
Next Step in Gene Delivery: Modern Approaches and Further Perspectives of AAV Tropism Modification



Abstract
:1. Introduction
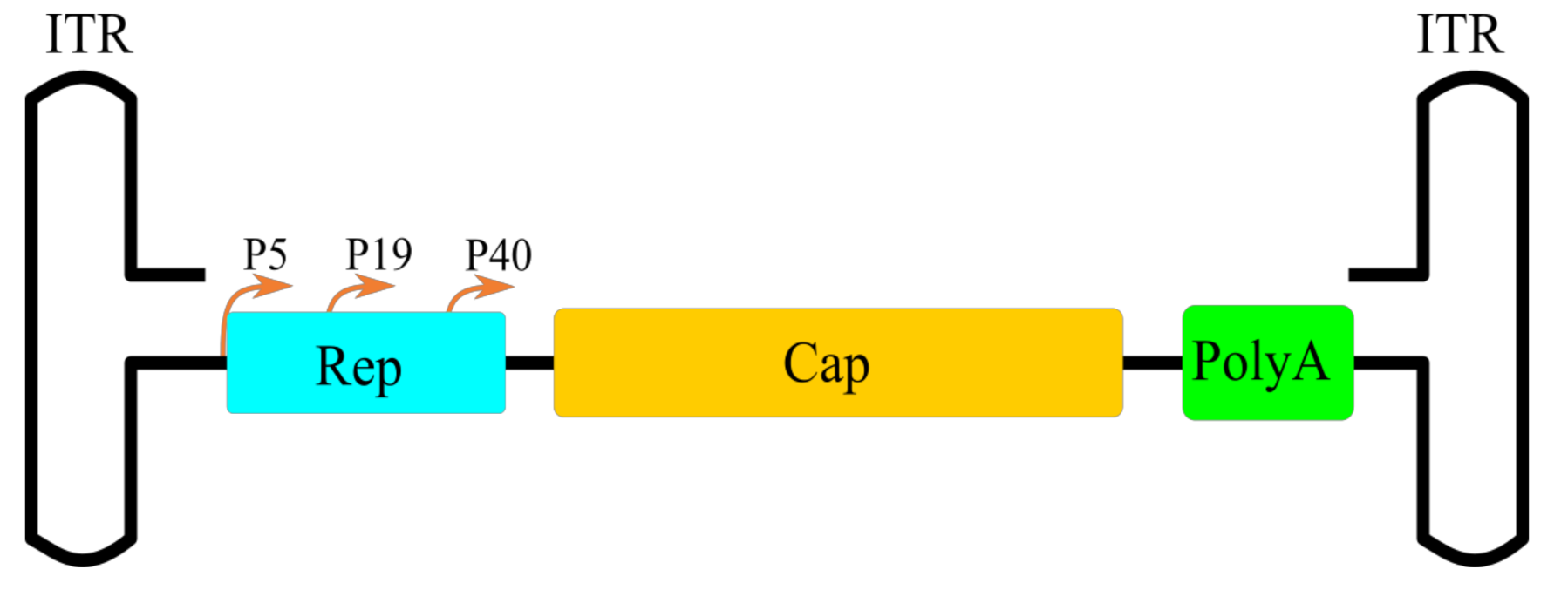
2. AAV Biology
3. Capsid Engineering for Tropism Modification
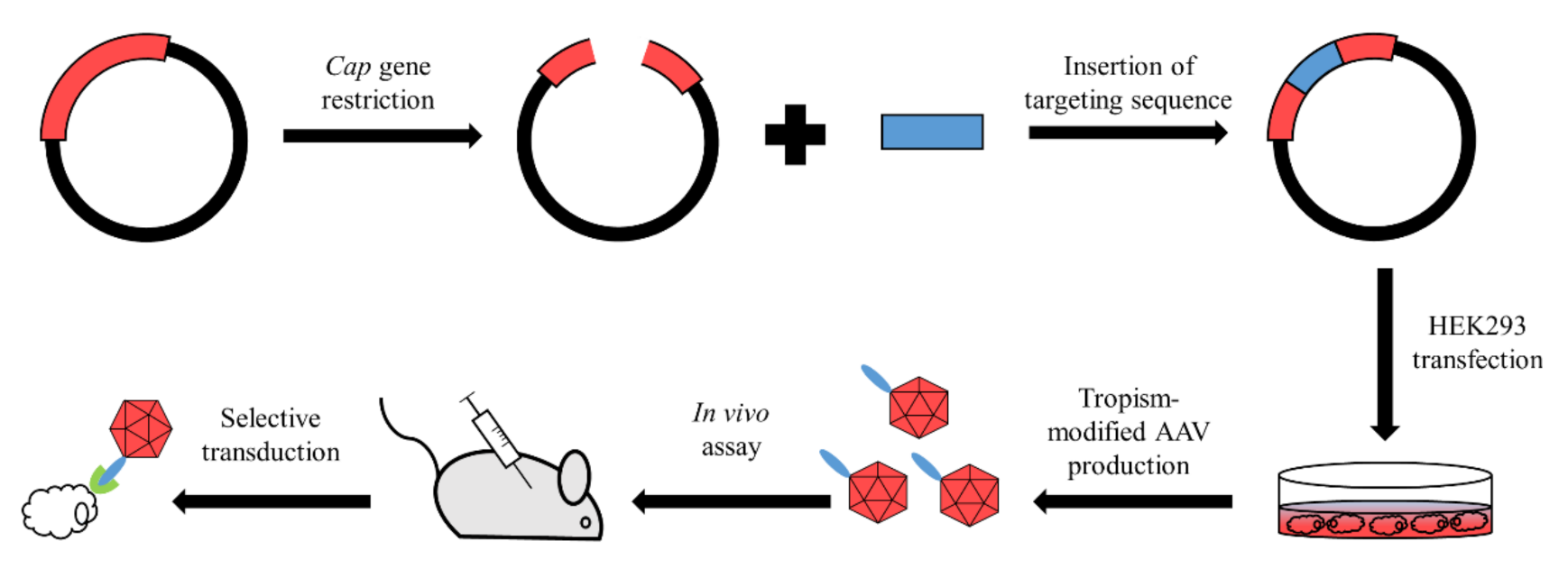
3.1. Rational Design
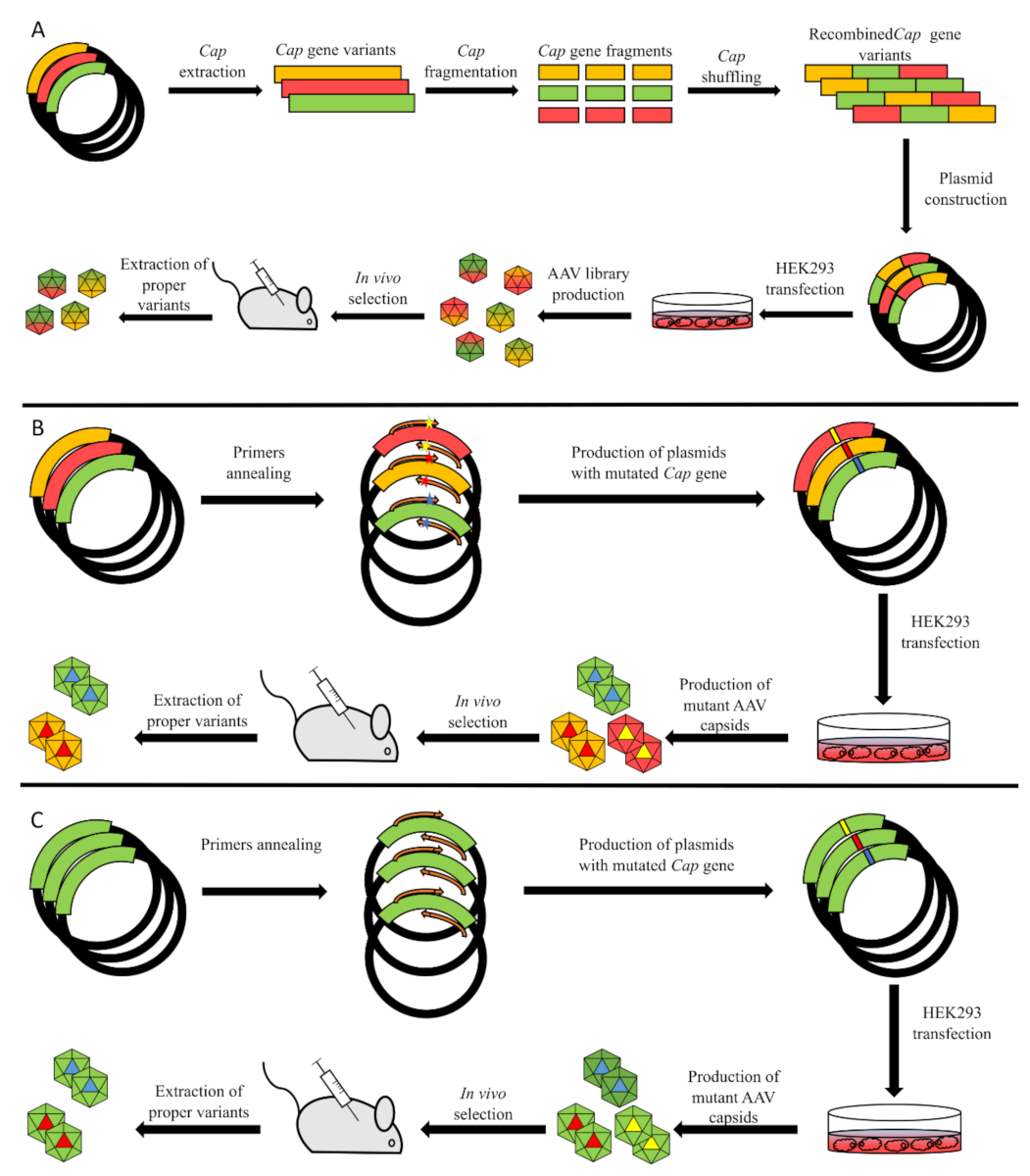
3.2. Directed Evolution
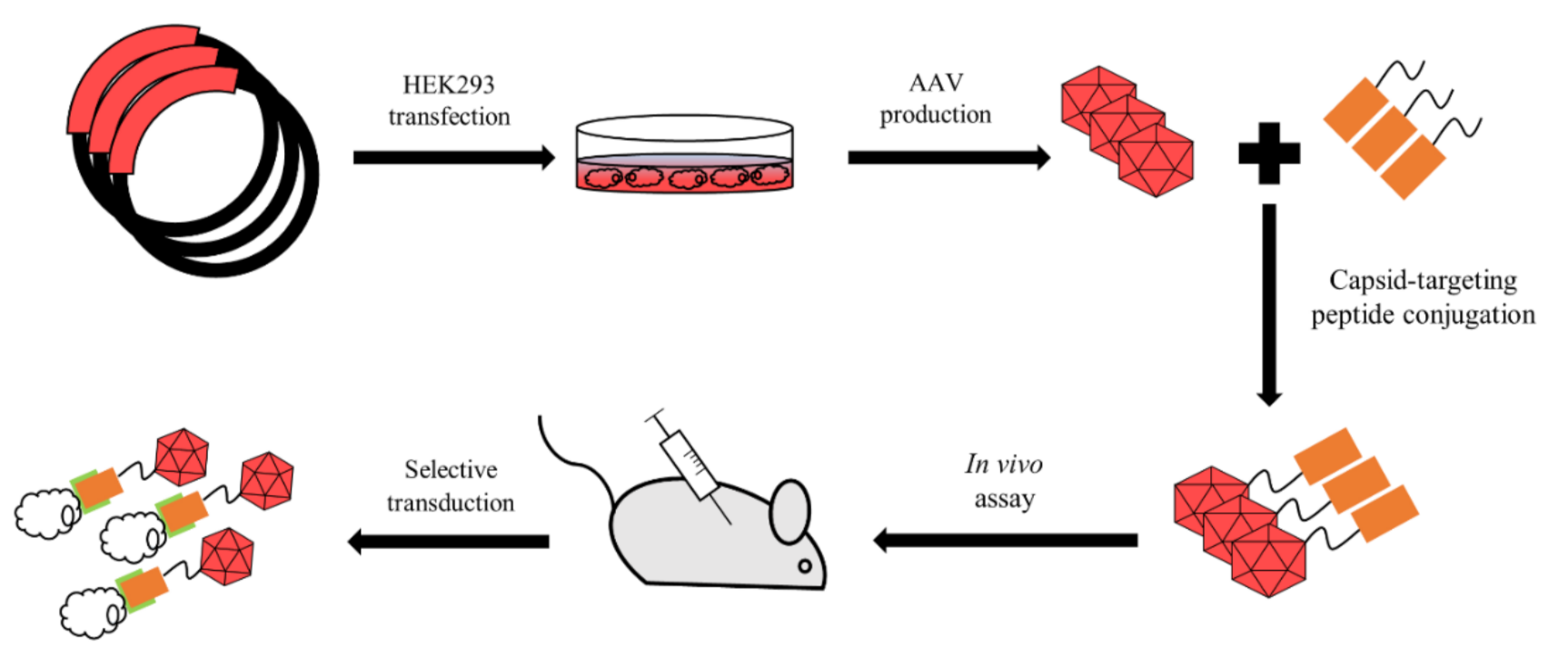
3.3. Chemical Conjugation
4. Discussion
5. Expert Opinion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lukashev, A.N.; Zamyatnin, A.A. Viral vectors for gene therapy: Current state and clinical perspectives. Biochemie 2016, 81, 700–708. [Google Scholar] [CrossRef]
- Raper, S.E.; Chirmule, N.; Lee, F.S.; Wivel, N.A.; Bagg, A.; Gao, G.P.; Wilson, J.M.; Batshaw, M.L. Fatal systemic inflammatory response syndrome in a ornithine transcarbamylase deficient patient following adenoviral gene transfer. Mol. Genet. Metab. 2003, 80, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, S.A.; Packard, B.S.; Aebersold, P.M.; Solomon, D.; Topalian, S.L.; Toy, S.T.; Simon, P.; Lotze, M.T.; Yang, J.C.; Seipp, C.A.; et al. Use of tumor-infiltrating lymphocytes and interleukin-2 in the immunotherapy of patients with metastatic melanoma. N. Engl. J. Med. 1988, 281, 1028–1034. [Google Scholar] [CrossRef]
- Hacein-Bey-Abina, S.; Garrigue, A.; Wang, G.P.; Soulier, J.; Lim, A.; Morillon, E.; Clappier, E.; Caccavelli, L.; Delabesse, E.; Beldjord, K.; et al. Insertional oncogenesis in 4 patients after retrovirus-mediated gene therapy of SCID-X1. J. Clin. Investig. 2008, 118, 3132–3142. [Google Scholar] [CrossRef] [PubMed]
- Montini, E.; Cesana, D.; Schmidt, M.; Sanvito, F.; Ponzoni, M.; Bartholomae, C.; Sergi, L.S.; Benedicenti, F.; Ambrosi, A.; di Serio, C.; et al. Hematopoietic stem cell gene transfer in a tumor-prone mouse model uncovers low genotoxicity of lentiviral vector integration. Nat. Biotechnol. 2006, 24, 687–696. [Google Scholar] [CrossRef]
- Li, C.; Samulski, R.J. Engineering adeno-associated virus vectors for gene therapy. Nat. Rev. Genet. 2020, 21, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Deviatkin, A.A.; Vakulenko, Y.A.; Akhmadishina, L.V.; Tarasov, V.V.; Beloukhova, M.I.; Zamyatnin, A.A.; Lukashev, A.N. Emerging concepts and challenges in rheumatoid arthritis gene therapy. Biomedicines 2020, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Cotmore, S.F.; Agbandje-McKenna, M.; Canuti, M.; Chiorini, J.A.; Eis-Hubinger, A.M.; Hughes, J.; Mietzsch, M.; Modha, S.; Ogliastro, M.; Pénzes, J.J.; et al. ICTV virus taxonomy profile: Parvoviridae. J. Gen. Virol. 2019, 100, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.S.; Ponnazhagan, S.; Srivastava, A. Rescue and replication signals of the adeno-associated virus 2 genome. J. Mol. Biol. 1995, 250, 573–580. [Google Scholar] [CrossRef]
- Yan, Z.; Lei-Butters, D.C.M.; Liu, X.; Zhang, Y.; Zhang, L.; Luo, M.; Zak, R.; Engelhardt, J.F. Unique biologic properties of recombinant AAV1 transduction in polarized human airway epithelia. J. Biol. Chem. 2006, 281, 29684–29692. [Google Scholar] [CrossRef] [Green Version]
- Passini, M.A.; Watson, D.J.; Vite, C.H.; Landsburg, D.J.; Feigenbaum, A.L.; Wolfe, J.H. Intraventricular brain injection of adeno-associated virus type 1 (AAV1) in neonatal mice results in complementary patterns of neuronal transduction to AAV2 and total long-term correction of storage lesions in the brains of β-glucuronidase-deficient mice. J. Virol. 2003, 77, 7034–7040. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Kido, M.; Lee, D.V.; Rabinowitz, J.E.; Samulski, R.J.; Jamieson, S.W.; Weitzman, M.D.; Thistlethwaite, P.A. Differential myocardial gene delivery by recombinant serotype-specific adeno-associated viral vectors. Mol. Ther. 2004, 10, 604–608. [Google Scholar] [CrossRef]
