
Selective Moonlighting Cell-Penetrating Peptides

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Machine Learning
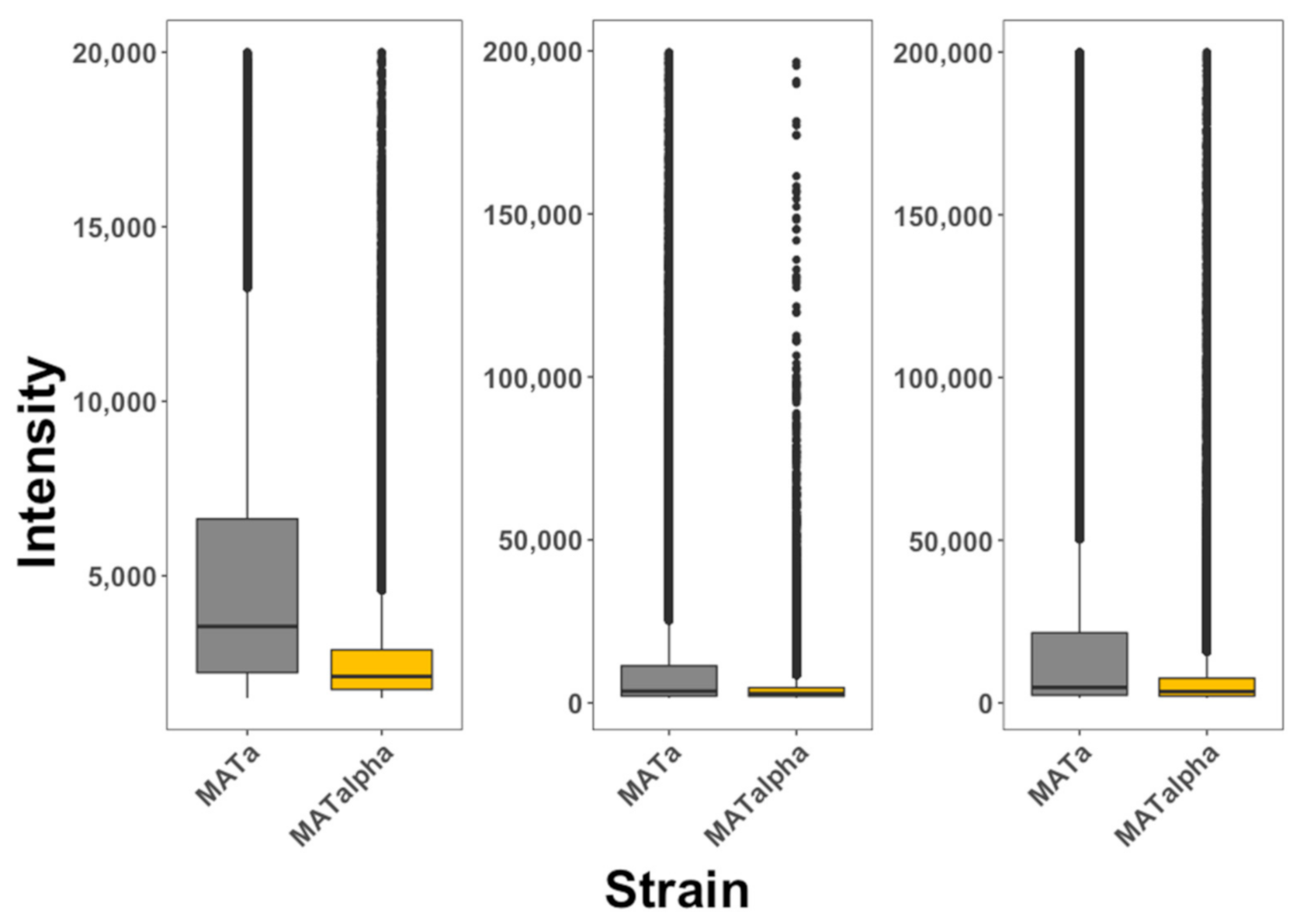
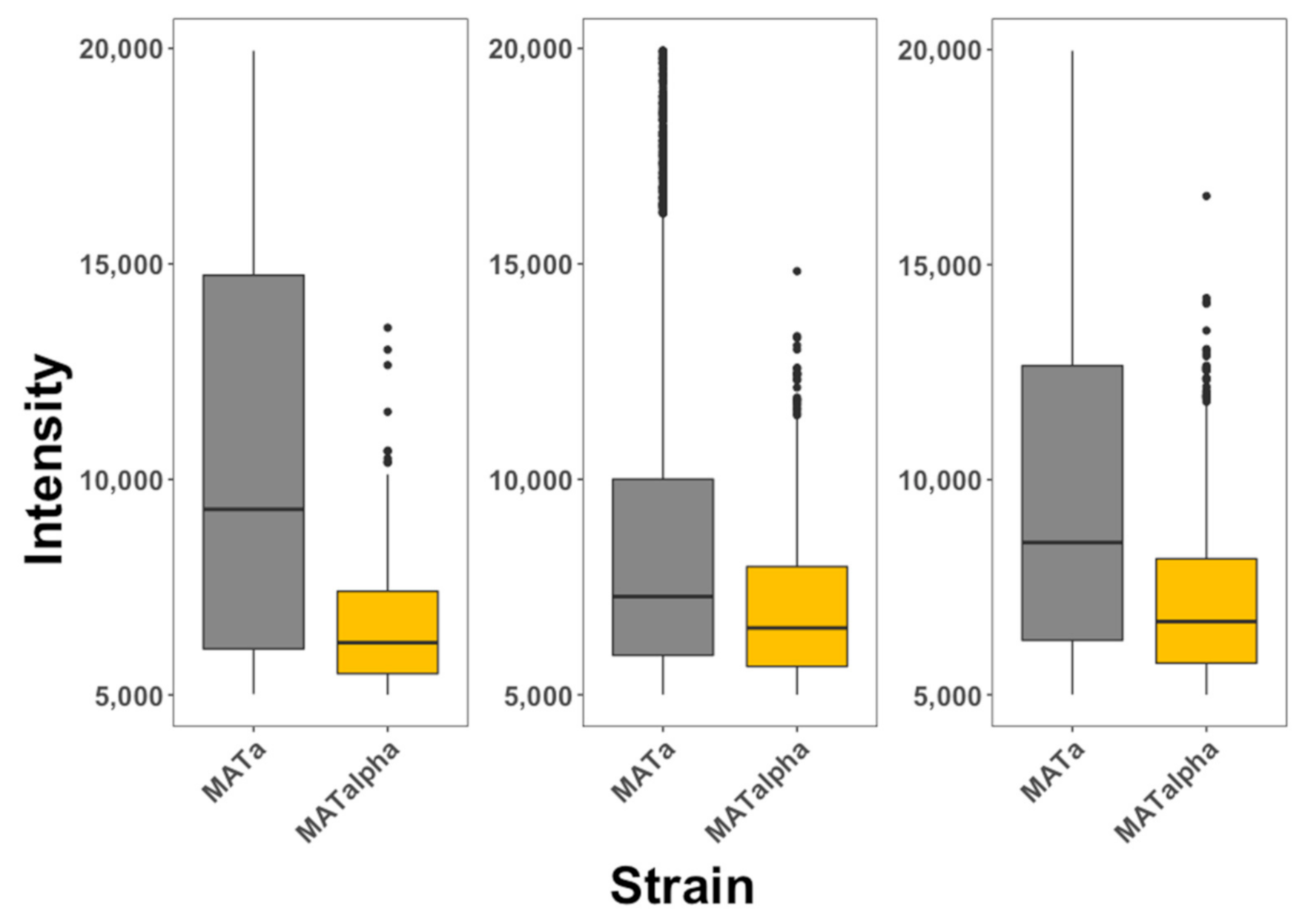
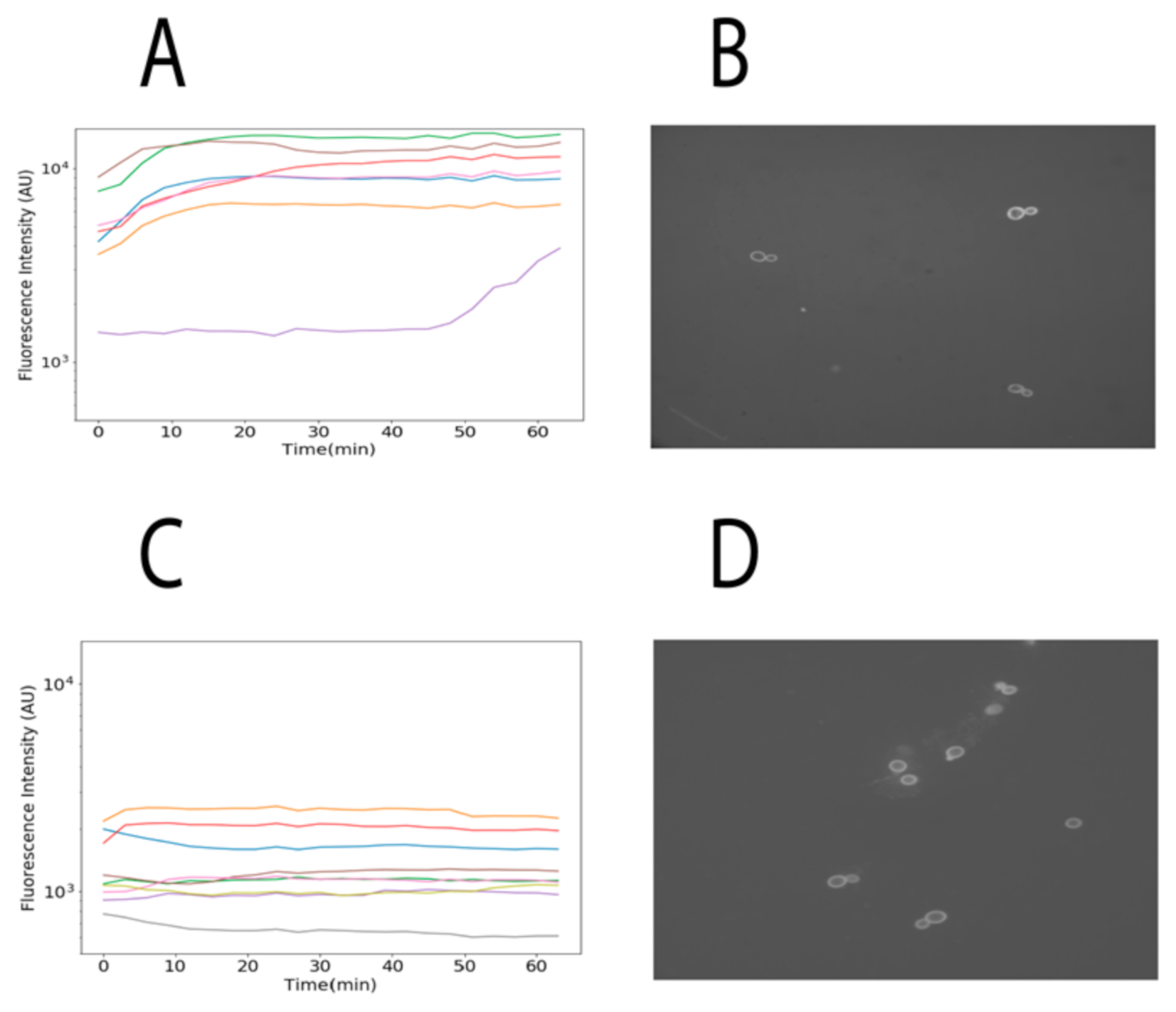
2.3. Quantifying the Internalization in Saccharomyces cerevisiae Cells
2.4. Quantifying the Internalization in HEK293T, HEK293T-NEP and HeLa Cells
2.4.1. Cell Culture
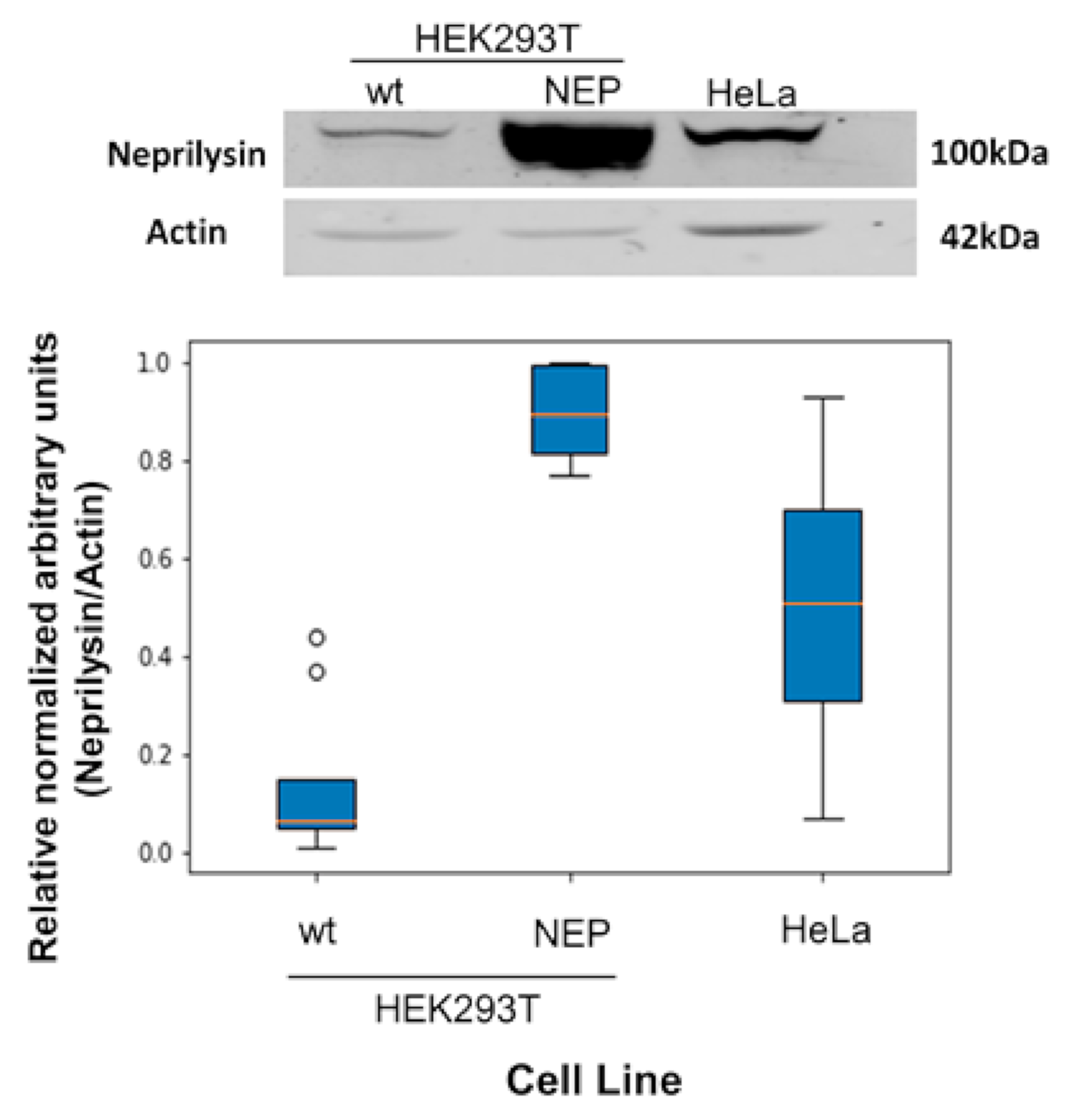
2.4.2. HEK293T-NEP Stable Transfection
2.4.3. Western-Blot
2.4.4. Internalization Assays
2.4.5. Cytotoxicity Assays
2.4.6. Data Analysis
3. Results
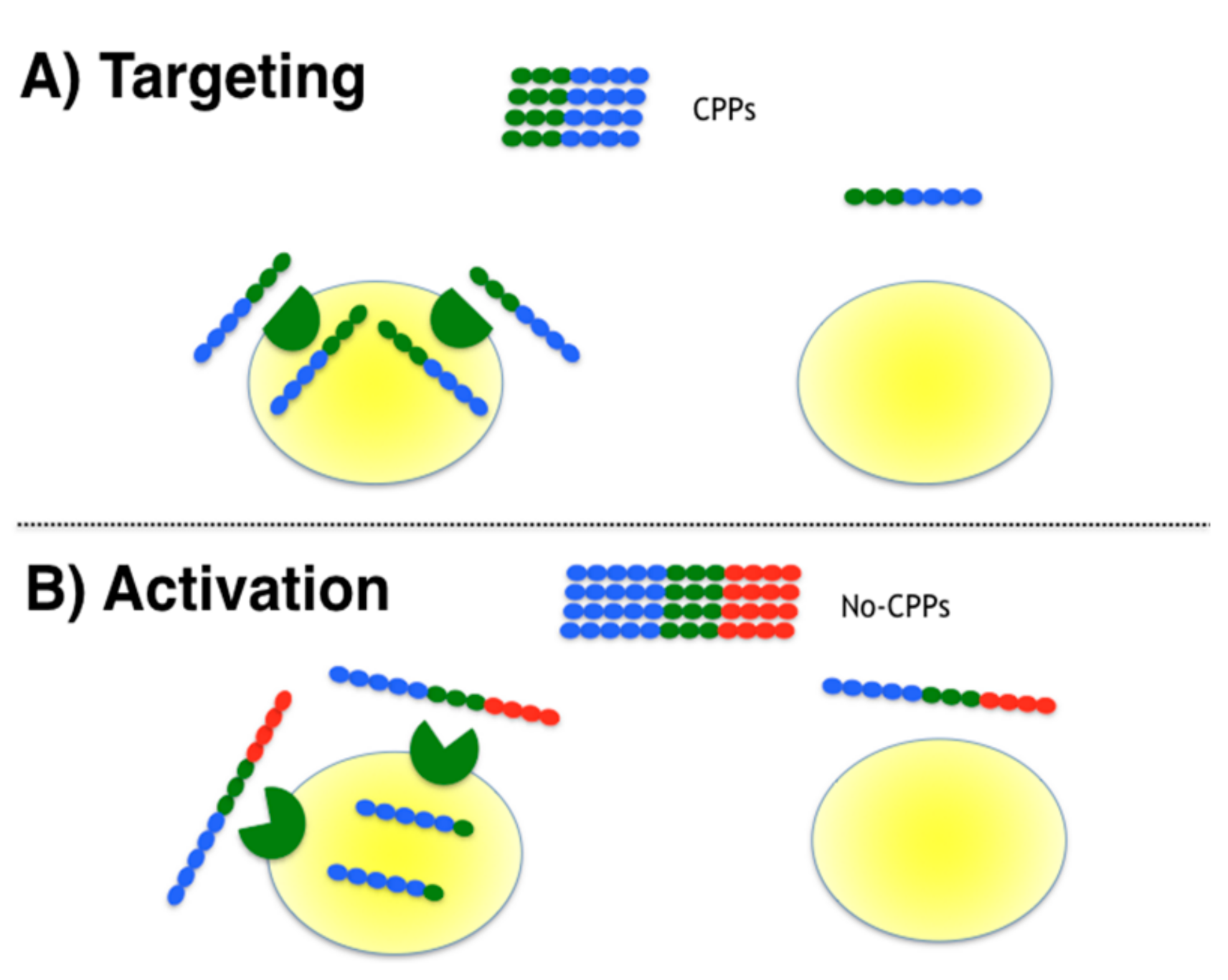
3.1. Designing Moonlighting CPPs
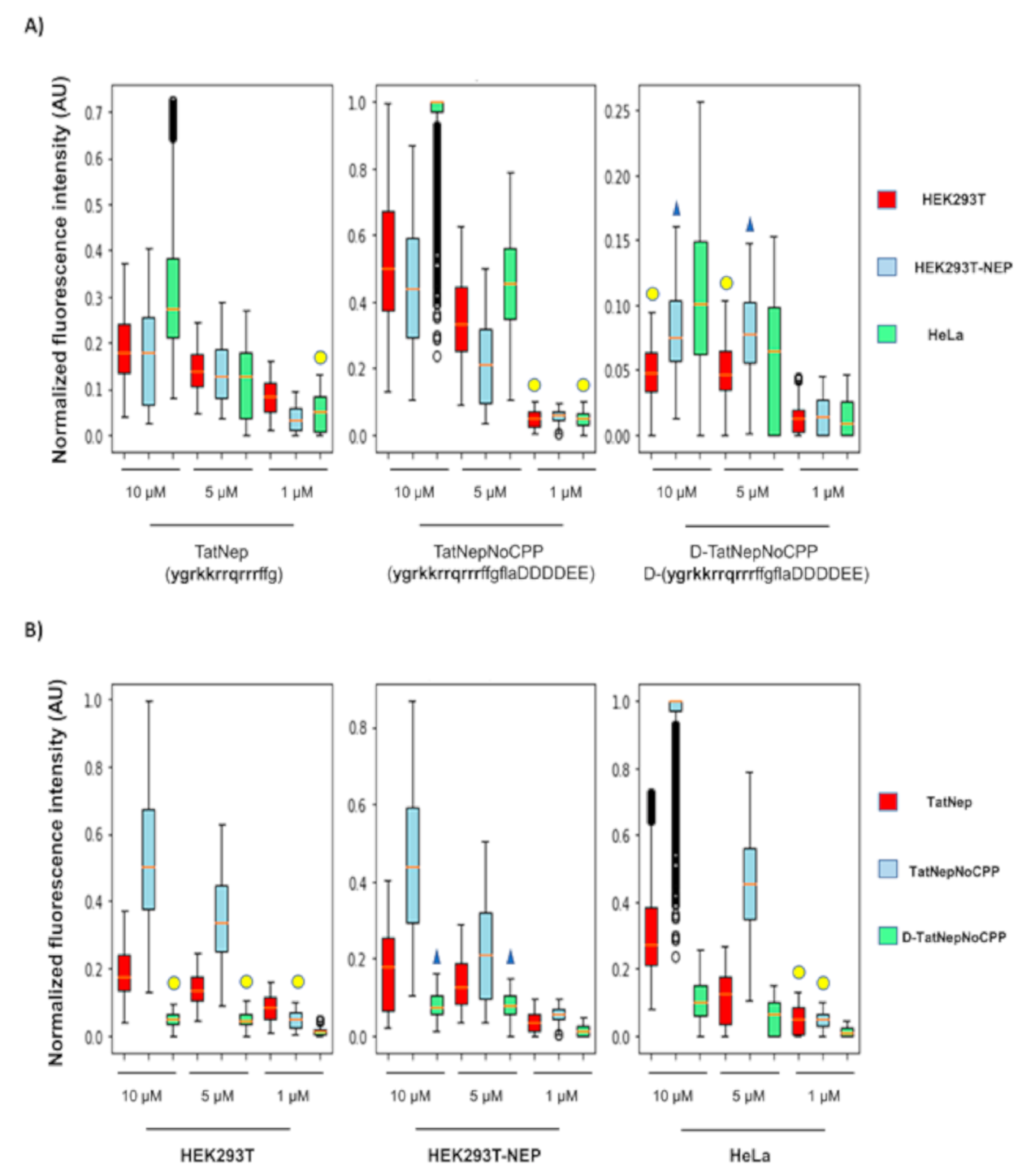
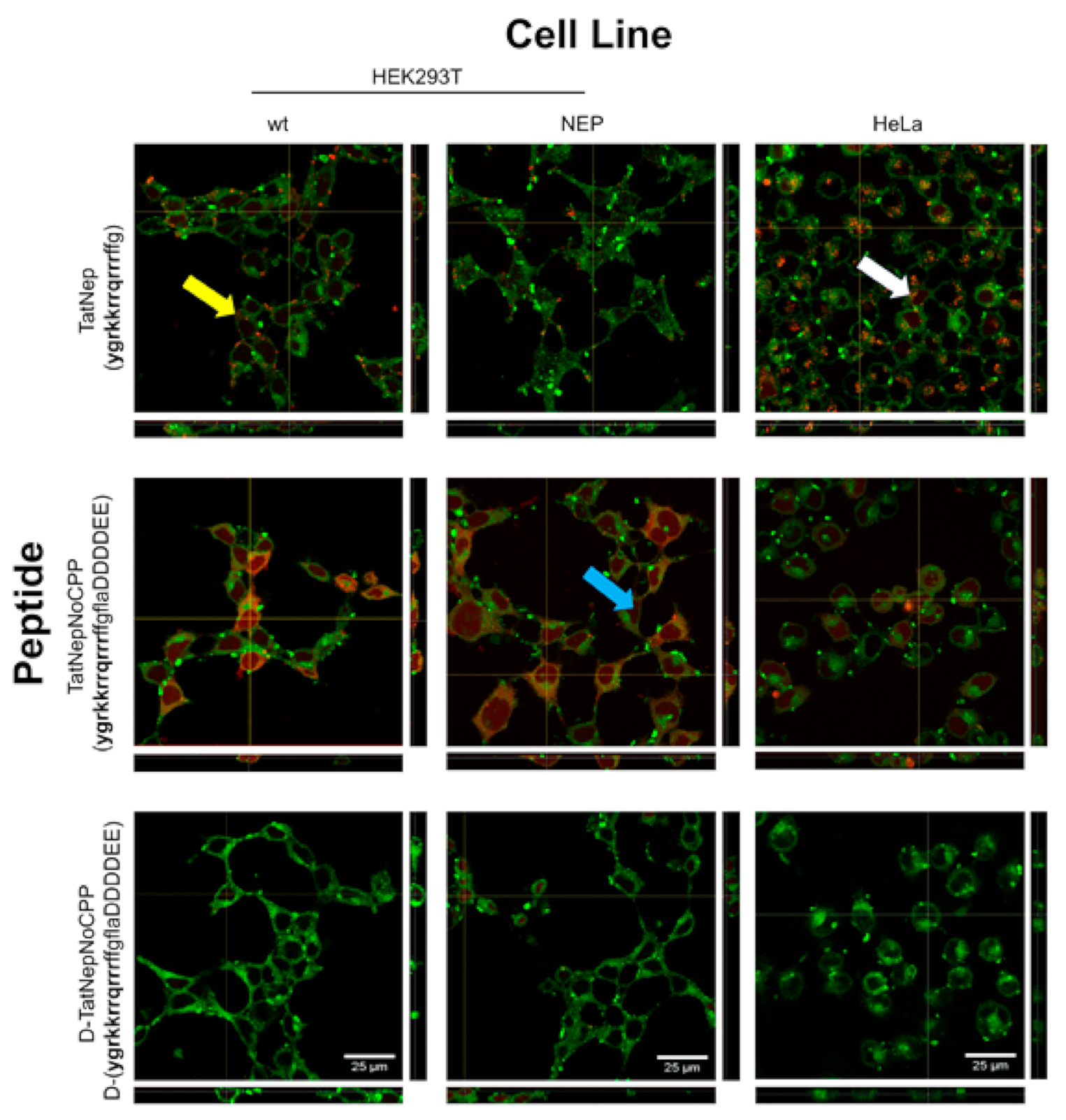
3.2. Experimental Testing of Targeted Moonlighting CPPs
3.3. Experimental Testing of Activatable Moonlighting CPPs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Richard, J.-P.; Melikov, K.; Vives, E.; Ramos, C.; Verbeure, B.; Gait, M.J.; Chernomordik, L.V.; Lebleu, B. Cell-penetrating Peptides. A reevaluation of the mechanism of cellular uptake. J. Biol. Chem. 2003, 278, 585–590. [Google Scholar] [CrossRef] [Green Version]
- Del Rio, G.; Klipp, E.; Herrmann, A. Using Confocal Microscopy and Computational Modeling to Investigate the Cell-Penetrating Properties of Antimicrobial Peptides. In Antimicrobial Peptides. Methods in Molecular Biology; Hansen, P., Ed.; Humana Press: New York, NY, USA, 2017; Volume 1548. [Google Scholar] [CrossRef]
