BL/10ScCr Mice: Mutations in Tlr4 Gene. Science 1998, 282, 2085–2088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, K.; Takeuchi, O.; Kawai, T.; Sanjo, H.; Ogawa, T.; Takeda, Y.; Takeda, K.; Akira, S. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: Evidence for TLR4 as the Lps gene product. J. Immunol. 1999, 162, 3749. [Google Scholar]
- Hiroaki, H.; Osamu, T.; Taro, K.; Tsuneyasu, K.; Shintaro, S.; Hideki, S.; Makoto, M.; Katsuaki, H.; Hermann, W.; Kiyoshi, T.; et al. A Toll-like receptor recognizes bacterial DNA. Nature 2000, 408, 740. [Google Scholar] [CrossRef]
- Hiroaki, H.; Tsuneyasu, K.; Osamu, T.; Shintaro, S.; Hideki, S.; Katsuaki, H.; Takao, H.; Hideyuki, T.; Kiyoshi, T.; Shizuo, A. Small anti-viral compounds activate immune cells via the TLR7 MyD88–dependent signaling pathway. Nat. Immunol. 2002, 3, 196. [Google Scholar] [CrossRef]
- Jorge, L.C.; Bennett, W.; Chaitali, B.; Juan, C.S. TLR8: The forgotten relative revindicated. Cell. Mol. Immunol. 2012, 9, 434. [Google Scholar] [CrossRef] [Green Version]
- Frasca, L.; Lande, R. Toll-like receptors in mediating pathogenesis in systemic sclerosis. Clin. Exp. Immunol. 2020, 201, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Momtazmanesh, S.; Perry, G.; Rezaei, N. Toll-like receptors in Alzheimer’s disease. J. Neuroimmunol. 2020, 348. [Google Scholar] [CrossRef]
- Valentina, C.; Maria Cecilia, G. Microbiome-Gut-Brain Axis and Toll-Like Receptors in Parkinson’s Disease. Int. J. Mol. Sci. 2018, 19, 1689. [Google Scholar] [CrossRef] [Green Version]
- Drouin-Ouellet, J.; St-Amour, I.; Saint-Pierre, M.; Lamontagne-Proulx, J.; Kriz, J.; Barker, R.A.; Cicchetti, F. Toll-like receptor expression in the blood and brain of patients and a mouse model of Parkinson’s disease. Int. J. Neuropsychopharmacol. 2014, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzamko, N.; Inesta-Vaquera, F.; Zhang, J.; Xie, C.; Cai, H.; Arthur, S.; Choi, H.; Cohen, P.; Pedrioli, P.; Clark, K.; et al. The IkappaB Kinase Family Phosphorylates the Parkinson’s Disease Kinase LRRK2 at Ser935 and Ser910 during Toll-Like Receptor Signaling. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noelker, C.; Morel, L.; Lescot, T.; Osterloh, A.; Alvarez-Fischer, D.; Breloer, M.; Henze, C.; Depboylu, C.; Skrzydelski, D.; Michel, P.P.; et al. Toll like receptor 4 mediates cell death in a mouse MPTP model of Parkinson disease. Sci. Rep. 2013, 3, 1393. [Google Scholar] [CrossRef] [PubMed]
- Codolo, G.; Plotegher, N.; Pozzobon, T.; Brucale, M.; Tessari, I.; Bubacco, L.; de Bernard, M. Triggering of Inflammasome by Aggregated [alpha]-Synuclein, an Inflammatory Response in Synucleinopathies.(Research Article). PLoS ONE 2013, 8, e55375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Moynagh, P.N. TLR signalling and activation of IRFs: Revisiting old friends from the NF-kappaB pathway. Trends Immunol. 2005, 26, 469–476. [Google Scholar] [CrossRef]
- Zhao, G.N.; Jiang, D.S.; Li, H. Interferon regulatory factors: At the crossroads of immunity, metabolism, and disease. Biochim. Biophys. Acta 2015, 1852, 365–378. [Google Scholar] [CrossRef] [Green Version]
- FDA Approves Cervarix, GlaxoSmithKline’s Cervical Cancer Vaccine. Available online: https://www.gsk.com/en-gb/media/press-releases/fda-approves-cervarix-glaxosmithkline-s-cervical-cancer-vaccine/# (accessed on 17 November 2020).