- Kitajima, K.; Marchadier, D.H.L.; Burstein, H.; Rader, D.J. Persistent liver expression of murine apoA-l using vectors based on adeno-associated viral vectors serotypes 5 and 1. Atherosclerosis 2006, 186, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, D.; Qian, S.; Chen, Z.; Zhu, T.; Xiao, X. Efficient and long-term intracardiac gene transfer in δ-sarcoglycan-deficiency hamster by adeno-associated virus-2 vectors. Gene Ther. 2003, 10, 1807–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, M.J.; Power, J.M.; Preovolos, A.; Mariani, J.A.; Hajjar, R.J.; Kaye, D.M. Recirculating cardiac delivery of AAV2/1SERCA2a improves myocardial function in an experimental model of heart failure in large animals. Gene Ther. 2008, 15, 1550–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, B.L.; Stein, C.S.; Heth, J.A.; Martins, I.; Kotin, R.M.; Derksen, T.A.; Zabner, J.; Ghodsi, A.; Chiorini, J.A. Recombinant adeno-associated virus type 2, 4, and 5 vectors: Transduction of variant cell types and regions in the mammalian central nervous system. Proc. Natl. Acad. Sci. USA 2000, 97, 3428–3432. [Google Scholar] [CrossRef]
- Vite, C.H.; Passini, M.A.; Haskins, M.E.; Wolfe, J.H. Adeno-associated virus vector-mediated transduction in the cat brain. Gene Ther. 2003, 10, 1874–1881. [Google Scholar] [CrossRef] [Green Version]
- Griffey, M.A.; Wozniak, D.; Wong, M.; Bible, E.; Johnson, K.; Rothman, S.M.; Wentz, A.E.; Cooper, J.D.; Sands, M.S. CNS-directed AAV2-mediated gene therapy ameliorates functional deficits in a murine model of infantile neuronal ceroid lipofuscinosis. Mol. Ther. 2006, 13, 538–547. [Google Scholar] [CrossRef]
- Ponnazhagan, S.; Mukherjee, P.; Yoder, M.C.; Wang, X.S.; Zhou, S.Z.; Kaplan, J.; Wadsworth, S.; Srivastava, A. Adeno-associated virus 2-mediated gene transfer in vivo: Organ-tropism and expression of transduced sequences in mice. Gene 1997, 190, 203–210. [Google Scholar] [CrossRef]
- Mount, J.D.; Herzog, R.W.; Tillson, D.M.; Goodman, S.A.; Robinson, N.; McCleland, M.L.; Bellinger, D.; Nichols, T.C.; Arruda, V.R.; Lothrop, C.D.; et al. Sustained phenotypic correction of hemophilia B dogs with a factor IX null mutation by liver-directed gene therapy. Blood 2002, 99, 2670–2676. [Google Scholar] [CrossRef] [Green Version]
- Moscioni, D.; Morizono, H.; McCarter, R.J.; Stern, A.; Cabrera-Luque, J.; Hoang, A.; Sanmiguel, J.; Wu, D.; Bell, P.; Gao, G.P.; et al. Long-Term correction of ammonia metabolism and prolonged survival in ornithine transcarbamylase-deficient mice following liver-directed treatment with adeno-associated viral vectors. Mol. Ther. 2006, 14, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Lisowski, L.; Dane, A.P.; Chu, K.; Zhang, Y.; Cunningham, S.C.; Wilson, E.M.; Nygaard, S.; Grompe, M.; Alexander, I.E.; Kay, M.A. Selection and evaluation of clinically relevant AAV variants in a xenograft liver model. Nature 2014, 506, 382–386. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Bell, P.; Somanathan, S.; Wang, Q.; He, Z.; Yu, H.; McMenamin, D.; Goode, T.; Calcedo, R.; Wilson, J.M. Comparative study of liver gene transfer with AAV vectors based on natural and engineered AAV capsids. Mol. Ther. 2015, 23, 1877–1887. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; Rabinowitz, J.; Provost, N.; Folliot, S.; Briot, D.; Cherel, Y.; Samulski, J.; Moullier, P. Recombinant adeno-associated virus serotype 4 mediates unique and exclusive long term transduction of retinal pigmented epithelium in rat, dog and nonhuman primate after subretinal delivery. Mol. Ther. 2003, 7, S92. [Google Scholar] [CrossRef]
- Zincarelli, C.; Soltys, S.; Rengo, G.; Rabinowitz, J.E. Analysis of AAV serotypes 1-9 mediated gene expression and tropism in mice after systemic injection. Mol. Ther. 2008, 16, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.S.; Schmidt, M.; Yan, Z.; Lindbloom, J.D.; Harding, T.C.; Donahue, B.A.; Engelhardt, J.F.; Kotin, R.; Davidson, B.L. Virus-mediated transduction of murine retina with adeno-associated virus: Effects of viral capsid and genome size. J. Virol. 2002, 76, 7651–7660. [Google Scholar] [CrossRef] [Green Version]