- Gestin, M.; Dowaidar, M.; Langel, Ü. Uptake Mechanism of Cell-Penetrating Peptides. In Peptides and Peptide-Based Biomaterials And their Biomedical Applications and Their Biomedical Applications; Sunna, A., Care, A., Bergquist, P., Eds.; Springer: Cham, Switzerland, 2017; Volume 1030. [Google Scholar] [CrossRef]
- Gallo, M.; Defaus, S.; Andreu, D. 1988–2018: Thirty years of drug smuggling at the nano scale. Challenges and opportunities of cell-penetrating peptides in biomedical research. Arch. Biochem. Biophys. 2019, 661, 74–86. [Google Scholar]
- Méndez-Ardoy, A.; Lostalé-Seijo, I.; Montenegro, J. Where in the Cell Is our Cargo? Methods Currently Used To Study Intracellular Cytosolic Localisation. ChemBioChem 2019, 20, 488–498. [Google Scholar] [CrossRef] [Green Version]
- Araste, F.; Abnous, K.; Hashemi, M.; Taghdisi, S.M.; Ramezani, M.; Alibolandi, M. Peptide-based targeted therapeutics: Focus on cancer treatment. J. Control. Release 2018, 292, 141–162. [Google Scholar] [CrossRef] [PubMed]
- Guidotti, G.; Brambilla, L.; Rossi, D. Cell-Penetrating Peptides: From Basic Research to Clinics. Trends Pharmacol. Sci. 2017, 38, 406–424. [Google Scholar] [CrossRef] [PubMed]
- Bode, S.; Löwik, D. Constrained cell penetrating peptides. Drug Discov. Today: Technol. 2017, 26, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Reissmann, S. Cell penetration: Scope and limitations by the application of cell-penetrating peptides. J. Pept. Sci. 2014, 20, 760–784. [Google Scholar] [CrossRef]