- Fox, C.B. Development of the CpG Adjuvant 1018: A Case Study. In Vaccine Adjuvants: Methods and Protocols; Springer: New York, NY, USA, 2017; p. 13. [Google Scholar]
- Robinson, R.A.; Devita, V.T.; Levy, H.B.; Baron, S.; Hubbard, S.P.; Levine, A.S. A phase I-II trial of multiple-dose polyriboinosic-polyribocytidylic acid in patieonts with leukemia or solid tumors. J. Natl. Cancer Inst. 1976, 57, 599. [Google Scholar] [CrossRef]
- Martins, K.A.; Bavari, S.; Salazar, A.M. Vaccine adjuvant uses of poly-IC and derivatives. Expert Rev. Vaccines 2015, 14, 447–459. [Google Scholar] [CrossRef]
- Thompson, K.; Strayer, D.; Salvato, P.; Thompson, C.; Klimas, N.; Molavi, A.; Hamill, A.; Zheng, Z.; Ventura, D.; Carter, W. Results of a double-blind placebo-controlled study of the double-stranded RNA drug polyI:PolyC 12 U in the treatment of HIV infection. Eur. J. Clin. Microbiol. Infect. Dis. 1996, 15, 580–587. [Google Scholar] [CrossRef]
- Strayer, D.R.; Carter, W.A.; Stouch, B.C.; Stevens, S.R.; Bateman, L.; Cimoch, P.J.; Lapp, C.W.; Peterson, D.L.; Mitchell, W.M. A Double-Blind, Placebo-Controlled, Randomized, Clinical Trial of the TLR-3 Agonist Rintatolimod in Severe Cases of Chronic Fatigue Syndrome (Treatment of CFS with a TLR-3 Agonist). PLoS ONE 2012, 7, e31334. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.E.B.; Haney, E.; McDonagh, M.; Pappas, M.; Daeges, M.; Wasson, N.; Fu, R.; Nelson, H.D. Treatment of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome: A Systematic Review for a National Institutes of Health Pathways to Prevention Workshop. Ann. Intern. Med. 2015, 162, 841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iribarren, K.; Bloy, N.; Buqué, A.; Cremer, I.; Eggermont, A.; Fridman, W.H.; Fucikova, J.; Galon, J.; Špíšek, R.; Zitvogel, L.; et al. Trial Watch: Immunostimulation with Toll-like receptor agonists in cancer therapy. Oncoimmunology 2015, 5, e1088631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasani, B.; Navabi, H.; Adams, M. Ampligen: A potential toll-like 3 receptor adjuvant for immunotherapy of cancer. Vaccine 2009, 27, 3401–3404. [Google Scholar] [CrossRef]
- Salazar, A.M.; Levy, H.B.; Ondra, S.; Kende, M.; Scherokman, B.; Brown, D.; Mena, H.; Martin, N.; Schwab, K.; Donovan, D.; et al. Long-term treatment of malignant gliomas with intramuscularly administered polyinosinic-polycytidylic acid stabilized with polylysine and carboxymethylcellulose: An open pilot study. Neurosurgery 1996, 38, 1096. [Google Scholar] [CrossRef]
- Sabbatini, P.; Tsuji, T.; Ferran, L.; Ritter, E.; Sedrak, C.; Tuballes, K.; Jungbluth, A.A.; Ritter, G.; Aghajanian, C.; Bell-Mcguinn, K.; et al. Phase I trial of overlapping long peptides from a tumor self-antigen and poly-ICLC shows rapid induction of integrated immune response in ovarian cancer patients. Clin. Cancer Res. J. Am. Assoc. Cancer Res. 2012, 18, 6497. [Google Scholar] [CrossRef] [Green Version]
- Shingrix. Available online: https://www.fda.gov/vaccines-blood-biologics/vaccines/shingrix (accessed on 28 November 2019).
- Garçon, N.; Chomez, P.; Van Mechelen, M. GlaxoSmithKline Adjuvant Systems in vaccines: Concepts, achievements and perspectives. Expert Rev. Vaccines 2007, 6, 723–739. [Google Scholar] [CrossRef]
- Cervarix. Available online: https://www.fda.gov/vaccines-blood-biologics/vaccines/cervarix (accessed on 28 November 2019).
- Kundi, M. New hepatitis B vaccine formulated with an improved adjuvant system. Expert Rev. Vaccines 2007, 6, 133–140. [Google Scholar] [CrossRef]
- Fendrix. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/fendrix (accessed on 28 November 2019).