- Lotery, A.J.; Davidson, B.L.; Yang, G.S.; Mullins, R.F.; Russell, S.R.; Schmidt, M.; Stone, E.M.; Lindbloom, J.D.; Chiorini, J.A.; Kotin, R.M. Adeno-associated virus type 5: Transduction Efficiency and cell-type specificity in the primate retina. Hum. Gene Ther. 2003, 14, 1663–1671. [Google Scholar] [CrossRef]
- Burger, C.; Gorbatyuk, O.S.; Velardo, M.J.; Peden, C.S.; Williams, P.; Zolotukhin, S.; Reier, P.J.; Mandel, R.J.; Muzyczka, N. Recombinant AAV viral vectors pseudotyped with viral capsids from serotypes 1, 2, and 5 display differential efficiency and cell tropism after delivery to different regions of the central nervous system. Mol. Ther. 2004, 10, 302–317. [Google Scholar] [CrossRef]
- Colle, M.A.; Piguet, F.; Bertrand, L.; Raoul, S.; Bieche, I.; Dubreil, L.; Sloothaak, D.; Bouquet, C.; Moullier, P.; Aubourg, P.; et al. Efficient intracerebral delivery of AAV5 vector encoding human ARSA in non-human primate. Hum. Mol. Genet. 2009, 19, 147–158. [Google Scholar] [CrossRef]
- Pañeda, A.; Vanrell, L.; Mauleon, I.; Crettaz, J.S.; Berraondo, P.; Timmermans, E.J.; Beattie, S.G.; Twisk, J.; van Deventer, S.; Prieto, J.; et al. Effect of adeno-associated virus serotype and genomic structure on liver transduction and biodistribution in mice of both genders. Hum. Gene Ther. 2009, 20, 908–917. [Google Scholar] [CrossRef] [PubMed]
- Towne, C.; Raoul, C.; Schneider, B.L.; Aebischer, P. Systemic AAV6 delivery mediating RNA interference against SOD1: Neuromuscular transduction does not alter disease progression in fALS mice. Mol. Ther. 2008, 16, 1018–1025. [Google Scholar] [CrossRef]
- Jiang, H.; Lillicrap, D.; Patarroyo-White, S.; Liu, T.; Qian, X.; Scallan, C.D.; Powell, S.; Keller, T.; McMurray, M.; Labelle, A.; et al. Multiyear therapeutic benefit of AAV serotypes 2, 6, and 8 delivering factor VIII to hemophilia A mice and dogs. Blood 2006, 108, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hori, T.; Fukutome, M.; Maejima, C.; Matsushima, H.; Kobayashi, K.; Kitazawa, S.; Kitahara, R.; Kitano, K.; Kobayashi, K.; Moritoh, S.; et al. Gene delivery to cone photoreceptors by subretinal injection of rAAV2/6 in the mouse retina. Biochem. Biophys. Res. Commun. 2019, 515, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhu, T.; Qiao, C.; Zhou, L.; Wang, B.; Zhang, J.; Chen, C.; Li, J.; Xiao, X. Adeno-associated virus serotype 8 efficiently delivers genes to muscle and heart. Nat. Biotechnol. 2005, 23, 321–328. [Google Scholar] [CrossRef]
- Broekman, M.L.D.; Comer, L.A.; Hyman, B.T.; Sena-Esteves, M. Adeno-associated virus vectors serotyped with AAV8 capsid are more efficient than AAV-1 or -2 serotypes for widespread gene delivery to the neonatal mouse brain. Neuroscience 2006, 138, 501–510. [Google Scholar] [CrossRef]
- Sarkar, R.; Mucci, M.; Addya, S.; Tetreault, R.; Bellinger, D.A.; Nichols, T.C.; Kazazian, H.H. Long-term efficacy of adeno-associated virus serotypes 8 and 9 in hemophilia a dogs and mice. Hum. Gene Ther. 2006, 17, 427–439. [Google Scholar] [CrossRef]
- Pacak, C.A.; Mah, C.S.; Thattaliyath, B.D.; Conlon, T.J.; Lewis, M.A.; Cloutier, D.E.; Zolotukhin, I.; Tarantal, A.F.; Byrne, B.J. Recombinant adeno-associated virus serotype 9 leads to preferential cardiac transduction in vivo. Circ. Res. 2006, 99. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, K.; Fuess, S.; Storm, T.A.; Gibson, G.A.; Mctiernan, C.F.; Kay, M.A.; Nakai, H. Robust systemic transduction with AAV9 vectors in mice: Efficient global cardiac gene transfer superior to that of AAV8. Mol. Ther. 2006, 14, 45–53. [Google Scholar] [CrossRef]
- Duque, S.; Joussemet, B.; Riviere, C.; Marais, T.; Dubreil, L.; Douar, A.M.; Fyfe, J.; Moullier, P.; Colle, M.A.; Barkats, M. Intravenous administration of self-complementary AAV9 enables transgene delivery to adult motor neurons. Mol. Ther. 2009, 17, 1187–1196. [Google Scholar] [CrossRef]