- Ellerby, H.M.; Arap, W.; Ellerby, L.M.; Kain, R.; Andrusiak, R.; Del Rio, G.; Krajewski, S.; Lombardo, C.R.; Rao, R.; Ruoslahti, E.; et al. Anti-cancer activity of targeted pro-apoptotic peptides. Nat. Med. 1999, 5, 1032–1038. [Google Scholar] [CrossRef]
- Rodríguez Plaza, J.G.; Villalón Rojas, A.; Herrera, S.; Garza-Ramos, G.; Torres Larios, A.; Amero, C.; Zarraga Granados, G.; Gutiérrez Aguilar, M.; Lara Ortiz, M.T.; Polanco Gonzalez, C.; et al. Moonlighting Peptides with Emerging Function. PLoS ONE 2012, 7, e40125. [Google Scholar] [CrossRef] [Green Version]
- Diener, C.; Martínez, G.G.R.; Blas, D.M.; González, D.A.C.; Corzo, G.; Castro-Obregon, S.; Del Río, G. Effective Design of Multifunctional Peptides by Combining Compatible Functions. PLoS Comput. Biol. 2016, 12, e1004786. [Google Scholar] [CrossRef]
- Arellano, V.J.; Martinell García, P.; Rodríguez Plaza, J.G.; Lara Ortiz, M.T.; Schreiber, G.; Volkmer, R.; Klipp, E.; Rio, G.D. An Antimicrobial Peptide Induces FIG1-Dependent Cell Death During Cell Cycle Arrest in Yeast. Front. Microbiol. 2018, 9, 1240. [Google Scholar] [CrossRef]
- Zárraga-Granados, G.; Muciño-Hernández, G.; Sánchez-Carbente, M.R.; Villamizar-Gálvez, W.; Peñas-Rincón, A.; Arredondo, C.; Andrés, M.E.; Wood, C.; Covarrubias, L.; Castro-Obregón, S. The nuclear receptor NR4A1 is regulated by SUMO modification to induce autophagic cell death. PLoS ONE 2020, 15, e0222072. [Google Scholar] [CrossRef] [Green Version]