- Heineman, T.C.; Cunningham, A.; Levin, M. Understanding the immunology of Shingrix, a recombinant glycoprotein E adjuvanted herpes zoster vaccine. Curr. Opin. Immunol. 2019, 59, 42–48. [Google Scholar] [CrossRef]
- Rosewich, M.; Lee, D.; Zielen, S. Pollinex Quattro: An innovative four injections immunotherapy in allergic rhinitis. Hum. Vaccines Immunother. 2013, 9, 1523–1531. [Google Scholar] [CrossRef] [Green Version]
- Didierlaurent, A.M.; Morel, S.; Lockman, L.; Giannini, S.L.; Bisteau, M.; Carlsen, H.; Kielland, A.; Vosters, O.; Vanderheyde, N.; Schiavetti, F.; et al. AS04, an aluminum salt- and TLR4 agonist-based adjuvant system, induces a transient localized innate immune response leading to enhanced adaptive immunity. J. Immunol. 2009, 183, 6186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillet, S.; Aubin, É.; Trépanier, S.; Poulin, J.-F.; Yassine-Diab, B.; ter Meulen, J.; Ward, B.; Landry, N. Humoral and cell-mediated immune responses to H5N1 plant-made virus-like particle vaccine are differentially impacted by alum and GLA-SE adjuvants in a Phase 2 clinical trial. NPJ Vaccines 2018, 3, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coler, R.N.; Day, T.A.; Ellis, R.; Piazza, F.M.; Beckmann, A.M.; Vergara, J.; Rolf, T.; Lu, L.; Alter, G.; Hokey, D.; et al. The TLR-4 agonist adjuvant, GLA-SE, improves magnitude and quality of immune responses elicited by the ID93 tuberculosis vaccine: First-in-human trial. NPJ Vaccines 2018, 3, 34. [Google Scholar] [CrossRef] [PubMed]
- Somaiah, N.; Chawla, S.P.; Block, M.S.; Morris, J.C.; Do, K.T.; Kim, J.W.; Druta, M.; Sankhala, K.K.; Hwu, P.; Gnjatic, S.; et al. Immune response, safety, and survival impact from CMB305 in NY-ESO-1+ recurrent soft tissue sarcomas (STS). J. Clin. Oncol. 2017, 35, 11006. [Google Scholar] [CrossRef]
- Bhatia, S.; Miller, N.J.; Lu, H.; Longino, N.V.; Ibrani, D.; Shinohara, M.M.; Byrd, D.R.; Parvathaneni, U.; Kulikauskas, R.; Ter Meulen, J.; et al. Intratumoral G100, a TLR4 Agonist, Induces Antitumor Immune Responses and Tumor Regression in Patients with Merkel Cell Carcinoma. Clin. Cancer Res. J. Am. Assoc. Cancer Res. 2019, 25, 1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coler, R.N.; Baldwin, S.L.; Shaverdian, N.; Bertholet, S.; Reed, S.J.; Raman, V.S.; Lu, X.; DeVos, J.; Hancock, K.; Katz, J.M.; et al. A Synthetic Adjuvant to Enhance and Expand Immune Responses to Influenza Vaccines. PLoS ONE 2010, 5, e13677. [Google Scholar] [CrossRef] [Green Version]
- Mett, V.; Komarova, E.A.; Greene, K.; Bespalov, I.; Brackett, C.; Gillard, B.; Gleiberman, A.S.; Toshkov, I.A.; Aygün-Sunar, S.; Johnson, C.; et al. Mobilan: A recombinant adenovirus carrying Toll-like receptor 5 self-activating cassette for cancer immunotherapy. Oncogene 2018, 37, 439. [Google Scholar] [CrossRef] [Green Version]
- Eremina, N.V.; Kazey, V.I.; Mishugin, S.V.; Leonenkov, R.V.; Pushkar, D.Y.; Mett, V.L.; Gudkov, A.V. First-in-human study of anticancer immunotherapy drug candidate mobilan: Safety, pharmacokinetics and pharmacodynamics in prostate cancer patients. Oncotarget 2020, 11, 1273–1288. [Google Scholar] [CrossRef]
- Dubensky, T.W.; Reed, S.G. Adjuvants for cancer vaccines. Semin. Immunol. 2010, 22, 155–161. [Google Scholar] [CrossRef]
- Bakhribah, H.; Dy, G.K.; Ma, W.W.; Zhao, Y.; Opyrchal, M.; Purmal, A.; Gollnick, S.; Brady, W.E.; Fetterly, G.J.; Ngamphaiboon, N.; et al. A phase I study of the toll-like receptor 5 (TLR5) agonist, entolimod in patients (pts) with advanced cancers. J. Clin. Oncol. 2015, 33, 3063. [Google Scholar] [CrossRef]
- Singh, V.K.; Seed, T.M. Entolimod as a radiation countermeasure for acute radiation syndrome. Drug Discov. Today 2020. [Google Scholar] [CrossRef] [PubMed]