- Foust, K.D.; Nurre, E.; Montgomery, C.L.; Hernandez, A.; Chan, C.M.; Kaspar, B.K. Intravascular AAV9 preferentially targets neonatal neurons and adult astrocytes. Nat. Biotechnol. 2009, 27, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.Q.; Ma, B.F.; Zhao, L.R.; Tatom, J.B.; Li, B.; Jiang, L.X.; Klein, R.L.; Duan, W.M. AAV9-mediated erythropoietin gene delivery into the brain protects nigral dopaminergic neurons in a rat model of Parkinson’s disease. Gene Ther. 2010, 17, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, T.; John Lye, R.; French, B.A.; Annex, B.H. Neuraminidase-mediated desialylation augments AAV9-mediated gene expression in skeletal muscle. J. Gene Med. 2018, 20, e3049. [Google Scholar] [CrossRef] [PubMed]
- Mori, S.; Wang, L.; Takeuchi, T.; Kanda, T. Two novel adeno-associated viruses from cynomolgus monkey: Pseudotyping characterization of capsid protein. Virology 2004, 330, 375–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, M.; Voutetakis, A.; Afione, S.; Zheng, C.; Mandikian, D.; Chiorini, J.A. Adeno-Associated Virus Type 12 (AAV12): A novel AAV serotype with sialic acid- and heparan sulfate proteoglycan-independent transduction activity. J. Virol. 2008, 82, 1399–1406. [Google Scholar] [CrossRef] [Green Version]
- Weitzman, M.D.; Linden, R.M. Adeno-Associated Virus Biology. Methods Mol. Biol. 2011, 807. [Google Scholar] [CrossRef]
- Gigout, L.; Rebollo, P.; Clement, N.; Warrington, K.H.; Muzyczka, N.; Linden, R.M.; Weber, T. Altering AAV tropism with mosaic viral capsids. Mol. Ther. 2005, 11, 856–865. [Google Scholar] [CrossRef]
- Zhou, X.; Muzyczka, N. In vitro packaging of adeno-associated virus DNA. J. Virol. 1998, 72, 3241–3247. [Google Scholar] [CrossRef] [Green Version]
- Wilmott, P.; Lisowski, L.; Alexander, I.E.; Logan, G.J. A user’s guide to the inverted terminal repeats of adeno-associated virus. Hum. Gene Ther. Methods 2019, 30, 206–213. [Google Scholar] [CrossRef]
- Weitzman, M.D.; Kyöstiö, S.R.M.; Kotin, R.M.; Owens, R.A. Adeno-associated virus (AAV) Rep proteins mediate complex formation between AAV DNA and its integration site in human DNA. Proc. Natl. Acad. Sci. USA 1994, 91, 5808–5812. [Google Scholar] [CrossRef] [Green Version]
- Berthet, C.; Raj, K.; Saudan, P.; Beard, P. How adeno-associated virus Rep78 protein arrests cells completely in S phase. Proc. Natl. Acad. Sci. USA 2005, 102, 13634–13639. [Google Scholar] [CrossRef] [Green Version]
- Zarate-Perez, F.; Mansilla-Soto, J.; Bardelli, M.; Burgner, J.W.; Villamil-Jarauta, M.; Kekilli, D.; Samso, M.; Linden, R.M.; Escalante, C.R. Oligomeric properties of adeno-associated virus rep68 reflect its multifunctionality. J. Virol. 2013, 87, 1232–1241. [Google Scholar] [CrossRef] [Green Version]
- Van Vliet, K.M.; Blouin, V.; Brument, N.; Agbandje-McKenna, M.; Snyder, R.O. The role of the adeno-associated virus capsid in gene transfer. Methods Mol. Biol. 2008, 437, 51–91. [Google Scholar] [CrossRef] [PubMed]
- King, J.A.; Dubielzig, R.; Grimm, D.; Kleinschmidt, J.A. DNA helicase-mediated packaging of adeno-associated virus type 2 genomes into preformed capsids. EMBO J. 2001, 20, 3282–3291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collaco, R.F.; Kalman-Maltese, V.; Smith, A.D.; Dignam, J.D.; Trempe, J.P. A Biochemical characterization of the adeno-associated virus rep40 helicase. J. Biol. Chem. 2003, 278, 34011–34017. [Google Scholar] [CrossRef] [Green Version]