- Ni, D.; Xu, P.; Gallagher, S. Immunoblotting and immunodetection. Curr. Protoc. Mol. Biol. 2016, 114, 10.8.1–10.8.37. [Google Scholar] [CrossRef] [PubMed]
- Blumer, K.J.; Reneke, J.E.; Thorner, J. The STE2 gene product is the ligand-binding component of the alpha-factor receptor of Saccharomyces cerevisiae. J. Biol. Chem. 1988, 263, 10836–10842. [Google Scholar] [CrossRef]
- Dube, P.; Konopka, J.B. Identification of a polar region in transmembrane domain 6 that regulates the function of the G protein-coupled alpha-factor receptor. Mol. Cell. Biol. 1998, 18, 7205–7215. [Google Scholar] [CrossRef] [Green Version]
- Bajwa, A.A.; Khadim, M.T.; Din, H.U.; Ali, S.S.; Jamil, U.; Khan, U.A.S. Immunohistochemical Expression of CD10, BCL6 and MUM1 in Differentiating Diffuse Large B Cell Lymphoma Subtypes. J. Coll. Physicians Surg. Pak. 2017, 27, 621–624. [Google Scholar] [PubMed]
- Hersh, L.B.; Morihara, K. Comparison of the subsite specificity of the mammalian neutral endopeptidase 24.11 (enkephalinase) to the bacterial neutral endopeptidase thermolysin. J. Biol. Chem. 1986, 261, 6433–6437. [Google Scholar] [PubMed]
- Uhlen, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Kowarsch, A.; Fuchs, A.; Frishman, D.; Pagel, P. Correlated Mutations: A Hallmark of Phenotypic Amino Acid Substitutions. PLoS Comput. Biol. 2010, 6, e1000923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fodor, A.A.; Aldrich, R.W. Influence of conservation on calculations of amino acid covariance in multiple sequence alignments. Proteins Struct. Funct. Bioinform. 2004, 56, 211–221. [Google Scholar] [CrossRef]