- Vasilakos, J.P.; Tomai, M.A. The use of Toll-like receptor 7/8 agonists as vaccine adjuvants. Expert Rev. Vaccines 2013, 12, 809–819. [Google Scholar] [CrossRef]
- Sabado, R.L.; Pavlick, A.; Gnjatic, S.; Cruz, C.M.; Vengco, I.; Hasan, F.; Spadaccia, M.; Darvishian, F.; Chiriboga, L.; HSolman, R.M.; et al. Resiquimod as an immunologic adjuvant for NY-ESO-1 protein vaccination in patients with high-risk melanoma. Cancer Immunol. Res. 2015, 3, 278–287. [Google Scholar] [CrossRef] [Green Version]
- Stanley, M.A. Imiquimod and the imidazoquinolones: Mechanism of action and therapeutic potential. Clin. Exp. Dermatol. 2002, 7, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Li, J.K.; Balic, J.J.; Yu, L.; Jenkins, B. TLR Agonists as Adjuvants for Cancer Vaccines. In Regulation of Inflammatory Signaling in Health and Disease; Springer: Singapore, 2017; Volume 1024, pp. 195–212. [Google Scholar]
- Adams, S.; O’Neill, D.W.; Nonaka, D.; Hardin, E.; Chiriboga, L.; Siu, K.; Cruz, C.M.; Angiulli, A.; Angiulli, F.; Ritter, E.; et al. Immunization of malignant melanoma patients with full-length NY-ESO-1 protein using TLR7 agonist imiquimod as vaccine adjuvant. J. Immunol. 2008, 181, 776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, S.; Kozhaya, L.; Martiniuk, F.; Meng, T.-C.; Chiriboga, L.; Liebes, L.; Hochman, T.; Shuman, N.; Axelrod, D.; Speyer, J.; et al. Topical TLR7 agonist imiquimod can induce immune-mediated rejection of skin metastases in patients with breast cancer. Clin. Cancer Res. J. Am. Assoc. Cancer Res. 2012, 18, 6748. [Google Scholar] [CrossRef] [Green Version]
- Scheiermann, J.; Klinman, D.M. Clinical evaluation of CpG oligonucleotides as adjuvants for vaccines targeting infectious diseases and cancer. Vaccine 2014, 32, 6377–6389. [Google Scholar] [CrossRef] [Green Version]
- Mutwiri, G.K.; Nichani, A.K.; Babiuk, S.; Babiuk, L.A. Strategies for enhancing the immunostimulatory effects of CpG oligodeoxynucleotides. J. Control. Release 2004, 97, 1–17. [Google Scholar] [CrossRef]
- Lopez, A.M.; Hecker, R.; Mutwiri, G.; van Drunen Littel-van Den Hurk, S.; Babiuk, L.A.; Townsend, H.G.G. Formulation with CpG ODN enhances antibody responses to an equine influenza virus vaccine. Vet. Immunol. Immunopathol. 2006, 114, 103–110. [Google Scholar] [CrossRef]
- Vollmer, J.; Weeratna, R.; Payette, P.; Jurk, M.; Schetter, C.; Laucht, M.; Wader, T.; Tluk, S.; Liu, M.; Davis, H.L.; et al. Characterization of three CpG oligodeoxynucleotide classes with distinct immunostimulatory activities. Eur. J. Immunol. 2004, 34, 251–262. [Google Scholar] [CrossRef]
- Cooper, C.L.; Davis, H.L.; Angel, J.B.; Morris, M.L.; Elfer, S.M.; Seguin, I.; Krieg, A.M.; Cameron, D.W. CPG 7909 adjuvant improves hepatitis B virus vaccine seroprotection in antiretroviral-treated HIV-infected adults. Aids 2005, 19, 1473. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.D.; Wu, Y.; Martin, L.B.; Shaffer, D.; Miura, K.; Aebig, J.; Orcutt, A.; Rausch, K.; Zhu, D.; Mogensen, A.; et al. Phase 1 Study in Malaria Naïve Adults of BSAM2/Alhydrogel®+CPG 7909, a Blood Stage Vaccine against P. falciparum Malaria (Phase 1 BSAM2/Alhydrogel®+CPG 7909). PLoS ONE 2012, 7, e46094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezoe, S.; Palacpac, N.M.Q.; Tetsutani, K.; Yamamoto, K.; Okada, K.; Taira, M.; Nishida, S.; Hirata, H.; Ogata, A.; Yamada, T.; et al. First-in-human randomised trial and follow-up study of Plasmodium falciparum blood-stage malaria vaccine BK-SE36 with CpG-ODN(K3). Vaccine 2020, 38, 7246–7257. [Google Scholar] [CrossRef] [PubMed]
- Schmoll, H.; Wittig, B.; Arnold, D. Maintenance treatment with the immunomodulator MGN1703, a Toll-like receptor 9 (TLR9) agonist, in patients with metastatic colorectal carcinoma and disease control after chemotherapy: A randomised, double-blind, placebo-controlled trial. J. Cancer Res. Clin. Oncol. 2014, 140, 9. [Google Scholar] [CrossRef] [Green Version]