- Trempe, J.P.; Carter, B.J. Alternate mRNA splicing is required for synthesis of adeno-associated virus VP1 capsid protein. J. Virol. 1988, 62, 3356–3363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becerra, S.P.; Rose, J.A.; Hardy, M.; Baroudy, B.M.; Anderson, C.W. Direct mapping of adeno-associated virus capsid proteins B and C: A possible ACG initiation codon. Proc. Natl. Acad. Sci. USA 1985, 82, 7919–7923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muralidhar, S.; Becerra, S.P.; Rose, J.A. Site-directed mutagenesis of adeno-associated virus type 2 structural protein initiation codons: Effects on regulation of synthesis and biological activity. J. Virol. 1994, 68, 170–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warrington, K.H.; Gorbatyuk, O.S.; Harrison, J.K.; Opie, S.R.; Zolotukhin, S.; Muzyczka, N. Adeno-associated virus type 2 VP2 capsid protein is nonessential and can tolerate large peptide insertions at its N terminus. J. Virol. 2004, 78, 6595–6609. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.; Bu, W.; Bhatia, S.; Hare, J.; Somasundaram, T.; Azzi, A.; Chapman, M.S. The atomic structure of adeno-associated virus (AAV-2), a vector for human gene therapy. Proc. Natl. Acad. Sci. USA 2002, 99, 10405–10410. [Google Scholar] [CrossRef] [Green Version]
- Girod, A.; Wobus, C.E.; Zádori, Z.; Ried, M.; Leike, K.; Tijssen, P.; Kleinschmidt, J.A.; Hallek, M. The VP1 capsid protein of adeno-associated virus type 2 is carrying a phospholipase A2 domain required for virus infectivity. J. Gen. Virol. 2002, 83, 973–978. [Google Scholar] [CrossRef] [Green Version]
- Sonntag, F.; Schmidt, K.; Kleinschmidt, J.A. A viral assembly factor promotes AAV2 capsid formation in the nucleolus. Proc. Natl. Acad. Sci. USA 2010, 107, 10220–10225. [Google Scholar] [CrossRef] [Green Version]
- Gao, G.; Vandenberghe, L.H.; Alvira, M.R.; Lu, Y.; Calcedo, R.; Zhou, X.; Wilson, J.M. Clades of adeno-associated viruses are widely disseminated in human tissues. J. Virol. 2004, 78, 6381–6388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, G.-P.; Alvira, M.R.; Wang, L.; Calcedo, R.; Johnston, J.; Wilson, J.M. Novel adeno-associated viruses from rhesus monkeys for human gene therapy. Proc. Natl. Acad. Sci. USA 2002, 99, 11854–11859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, A.M.; Nicolson, S.C.; Warischalk, J.K.; Samulski, R.J. AAV’s anatomy: Roadmap for optimizing vectors for translational success. Curr. Gene Ther. 2010, 10, 319–340. [Google Scholar] [CrossRef] [PubMed]
- Böttcher, B.; Kronenberg, S.; Kleinschmidt, J. Electron cryo-microscopy and image reconstruction of adeno-associated virus type 2 empty capsids. Microsc. Microanal. 2002, 8, 212–213. [Google Scholar] [CrossRef] [Green Version]
- Rabinowitz, J.E.; Rolling, F.; Li, C.; Conrath, H.; Xiao, W.; Xiao, X.; Samulski, R.J. Cross-packaging of a single adeno-associated virus (AAV) type 2 vector genome into multiple AAV Serotypes enables transduction with broad specificity. J. Virol. 2002, 76, 791–801. [Google Scholar] [CrossRef] [Green Version]
- Grimm, D.; Pandey, K.; Nakai, H.; Storm, T.A.; Kay, M.A. Liver transduction with recombinant adeno-associated virus is primarily restricted by capsid serotype not vector genotype. J. Virol. 2006, 80, 426–439. [Google Scholar] [CrossRef] [Green Version]
- Summerford, C.; Samulski, R.J. Membrane-associated heparan sulfate proteoglycan is a receptor for adeno-associated virus type 2 virions. J. Virol. 1998, 72, 1438–1445. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-L.; Jensen, R.L.; Schnepp, B.C.; Connell, M.J.; Shell, R.; Sferra, T.J.; Bartlett, J.S.; Clark, K.R.; Johnson, P.R. Molecular characterization of adeno-associated viruses infecting children. J. Virol. 2005, 79, 14781–14792. [Google Scholar] [CrossRef] [Green Version]
- Pillay, S.; Carette, J.E. Host determinants of adeno-associated viral vector entry. Curr. Opin. Virol. 2017, 24, 124–131. [Google Scholar] [CrossRef]
- Herrmann, A.K.; Grimm, D. High-throughput dissection of AAV–host interactions: The fast and the curious. J. Mol. Biol. 2018, 430, 2626–2640. [Google Scholar] [CrossRef] [PubMed]