- Göbel, U.; Sander, C.; Schneider, R.; Valencia, A. Correlated mutations and residue contacts in proteins. Proteins Struct. Funct. Genet. 1994, 18, 309–317. [Google Scholar] [CrossRef]
- Jiang, T.; Olson, E.S.; Nguyen, Q.T.; Roy, M.; Jennings, P.A.; Tsien, R.Y. Tumor imaging by means of proteolytic activation of cell-penetrating peptides. Proc. Natl. Acad. Sci. USA 2004, 101, 17867–17872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Shi, N.-Q.; Gao, W.; Xiang, B. Enhancing cellular uptake of activable cell-penetrating peptide-doxorubicin conjugate by enzymatic cleavage. Int. J. Nanomed. 2012, 7, 1613–1621. [Google Scholar] [CrossRef] [Green Version]
- Yoo, J.; Rejinold, N.S.; Lee, D.; Jon, S.; Kim, Y.-C. Protease-activatable cell-penetrating peptide possessing ROS-triggered phase transition for enhanced cancer therapy. J. Control. Release 2017, 264, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Plaza, J.G.R.; Morales-Nava, R.; Diener, C.; Schreiber, G.; Gonzalez, Z.D.; Ortiz, M.T.L.; Blake, I.O.; Pantoja, O.; Volkmer, R.; Klipp, E.; et al. Cell Penetrating Peptides and Cationic Antibacterial Peptides. J. Biol. Chem. 2014, 289, 14448–14457. [Google Scholar] [CrossRef] [Green Version]
- Oefner, C.; D’Arcy, A.; Hennig, M.; Winkler, F.K.; Dale, G.E. Structure of human neutral endopeptidase (neprilysin) complexed with phosphoramidon. J. Mol. Biol. 2000, 296, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Sato, B.; Katagiri, Y.U.; Iijima, K.; Yamada, H.; Ito, S.; Kawasaki, N.; Okita, H.; Fujimoto, J.; Kiyokawa, N. The