- Schleimann, M.H.; Kobberø, M.-L.; Vibholm, L.K.; Kjær, K.; Giron, L.B.; Busman-Sahay, K.; Chan, C.N.; Nekorchuk, M.; Schmidt, M.; Wittig, B.; et al. TLR9 agonist MGN1703 enhances B cell differentiation and function in lymph nodes. EBioMedicine 2019, 45, 328–340. [Google Scholar] [CrossRef]
- Krarup, A.R.; Abdel-Mohsen, M.; Schleimann, M.H.; Vibholm, L.; Engen, P.A.; Dige, A.; Wittig, B.; Schmidt, M.; Green, S.J.; Naqib, A.; et al. The TLR9 agonist MGN1703 triggers a potent type I interferon response in the sigmoid colon. Mucosal Immunol. 2017, 11. [Google Scholar] [CrossRef] [Green Version]
- Wittig, B.; Schmidt, M.; Scheithauer, W.; Schmoll, H.-J. MGN1703, an immunomodulator and toll-like receptor 9 (TLR-9) agonist: From bench to bedside. Crit. Rev. Oncol. Hematol. 2015, 94, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Ribas, A.; Medina, T.; Kummar, S.; Amin, A.; Kalbasi, A.; Drabick, J.J.; Barve, M.; Daniels, G.A.; Wong, D.J.; Schmidt, E.V.; et al. SD-101 in Combination with Pembrolizumab in Advanced Melanoma: Results of a Phase Ib, Multicenter Study. Cancer Discov. 2018, 8, 1250. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Campos, J.; Gallotta, M.; Gong, M.; Crain, C.; Naik, E.; Coffman, R.L.; Guiducci, C. Intratumoral injection of a CpG oligonucleotide reverts resistance to PD-1 blockade by expanding multifunctional CD8 + T cells. Proc. Natl. Acad. Sci. USA 2016, 113, E7240–E7249. [Google Scholar] [CrossRef] [Green Version]
- Frank, M.J.; Reagan, P.M.; Bartlett, N.L.; Gordon, L.I.; Friedberg, J.W.; Czerwinski, D.K.; Long, S.R.; Hoppe, R.T.; Janssen, R.; Candia, A.F.; et al. Vaccination with a TLR9 Agonist and Local Low-Dose Radiation Induces Systemic Responses in Untreated Indolent Lymphoma. Cancer Discov. 2018, 8, 1258. [Google Scholar] [CrossRef] [Green Version]
- Schellack, C.; Prinz, K.; Egyed, A.; Fritz, J.H.; Wittmann, B.; Ginzler, M.; Swatosch, G.; Zauner, W.; Kast, C.; Akira, S.; et al. IC31, a novel adjuvant signaling via TLR9, induces potent cellular and humoral immune responses. Vaccine 2006, 24, 5461–5472. [Google Scholar] [CrossRef]
- Nemes, E.; Geldenhuys, H.; Rozot, V.; Rutkowski, K.T.; Ratangee, F.; Bilek, N.; Mabwe, S.; Makhethe, L.; Erasmus, M.; Toefy, A.; et al. Prevention of M. tuberculosis Infection with H4:IC31 Vaccine or BCG Revaccination. New Engl. J. Med. 2018, 379, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Talat Iqbal, N.; Hussain, R. Non-specific immunity of BCG vaccine: A perspective of BCG immunotherapy. Trials Vaccinol. 2014, 3, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Vacchelli, E.; Galluzzi, L.; Eggermont, A.; Fridman, W.H.; Galon, J.; Sautès-Fridman, C.; Tartour, E.; Zitvogel, L.; Kroemer, G. Trial watch: FDA-approved Toll-like receptor agonists for cancer therapy. Oncoimmunology 2012, 1, 894–907. [Google Scholar] [CrossRef] [Green Version]
- Krone, B.; Kölmel, K.F.; Grange, J.M.; Mastrangelo, G.; Henz, B.M.; Botev, I.N.; Niin, M.; Seebacher, C.; Lambert, D.; Shafir, R.; et al. Impact of vaccinations and infectious diseases on the risk of melanoma—Evaluation of an EORTC case–control study. Eur. J. Cancer 2003, 39, 2372–2378. [Google Scholar] [CrossRef]
- Kölmel, K.F.; Grange, J.M.; Krone, B.; Mastrangelo, G.; Rossi, C.R.; Henz, B.M.; Seebacher, C.; Botev, I.N.; Niin, M.; Lambert, D.; et al. Prior immunisation of patients with malignant melanoma with vaccinia or BCG is associated with better survival. An European Organization for Research and Treatment of Cancer cohort study on 542 patients. Eur. J. Cancer 2005, 41, 118–125. [Google Scholar] [CrossRef]