- Zengel, J.; Carette, J.E. Structural and Cellular Biology of Adeno-Associated Virus Attachment and Entry, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 106, ISBN 9780128207543. [Google Scholar]
- Yang, Q.; Mamounas, M.; Yu, G.; Kennedy, S.; Leaker, B.; Mersn, J.; Yu, F.W.M.A.N.G.; Barber, J.R.; Al, Y.E.T. Development of novel cell surface CD34-Targeted recombinant adenoassociated virus vectors for gene therapy. Hum. Gene Ther. 1998, 1937, 1929–1937. [Google Scholar] [CrossRef]
- Wu, P.; Xiao, W.; Conlon, T.; Hughes, J.; Agbandje-McKenna, M.; Ferkol, T.; Flotte, T.; Muzyczka, N. Mutational analysis of the adeno-associated virus type 2 (AAV2) capsid gene and construction of AAV2 vectors with altered tropism. J. Virol. 2000, 74, 8635–8647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girod, A.; Ried, M.; Wobus, C.; Lahm, H.; Leike, K.; Kleinschmidt, J.; Deléage, G.; Hallek, M. Genetic capsid modifications allow efficient re-targeting of adeno- associated virus type 2. Nat. Med. 1999, 5, 1052–1056. [Google Scholar] [CrossRef] [PubMed]
- Nicklin, S.A.; Buening, H.; Dishart, K.L.; de Alwis, M.; Girod, A.; Hacker, U.; Thrasher, A.J.; Ali, R.R.; Hallek, M.; Baker, A.H. Efficient and selective AAV2-mediated gene transfer directed to human vascular endothelial cells. Mol. Ther. 2001, 4, 174–181. [Google Scholar] [CrossRef]
- White, S.J.; Nicklin, S.A.; Büning, H.; Brosnan, M.J.; Leike, K.; Papadakis, E.D.; Hallek, M.; Baker, A.H. Targeted gene delivery to vascular tissue in vivo by tropism-modified adeno-associated virus vectors. Circulation 2004, 109, 513–519. [Google Scholar] [CrossRef]
- Work, L.M. Vascular bed-targeted in vivo gene delivery using tropism-modified adeno-associated viruses. Mol. Ther. 2006, 13, 1–31. [Google Scholar] [CrossRef]
- Grifman, M.; Trepel, M.; Speece, P.; Gilbert, L.B.; Arap, W.; Pasqualini, R.; Weitzman, M.D. Incorporation of tumor-targeting peptides into recombinant adeno-associated virus capsids. Mol. Ther. 2001, 3, 964–975. [Google Scholar] [CrossRef]
- Ried, M.U.; Girod, A.; Leike, K.; Büning, H.; Hallek, M. Adeno-associated virus capsids displaying immunoglobulin-binding domains permit antibody-mediated vector retargeting to specific cell surface receptors. J. Virol. 2002, 76, 4559–4566. [Google Scholar] [CrossRef] [Green Version]
- Müller, O.J.; Kaul, F.; Weitzman, M.D.; Pasqualini, R.; Arap, W.; Kleinschmidt, J.A.; Trepel, M. Random peptide libraries displayed on adeno-associated virus to select for targeted gene therapy vectors. Nat. Biotechnol. 2003, 21, 1040–1046. [Google Scholar] [CrossRef]
- Bowles, D.E.; McPhee, S.W.J.; Li, C.; Gray, S.J.; Samulski, J.J.; Camp, A.S.; Li, J.; Wang, B.; Monahan, P.E.; Rabinowitz, J.E.; et al. Phase 1 gene therapy for duchenne muscular dystrophy using a translational optimized AAV vector. Mol. Ther. 2012, 20, 443–455. [Google Scholar] [CrossRef] [Green Version]
- Boucas, J.; Lux, K.; Huber, A.; Schievenbusch, S.; von Freyend, M.J.; Perabo, L.; Quadt-Humme, S.; Odenthal, M.; Hallek, M.; Büning, H. Engineering adeno-associated virus serotype 2-based targeting vectors using a new insertion site-position 453-and single point mutations. J. Gene Med. 2009, 12, 1103–1113. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Liu, L.; Nass, S.; O’Riordan, C.; Pastor, E.; Zhang, X.K. Direct liquid chromatography/mass spectrometry analysis for complete characterization of recombinant adeno-associated virus capsid proteins. Hum. Gene Ther. Methods 2017, 28, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Frederick, A.; Sullivan, J.; Liu, L.; Adamowicz, M.; Lukason, M.; Raymer, J.; Luo, Z.; Jin, X.; Rao, K.N.; O’Riordan, C. Engineered capsids for efficient gene delivery to the retina and cornea. Hum. Gene Ther. 2020, 31, 756–774. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.S.; Shemorry, A.; Varshavsky, A. N-terminal acetylation of cellular proteins creates specific degradation signals. Science 2010, 327, 973–977. [Google Scholar] [CrossRef] [Green Version]
