
Prevalence of Mutations in the pfcoronin Gene and Association with Ex Vivo Susceptibility to Common Quinoline Drugs against Plasmodium falciparum

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
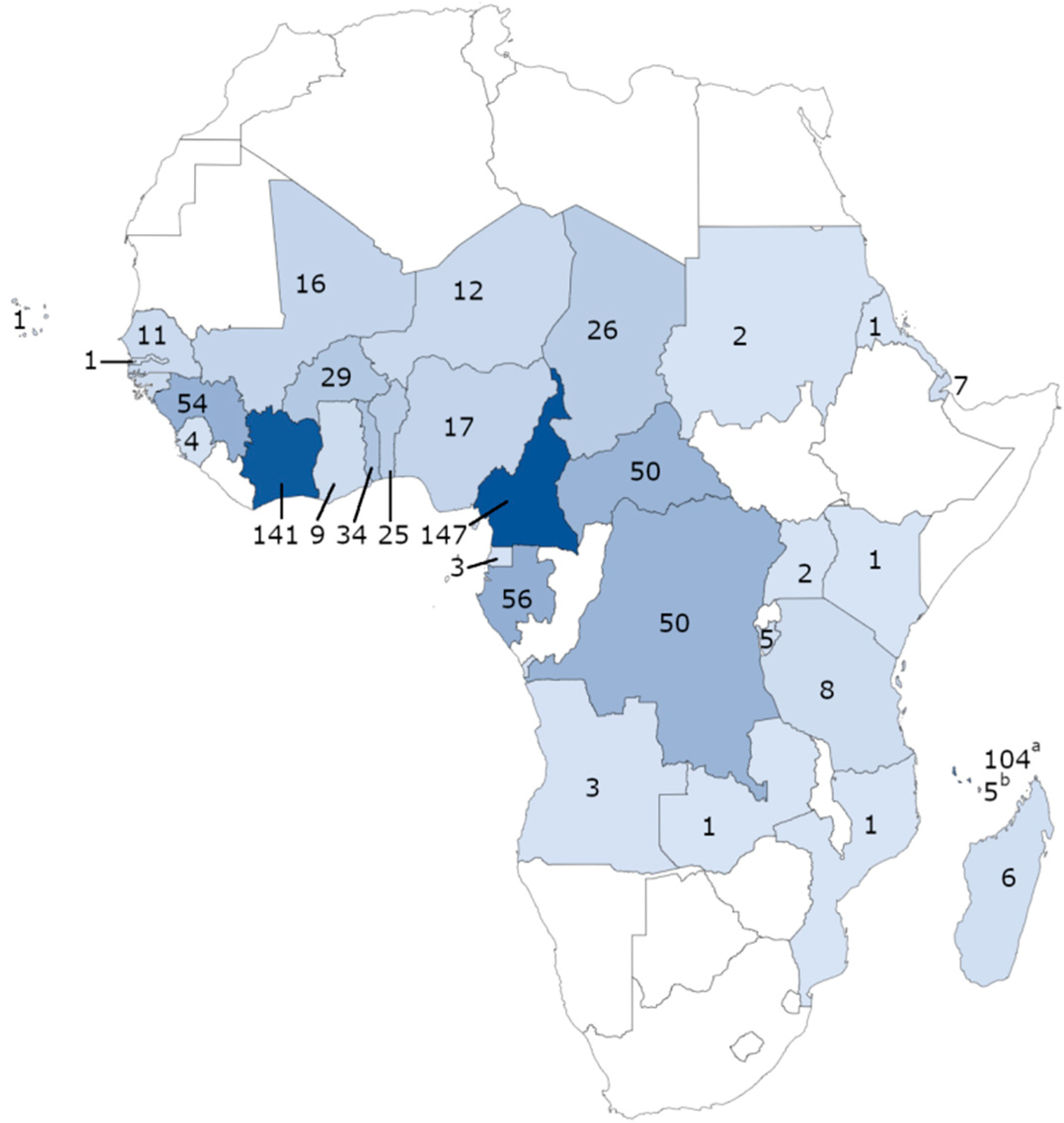
2.1. Sample Collection
2.2. Gene Sequencing
2.3. Ex Vivo Assay
2.4. Data Management and Statistical Analysis
3. Results
3.1. Analysis of pfcoronin Gene
3.2. Drug Susceptibility
3.3. Association between In Vitro Responses (IC50) and pfcoronin Mutations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guidelines for the Treatment of Malaria. Available online: https://apps.who.int/iris/handle/10665/162441 (accessed on 13 July 2021).