human CD10 lacking an N-glycan at Asn628 is deficient in surface expression and neutral endopeptidase activity. Biochim. Biophys. Acta (BBA) Gen. Subj. 2012, 1820, 1715–1723. [Google Scholar] [CrossRef]
- Mueller, J.; Kretzschmar, I.; Volkmer, R.; Boisguerin, P. Comparison of cellular uptake using 22 CPPs in 4 different cell lines. Bioconjugate Chem. 2008, 19, 2363–2374. [Google Scholar] [CrossRef]
- Wallbrecher, R.; Ackels, T.; Olea, R.A.; Klein, M.J.; Caillon, L.; Schiller, J.; Bovée-Geurts, P.H.; van Kuppevelt, T.H.; Ulrich, A.S.; Spehr, M.; et al. Membrane permeation of arginine-rich cell-penetrating peptides independent of transmembrane potential as a function of lipid composition and membrane fluidity. J. Control. Release 2017, 256, 68–78. [Google Scholar] [CrossRef]
- Tyagi, M.; Rusnati, M.; Presta, M.; Giacca, M. Internalization of HIV-1 Tat Requires Cell Surface Heparan Sulfate Proteoglycans J. Biol. Chem. 2001, 276, 3254–3261. [Google Scholar] [CrossRef] [Green Version]
- Galvao, J.; Davis, B.; Tilley, M.; Normando, E.; Duchen, M.R.; Cordeiro, M.F. Unexpected low-dose toxicity of the universal solvent DMSO. FASEB J. 2014, 28, 1317–1330. [Google Scholar] [CrossRef]
- Rawlings, A.E. Membrane proteins: Always an insoluble problem? Biochem. Soc. Trans. 