- Kidner, T.B.; Morton, D.L.; Lee, D.J.; Hoban, M.; Foshag, L.J.; Turner, R.R.; Faries, M.B. Combined intralesional Bacille Calmette-Guérin (BCG) and topical imiquimod for in-transit melanoma. J. Immunother. 2012, 35, 716. [Google Scholar] [CrossRef] [Green Version]
- Agarwala, S.S.; Neuberg, D.; Park, Y.; Kirkwood, J.M. Mature results of a phase III randomized trial of bacillus Calmette–Guerin (BCG) versus observation and BCG plus dacarbazine versus BCG in the adjuvant therapy of American Joint Committee on Cancer Stage I–III melanoma (E1673). Cancer 2004, 100, 1692–1698. [Google Scholar] [CrossRef]
- Lamm, D.L.; Riggs, D.R.; Traynelis, C.L.; Nseyo, U.O. Apparent Failure of Current Intravesical Chemotherapy Prophylaxis to Influence the Long-Term Course of Superficial Transitional Cell Carcinoma of the Bladder. J. Urol. 1995, 153, 1444. [Google Scholar] [CrossRef]
- Herr, H.W. Intravesical BCG: Current results, natural history and implicants for urothelial cancer prevention. J. Cell. Biochem. 1992, 50, 112–119. [Google Scholar] [CrossRef]
- Gil, R.-S.; Michael, S.G.; Bernard, H.B. The mechanism of action of BCG therapy for bladder cancer—A current perspective. Nat. Rev. Urol. 2014, 11, 153. [Google Scholar] [CrossRef]
- Zhang, C.; Maruggi, G.; Shan, H.; Li, J. Advances in mRNA Vaccines for Infectious Diseases. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, D.K.; Jasny, E.; Yoon, H.; Horscroft, N.; Schanen, B.; Geter, T.; Fotin-Mleczek, M.; Petsch, B.; Wittman, V. Adjuvant effects of a sequence-engineered mRNA vaccine: Translational profiling demonstrates similar human and murine innate response. (Report). J. Transl. Med. 2017, 15, 1. [Google Scholar] [CrossRef] [Green Version]
- Norbert, P.; Michael, J.H.; Frederick, W.P.; Drew, W. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17. [Google Scholar] [CrossRef] [Green Version]
- Josh, C.; Ulrich, K.; Annette, O. Regulation of antiviral T cell responses by type I interferons. Nat. Rev. Immunol. 2015, 15, 231. [Google Scholar] [CrossRef]
- Linares-Fernández, S.; Lacroix, C.; Exposito, J.-Y.; Verrier, B. Tailoring mRNA Vaccine to Balance Innate/Adaptive Immune Response. Trends Mol. Med. 2020, 26, 311–323. [Google Scholar] [CrossRef]
- Kübler, H.; Scheel, B.; Gnad-Vogt, U.; Miller, K.; Schultze-Seemann, W.; Vom Dorp, F.; Parmiani, G.; Hampel, C.; Wedel, S.; Trojan, L.; et al. Self-adjuvanted mRNA vaccination in advanced prostate cancer patients: A first-in-man phase I/IIa study. J. Immunother. Cancer 2015, 3, 26. [Google Scholar] [CrossRef] [Green Version]
- Topol, E.J. Messenger RNA vaccines against SARS-CoV-2. Cell 2021. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. New Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and T1 T cell responses. Nature 2020, 586, 594. [Google Scholar] [CrossRef]
- Walsh, E.E.; Frenck, R.W.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based Covid-19 Vaccine Candidates. New Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. New Engl. J. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Suschak, J.J.; Wang, S.; Fitzgerald, K.A.; Lu, S. Identification of Aim2 as a sensor for DNA vaccines. J. Immunol. 2015, 194, 630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ken, J.I.; Tatsukata, K.; Shohei, K.; Kosuke, M.; Himanshu, K.; Taro, K.; Satoshi, U.; Osamu, T.; Fumihiko, T.; Cevayir, C.; et al. TANK-binding kinase-1 delineates innate and adaptive immune responses to DNA vaccines. Nature 2008, 451, 725. [Google Scholar] [CrossRef]
- Hesse, E.M.; Shimabukuro, T.T.; Su, J.R.; Hibbs, B.F.; Dooling, K.L.; Goud, R.; Lewis, P.; Ng, C.S.; Cano, M.V. Postlicensure Safety Surveillance of Recombinant Zoster Vaccine (Shingrix)—United States, October 2017-June 2018. Mmwr. Morb. Mortal. Wkly. Rep. 2019, 68, 91. [Google Scholar] [CrossRef] [PubMed]