- World Malaria Report 2020. Available online: https://www.who.int/publications-detail-redirect/9789240015791 (accessed on 13 July 2021).
- World Malaria Report 2019. Available online: http://www.who.int/malaria/publications/world-malaria-report-2019/en/ (accessed on 13 July 2021).
- Kendjo, E.; Houzé, S.; Mouri, O.; Taieb, A.; Gay, F.; Jauréguiberry, S.; Tantaoui, I.; Ndour, P.A.; Buffet, P.; Piarroux, M.; et al. Epidemiologic trends in malaria incidence among travelers returning to metropolitan France, 1996-2016. JAMA Netw. Open 2019, 2, e191691. [Google Scholar] [CrossRef] [PubMed]
- Thellier, M.; Simard, F.; Musset, L.; Cot, M.; Velut, G.; Kendjo, E.; Pradines, B. Changes in malaria epidemiology in France and worldwide, 2000–2015. Med. Mal. Infect. 2020, 50, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Pradines, B.; Thellier, M.; Kendjo, E.; Houzé, S. Epidémiologie du paludisme d’importatation en France. Rev. Prat. 2019, 69, 150–152. [Google Scholar]
- Rapport d’activité 2020 du Centre National de Référence du Paludisme. Available online: https://cnr-paludisme.fr/activites-dexpertise/rapports-dactivites/ (accessed on 13 July 2021).
- Mbacham, F.W.; Ayong, L.; Guewo-Fokeng, M.; Makoge, V. Current situation of malaria in Africa. Methods Mol. Biol. 2019, 2013, 29–44. [Google Scholar]
- Dondorp, A.M.; Nosten, F.; Yi, P.; Das, D.; Phyo, A.P.; Tarning, J.; Lwin, K.M.; Ariey, F.; Hanpithakpong, W.; Lee, S.J.; et al. Artemisinin resistance in Plasmodium falciparum malaria. N. Engl. J. Med. 2009, 361, 455–467. [Google Scholar] [CrossRef] [Green Version]
- Phyo, A.P.; Nkhoma, S.; Stepniewska, K.; Ashley, E.A.; Nair, S.; McRready, R.; Moo, C.L.; Al-Saai, S.; Dondorp, A.M.; Lwin, K.M.; et al. Emergence of artemisinin-resistant malaria on the western border of Thailand: A longitudinal study. Lancet 2012, 379, 1960–1966. [Google Scholar] [CrossRef] [Green Version]
- Imwong, M.; Suwannasin, K.; Kunasol, C.; Sutawong, K.; Mayxay, M.; Rekol, H.; Smithuis, F.M.; Hlaing, T.M.; Tun, K.M.; van der Pluijm, R.W.; et al. The spread of artemisinin-resistant Plasmodium falciparum in the Greater Mekong subregion: A molecular epidemiology observational study. Lancet Infect. Dis. 2017, 17, 491–497. [Google Scholar] [CrossRef] [Green Version]
- Ashley, E.A.; Dhorda, M.; Fairhurst, R.M.; Amaratunga, C.; Lim, P.; Suon, S.; Sreng, S.; Anderson, J.M.; Mao, S.; Sam, B.; et al. Spread of artemisinin resistance in Plasmodium falciparum malaria. N. Engl. J. Med. 2014, 371, 411–423. [Google Scholar] [CrossRef] [Green Version]
- Ménard, D.; Khim, N.; Beghain, J.; Adegnika, A.A.; Alam, M.S.; Amodu, O.; Rahim-Awab, G.; Barnadas, C.; Berry, A.; Boum, Y.; et al. A worldwide map of Plasmodium falciparum K13-propeller polymorphisms. N. Engl. J. Med. 2016, 374, 2453–2464. [Google Scholar] [CrossRef]
- Madamet, M.; Kouta, M.B.; Wade, K.A.; Lo, G.; Diawara, S.; Fall, M.; Bercion, R.; Nakoulima, A.; Fall, K.B.; Benoit, N.; et al. Absence of association between polymorphisms in the K13 gene and the presence of Plasmodium falciparum parasites at day 3 after treatment with artemisinin derivatives in Senegal. Int. J. Antimicrob. Agents 2017, 49, 754–756. [Google Scholar] [CrossRef]