- Maheshri, N.; Koerber, J.T.; Kaspar, B.K.; Schaffer, D.V. Directed evolution of adeno-associated virus yields enhanced gene delivery vectors. Nat. Biotechnol. 2006, 24, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Giver, L.; Shao, Z.; Affholter, J.A.; Arnold, F.H. Molecular evolution by staggered extension process (StEP) in vitro recombination. Nat. Biotechnol. 1998, 16, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, D.V.; Maheshri, N. Directed evolution of AAV mutants for enhanced gene delivery. Annu. Int. Conf. IEEE Eng. Med. Biol. Proc. 2004, 2, 3520–3523. [Google Scholar] [CrossRef]
- Pulicherla, N.; Shen, S.; Yadav, S.; Debbink, K.; Govindasamy, L.; Agbandje-Mckenna, M.; Asokan, A. Engineering liver-detargeted AAV9 vectors for cardiac and musculoskeletal gene transfer. Mol. Ther. 2011, 19, 1070–1078. [Google Scholar] [CrossRef] [Green Version]
- Marsic, D.; Govindasamy, L.; Currlin, S.; Markusic, D.M.; Tseng, Y.S.; Herzog, R.W.; Agbandje-Mckenna, M.; Zolotukhin, S. Vector design tour de force: Integrating combinatorial and rational approaches to derive novel adeno-associated virus variants. Mol. Ther. 2014, 22, 1900–1909. [Google Scholar] [CrossRef] [Green Version]
- Voigt, C.A.; Martinez, C.; Wang, Z.G.; Mayo, S.L.; Arnold, F.H. Protein building blocks preserved by recombination. Nat. Struct. Biol. 2002, 9, 553–558. [Google Scholar] [CrossRef]
- Ojala, D.S.; Sun, S.; Santiago-Ortiz, J.L.; Shapiro, M.G.; Romero, P.A.; Schaffer, D.V. In vivo selection of a computationally designed SCHEMA AAV library yields a novel variant for infection of adult neural stem cells in the SVZ. Mol. Ther. 2018, 26, 304–319. [Google Scholar] [CrossRef] [Green Version]
- Endelman, J.B.; Silberg, J.J.; Wang, Z.G.; Arnold, F.H. Site-directed protein recombination as a shortest-path problem. Protein Eng. Des. Sel. 2004, 17, 589–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deverman, B.E.; Pravdo, P.L.; Simpson, B.P.; Kumar, S.R.; Chan, K.Y.; Banerjee, A.; Wu, W.L.; Yang, B.; Huber, N.; Pasca, S.P.; et al. Cre-dependent selection yields AAV variants for widespread gene transfer to the adult brain. Nat. Biotechnol. 2016, 34, 204–209. [Google Scholar] [CrossRef]
- Chan, K.Y.; Jang, M.J.; Yoo, B.B.; Greenbaum, A.; Ravi, N.; Wu, W.L.; Sánchez-Guardado, L.; Lois, C.; Mazmanian, S.K.; Deverman, B.E.; et al. Engineered AAVs for efficient noninvasive gene delivery to the central and peripheral nervous systems. Nat. Neurosci. 2017, 20, 1172–1179. [Google Scholar] [CrossRef]
- Kumar, S.R.; Miles, T.F.; Chen, X.; Brown, D.; Dobreva, T.; Huang, Q.; Ding, X.; Luo, Y.; Einarsson, P.H.; Greenbaum, A.; et al. Multiplexed Cre-dependent selection yields systemic AAVs for targeting distinct brain cell types. Nat. Methods 2020, 17, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Ortiz, J.; Ojala, D.S.; Westesson, O.; Weinstein, J.R.; Wong, S.Y.; Steinsapir, A.; Kumar, S.; Holmes, I.; Schaffer, D.V. AAV ancestral reconstruction library enables selection of broadly infectious viral variants. Gene Ther. 2015, 22, 934–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zinn, E.; Pacouret, S.; Khaychuk, V.; Turunen, H.T.; Carvalho, L.S.; Andres-Mateos, E.; Shah, S.; Shelke, R.; Maurer, A.C.; Maurer, E.; et al. In silico reconstruction of the viral evolutionary lineage yields a potent gene therapy vector. Cell Rep. 2015, 12, 1056–1068. [Google Scholar] [CrossRef] [Green Version]
- Pan, B.; Askew, C.; Galvin, A.; Heman-Ackah, S.; Asai, Y.; Indzhykulian, A.A.; Jodelka, F.M.; Hastings, M.L.; Lentz, J.J.; Vandenberghe, L.H.; et al. Gene therapy restores auditory and vestibular function in a mouse model of Usher syndrome type 1c. Nat. Biotechnol. 2017, 35, 264–272. [Google Scholar] [CrossRef]