2016, 44, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Ellerby, H.M.; Lee, S.; Ellerby, L.M.; Chen, S.; Kiyota, T.; del Rio, G.; Sugihara, G.; Sun, Y.; Bredesen, D.E.; Arap, W.; et al. An artificially designed pore-forming protein with anti-tumor effects. J. Biol. Chem. 2003, 278, 35311–35316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jong, H.; Bonger, K.M.; Löwik, D.W.P.M. Activatable cell-penetrating peptides: 15 years of research. RSC Chem. Biol. 2020, 1, 192–203. [Google Scholar] [CrossRef]
- Barnhart, K.F.; Christianson, D.R.; Hanley, P.W.; Driessen, W.H.P.; Bernacky, B.J.; Baze, W.B.; Wen, S.; Tian, M.; Ma, J.; Kolonin, M.G.; et al. A peptidomimetic targeting white fat causes weight loss and improved insulin resistance in obese monkeys. Sci. Transl. Med. 2011, 3, 108ra112. [Google Scholar] [CrossRef] [Green Version]
- Diener, C.; Schreiber, G.; Giese, W.; Del Rio, G.; Schröder, A.; Klipp, E. Yeast mating and image-based quantification of spatial pattern formation. PLoS Comput. Biol. 2014, 10, e1003690. [Google Scholar] [CrossRef] [Green Version]
- Carrodus, N.L.; Teng, K.S.; Munro, K.M.; Kennedy, M.J.; Gunnersen, J.M. Differential labeling of cell-surface and internalized proteins after antibody feeding of live cultured neurons. J. Vis. Exp. 2014. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Sequence | P(CPP) |
|---|---|---|
| α-NLS-C | RLWHWLQLKPGQPMYWRQPKSKRKVRR | 0.91 |
| NLS-α-CE | FRKWRRKPKKKRKVWWRKVKRRWHWLQLKPGQPMY | 0.97 |
| Chimera | KRRWRFVWMNPKKKRKVPPWPYLLWWHWLQLKPGQPMY | 0.88 |
| TatNep | YGRKKRRQRRRFFG | 0.99 |
| TatNepNoCPP | YGRKKRRQRRRFFGFLADDDDEE | 0.112 |
| D-TatNepNoCPP | D-(YGRKKRRQRRRFFGFLADDDDEE) | 0.11 |