- Haskins-Coulter, T.; Southern, J.; Andrews, N.; Miller, E. Reactogenicity of Cervarix and Gardasil human papillomavirus (HPV) vaccines in a randomized single blind trial in healthy UK adolescent females. Hum. Vaccines Immunother. 2017, 13, 1412–1420. [Google Scholar] [CrossRef]
- Fabrizi, F.; Cerutti, R.; Nardelli, L.; Tripodi, F.; Messa, P. HBV vaccination with Fendrix is effective and safe in pre-dialysis CKD population. Clin. Res. Hepatol. Gastroenterol. 2020, 44, 49–56. [Google Scholar] [CrossRef]
- Patel, P.; Salapatek, A.M.F. Pollinex® Quattro: A novel and well-tolerated, ultra short-course allergy vaccine. Expert Rev. Vaccines 2006, 5, 617–629. [Google Scholar] [CrossRef]
- Harrison, L.; Skinner, S.; Marbury, T.; Owens, M.; Kurup, S.; McKane, S.; Greene, R. Pharmacokinetics and safety of imiquimod 5% cream in the treatment of actinic keratoses of the face, scalp, or hands and arms. Arch. Dermatol. Res. 2004, 296, 6–11. [Google Scholar] [CrossRef]
- Hyer, R.; McGuire, D.K.; Xing, B.; Jackson, S.; Janssen, R. Safety of a two-dose investigational hepatitis B vaccine, HBsAg-1018, using a toll-like receptor 9 agonist adjuvant in adults. Vaccine 2018, 36, 2604–2611. [Google Scholar] [CrossRef]
- Van Den Ende, C.; Marano, C.; Van Ahee, A.; Bunge, E.M.; De Moerlooze, L. The immunogenicity and safety of GSK’s recombinant hepatitis B vaccine in adults: A systematic review of 30 years of experience. Expert Rev. Vaccines 2017, 16, 811–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luabeya, A.K.K.; Kagina, B.M.N.; Tameris, M.D.; Geldenhuys, H.; Hoff, S.T.; Shi, Z.; Kromann, I.; Hatherill, M.; Mahomed, H.; Hanekom, W.A.; et al. First-in-human trial of the post-exposure tuberculosis vaccine H56:IC31 in Mycobacterium tuberculosis infected and non-infected healthy adults. Vaccine 2015, 33, 4130–4140. [Google Scholar] [CrossRef] [PubMed]
- Lamm, D.L.; Blumenstein, B.A.; Crawford, E.D.; Montie, J.E.; Scardino, P.; Grossman, H.B.; Stanisic, T.H.; Smith, J.A.; Sullivan, J.; Sarosdy, M.F.; et al. A Randomized Trial of Intravesical Doxorubicin and Immunotherapy with Bacille Calmette–Guérin for Transitional-Cell Carcinoma of the Bladder. New Engl. J. Med. 1991, 325, 1205–1209. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Qu, S.; Chen, X.; Wu, Q.; Shi, M. Promising Targets for Cancer Immunotherapy: TLRs, RLRs, and STING-Mediated Innate Immune Pathways. Int. J. Mol. Sci. 2017, 18, 404. [Google Scholar] [CrossRef] [PubMed]
- Jennifer, K.D.; Ashley, M. Toll-like receptors: The swiss army knife of immunity and vaccine development. Clin. Transl. Immunol. 2016, 5, e85. [Google Scholar] [CrossRef]


{kind=link}
| TLR | TLR Agonist | Application | Status | Safety Profile |
|---|---|---|---|---|
| TLR3 | Poly-IC | Leukaemia and solid tumours | Phase I/II [20] | Severe toxic side effects in some patients (fever in 66%, transient elevation of serum glutamic-oxaloacetic transaminase and serum glutamic-pyruvic transaminase in 25%, minimal laboratory evidence of coagulation abnormalities in 59%, and hypersensitivity in 5%). |
| Poly-IC12U | HIV type 1 infection | Phase I. [22] | No significant adverse events (AEs). | |
| CFS/ME | Phase III [24] | No significant AEs. | ||
| Ovarian cancer and Peritoneal malignancies | Phase I ongoing (NCT01312389) [25] | N/A | ||
| Poly-ICLC | Anaplastic brain cancers | Phase I/II [27] | No significant AEs. | |
| Ovarian cancer | Phase I [28] | Generally well tolerated. | ||
| TLR4 | MPL | Herpes zoster | Approved vaccine Shingrix | Well tolerated, serious AEs are rare [91]. |
| Papillomavirus | Approved vaccine Cervarix | Localised site of injection reactogenicity reported due to the AS04 adjuvant [92]. | ||