- Ponnazhagan, S.; Mahendra, G.; Kumar, S.; Thompson, J.A.; Castillas, M. Conjugate-based targeting of recombinant adeno-associated virus type 2 vectors by using avidin-linked ligands. J. Virol. 2002, 76, 12900–12907. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Fang, Y.; Zhou, Y.; Zandi, E.; Lee, C.L.; Joo, K.I.; Wang, P. Site-specific modification of adeno-associated viruses via a genetically engineered aldehyde tag. Small 2013, 9, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Kelemen, R.E.; Mukherjee, R.; Cao, X.; Erickson, S.B.; Zheng, Y.; Chatterjee, A. A precise chemical strategy to alter the receptor specificity of the adeno-associated virus. Angew. Chem. Int. Ed. 2016, 55, 10645–10649. [Google Scholar] [CrossRef] [PubMed]
- Zdechlik, A.C.; He, Y.; Aird, E.J.; Gordon, W.R.; Schmidt, D. Programmable assembly of adeno-associated virus-antibody composites for receptor-mediated gene delivery. Bioconjug. Chem. 2020, 31, 1093–1106. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, C.; Adjali, O.; Mingozzi, F. Unraveling the complex story of immune responses to AAV vectors trial after trial. Hum. Gene Ther. 2017, 28, 1061–1074. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serotype | Origin | Primary Receptor | Secondary Receptor | Natural Tropism | Selected Ongoing Clinical Trials | Links |
|---|---|---|---|---|---|---|
| AAV1 | Non-human primate | Sialic acid | AAV receptor (AAVR) | Muscle, CNS, heart, liver, lungs | No trials underway | [10,11,12,13] |
| AAV2 | Human | Heparan sulfate proteoglycan (HSPG) | Integrin, fibroblast growth factor receptor (FGFR), hepatocyte growth factor receptor (HGFR), laminin receptor (LamR) | Heart, CNS, liver, lungs, retina | Pompe disease (NCT03533673), Parkinson’s disease (NCT01621581), hemophilia (NCT03489291) | [14,15,16,17,18,19,20,21] |
| AAV3 | Non-human primate | HSPG | LamR, FGFR, HGFR, AAVR | Liver | No trials underway | [22,23] |
| AAV4 | Non-human primate | Sialic acid | Unknown | Retina, lungs, kidney | No trials underway | [24,25] |
| AAV5 | Human | Sialic acid | Platelet-derived growth factor receptor (PDGFR), AAVR | Retina, CNS, liver | Hemophilia (NCT03520712) | [13,16,26,27,28,29,30] |
| AAV6 | Human | HSPG, sialic acid | EGFR, AAVR | Heart, liver, muscle, retina | Hemophilia (NCT03061201) mucopolysaccharidosis type I (NCT02702115) | [31,32,33] |
| AAV7 | Non-human primate | Unknown | Unknown | Liver | [25] | |
| AAV8 | Non-human primate | Unknown | LamR, AAVR | Muscle, heart, CNS, liver | Eye disease (NCT03066258), hemophilia (NCT00979238), myopathy (NCT03199469) | [21,30,34,35,36] |
| AAV9 | Human | Galactose | LamR, AAVR | Heart, CNS, liver | Muscle diseases (NCT03362502), Pompe disease (NCT02240407), Danon disease (NCT03489291) | [25,36,37,38,39,40,41,42] |
| AAV10 | Non-human primate | Unknown | Unknown | Muscle, myoblast tissue | No trials underway | [43] |
| AAV11 | Non-human primate | Unknown | Unknown | Muscle, myoblast tissue | No trials underway | [43] |
| AAV12 | Non-human primate | Unknown | Unknown | Salivary glands, muscle | No trials underway | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korneyenkov, M.A.; Zamyatnin, A.A., Jr. Next Step in Gene Delivery: Modern Approaches and Further Perspectives of AAV Tropism Modification. Pharmaceutics 2021, 13, 750. https://doi.org/10.3390/pharmaceutics13050750
Korneyenkov MA, Zamyatnin AA Jr. Next Step in Gene Delivery: Modern Approaches and Further Perspectives of AAV Tropism Modification. Pharmaceutics. 2021; 13(5):750. https://doi.org/10.3390/pharmaceutics13050750
Chicago/Turabian StyleKorneyenkov, Maxim A., and Andrey A. Zamyatnin, Jr. 2021. "Next Step in Gene Delivery: Modern Approaches and Further Perspectives of AAV Tropism Modification" Pharmaceutics 13, no. 5: 750. https://doi.org/10.3390/pharmaceutics13050750
APA StyleKorneyenkov, M. A., & Zamyatnin, A. A., Jr. (2021). Next Step in Gene Delivery: Modern Approaches and Further Perspectives of AAV Tropism Modification. Pharmaceutics, 13(5), 750. https://doi.org/10.3390/pharmaceutics13050750