| Cell Line | Genotype | Provider |
|---|---|---|
| S. cerevisiae MATa | BY4741: his3D1; leu2D0; met15D0; ura3D0 | * |
| S. cerevisiae MATα | BY4742: his3D1; leu2D0; lys2D0; ura3D0 | Euroscarf |
| S. cerevisiae MATa: RPL9A-GFP | BY4741: his3D1; leu2D0; met15D0; ura3D0; RPL9A-GFP | * |
| S. cerevisiae MATa:bar1Δ | BY4741: his3D1; leu2D0; met15D0; ura3D0; bar1Δ | * |
| HEK293T | ND | # |
| HeLa | ND | # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morán-Torres, R.; Castillo González, D.A.; Durán-Pastén, M.L.; Aguilar-Maldonado, B.; Castro-Obregón, S.; Del Rio, G. Selective Moonlighting Cell-Penetrating Peptides. Pharmaceutics 2021, 13, 1119. https://doi.org/10.3390/pharmaceutics13081119
Morán-Torres R, Castillo González DA, Durán-Pastén ML, Aguilar-Maldonado B, Castro-Obregón S, Del Rio G. Selective Moonlighting Cell-Penetrating Peptides. Pharmaceutics. 2021; 13(8):1119. https://doi.org/10.3390/pharmaceutics13081119
Chicago/Turabian StyleMorán-Torres, Rafael, David A. Castillo González, Maria Luisa Durán-Pastén, Beatriz Aguilar-Maldonado, Susana Castro-Obregón, and Gabriel Del Rio. 2021. "Selective Moonlighting Cell-Penetrating Peptides" Pharmaceutics 13, no. 8: 1119. https://doi.org/10.3390/pharmaceutics13081119
APA StyleMorán-Torres, R., Castillo González, D. A., Durán-Pastén, M. L., Aguilar-Maldonado, B., Castro-Obregón, S., & Del Rio, G. (2021). Selective Moonlighting Cell-Penetrating Peptides. Pharmaceutics, 13(8), 1119. https://doi.org/10.3390/pharmaceutics13081119