| Hepatitis B | Approved vaccine Fendrix | Mild local and systemic AEs in up to 50% of vaccines [93]. | ||
| Allergic rhinitis | Approved immunotherapy Pollinex Quattro | Local reactogenicity in 5–6% of vaccinees and rare <0.7% systemic effects (rhinitis, breathing problems) [94]. | ||
| GLA-SE | Influenza | Phase II [37] | Generally well tolerated. | |
| Tuberculosis | Phase I [38] | Generally well tolerated. | ||
| Soft tissue sarcoma | Phase I [39] | Generally well tolerated. | ||
| Merkel cell carcinoma | First-in-human study [40] | Generally well tolerated. | ||
| TLR5 | Mobilan | Prostate cancer | Phase I [43] | Satisfactory safety and tolerability. |
| Entolimod | Advanced cancers | Phase I/investigational new drug, fast-track, and orphan drug status [45] | Some common toxicities observed but can be combined with other immunotherapeutic agents against cancer. | |
| TLR7/8 | Resiquimod | High-risk melanoma | Phase I [48] | Generally well tolerated. |
| TLR7 | Imiquimod | Genital warts, superficial basal cell carcinoma, actinic keratosis | Approved cream Aldara | Local application site reactions, minimal systemic absorption [95]. |
| Malignant melanoma | Phase I [51] | Excellent safety profile for topical application. | ||
| Breast cancer skin metastases | Phase II [52] | Transient mild to moderate local and systemic AEs. | ||
| TLR9 | CpG 1018 | Hepatitis B | Approved vaccines Heplisav-B (CpG 1018) and Engerix-B (CpG 7909) | Heplisav-B compared favourably to Engerix-B in both safety and efficacy [96]. Fatigue, headache in up to 25% of vaccines [97]. |
| CpG 7909 | Malaria | Phase I [58] | Most related adverse events were mild or moderate, but 4 volunteers experienced severe systemic reactions and two were withdrawn from vaccinations due to adverse events. | |
| MGN1703 | Metastatic colorectal carcinoma | Phase II [60] | AEs mild to moderate and limited to the injection-site. | |
| SD-101 | Advanced melanoma | Phase Ib [64] | Injection-site reactions and transient, mild-to-moderate “flu-like” AEs. | |
| SD101 | Low-grade non-Hodgkin lymphoma | Phase I/II [66] | No serious AEs. | |
| IC31 | Tuberculosis | Phase I [98] | No serious adverse events were reported. Nine subjects (36%) presented with transient cardiovascular AEs. | |
| TLR2, TLR4, TLR9 | BCG | Bladder cancer | Phase II/III, comparison to Doxorubicin [99] | Treatment with BCG resulted in more frequent but not more severe adverse reactions than doxorubicin therapy. |
| TLR2, TLR4, TLR7, TLR9 | BCG + 5% imiquimod | In-transit melanoma | Phase III [73] | Well tolerated. |
| TLR3, TLR7, TLR8 | mRNA as vaccine platform with intrinsic adjuvanting effect | Prostate cancer | Phase I/IIa [83] | SAEs considered possibly treatment-related were reported in 2 (5%) of patients (urinary retention, hydronephrosis) Resolved after symptomatic and antibiotic treatment. |
| SARS-CoV-2 | Phase II/III [84,85] | Generally well tolerated. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luchner, M.; Reinke, S.; Milicic, A. TLR Agonists as Vaccine Adjuvants Targeting Cancer and Infectious Diseases. Pharmaceutics 2021, 13, 142. https://doi.org/10.3390/pharmaceutics13020142
Luchner M, Reinke S, Milicic A. TLR Agonists as Vaccine Adjuvants Targeting Cancer and Infectious Diseases. Pharmaceutics. 2021; 13(2):142. https://doi.org/10.3390/pharmaceutics13020142
Chicago/Turabian StyleLuchner, Marina, Sören Reinke, and Anita Milicic. 2021. "TLR Agonists as Vaccine Adjuvants Targeting Cancer and Infectious Diseases" Pharmaceutics 13, no. 2: 142. https://doi.org/10.3390/pharmaceutics13020142
APA StyleLuchner, M., Reinke, S., & Milicic, A. (2021). TLR Agonists as Vaccine Adjuvants Targeting Cancer and Infectious Diseases. Pharmaceutics, 13(2), 142. https://doi.org/10.3390/pharmaceutics13020142