- Dieye, B.; Affara, M.; Sangare, L.; Joof, F.; Ndiaye, Y.D.; Gomis, J.F.; Ndiaye, M.; Mbaye, A.; Diakite, M.; Sy, N.; et al. West Africa international centers of excellence for malaria research: Drug resistance patterns to artemether-lumefantrine in Senegal, Mali, and The Gambia. Am. J. Trop. Med. Hyg. 2016, 95, 1054–1060. [Google Scholar] [CrossRef]
- Diallo, M.A.; Yade, M.S.; Ndiaye, Y.D.; Diallo, I.; Diongue, K.; Sy, S.A.; Sy, M.; Seck, M.C.; Ndiaye, M.; Dieye, B.; et al. Efficacy and safety of artemisinin-based combination therapy and the implications of Pfkelch13 and Pfcoronin molecular markers in treatment failure in Senegal. Sci. Rep. 2020, 10, 8907. [Google Scholar] [CrossRef] [PubMed]
- Delandre, O.; Daffe, S.M.; Gendrot, M.; Diallo, M.N.; Madamet, M.; Kounta, M.B.; Diop, M.N.; Bercion, R.; Sow, A.; Ngom, P.M.; et al. Absence of association between polymorphisms in the pfcoronin and pfk13 genes and the presence of Plasmodium falciparum parasites after treatment with artemisinin derivatives in Senegal. Int. J. Antimicrob. Agents 2020, 56, 106190. [Google Scholar] [PubMed]
- Gobbi, F.; Buonfrate, D.; Menegon, M.; Lunardi, G.; Angheben, A.; Severini, C.; Gori, S.; Bisoffi, Z. Failure of dihydroartemisinin-piperaquine treatment of uncomplicated Plasmodium falciparum malaria in a traveller coming from Ethiopia. Malar. J. 2016, 15, 525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, G.; L’Episcopia, M.; Menegon, M.; Souza, S.S.; Dongho, B.G.D.; Vullo, V.; Lucchi, N.W.; Severini, C. Dihydroartemisinin-piperaquine treatment failure in uncomplicated Plasmodium falciparum malaria case imported from Ethiopia. Infection 2018, 46, 867–870. [Google Scholar] [CrossRef]
- Sutherland, C.J.; Lansdell, P.; Sanders, M.; Muwanguzi, J.; van Schalkwyk, D.A.; Kaur, H.; Nolder, D.; Tucker, J.; Bennett, H.M.; Otto, T.D.; et al. pfk13-independent treatment failure in four imported cases of Plasmodium falciparum malaria treated with artemether-lumefantrine in the United Kingdom. Antimicrob. Agents Chemother. 2017, 61, e02382-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plucinski, M.M.; Dimbu, P.R.; Macaia, A.P.; Ferreira, C.M.; Samutondo, C.; Quivinja, J.; Afonso, M.; Kiniffo, R.; Mbounga, E.; Kelley, J.S.; et al. Efficacy of artemether-lumefantrine, artesunate-amodiaquine, and dihydroartemisinin-piperaquine for treatment of uncomplicated Plasmodium falciparum malaria in Angola, 2015. Malar. J. 2017, 16, 62. [Google Scholar] [CrossRef] [Green Version]
- Malvy, D.; Torrentino-Madamet, M.; L’Ollivier, C.; Receveur, M.C.; Jeddi, F.; Delhaes, L.; Piarroux, R.; Millet, P.; Pradines, B. Plasmodium falciparum recrudescence two years after treatment of an uncomplicated infection without return to an area where malaria is endemic. Antimicrob. Agents Chemother. 2018, 62, e01892-17. [Google Scholar] [CrossRef] [Green Version]
- Report on Antimalarial Drug Efficacy, Resistance and Responses (2010–2019). Available online: https://www.who.int/publications/i/item/9789240012813 (accessed on 9 July 2021).
- Ariey, F.; Witkoxski, B.; Amaratunga, C.; Beghain, J.; Langlois, A.C.; Khim, N.; Kim, S.; Duru, V.; Bouchier, C.; Ma, L.; et al. A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature 2014, 505, 50–55. [Google Scholar] [CrossRef]
- Foguim Tsombeng, F.; Gendrot, M.; Robert, M.G.; Madamet, M.; Pradines, B. Are k13 and plasmepsin II genes, involved in Plasmodium falciparum resistance to artemisinin derivatives and piperaquine in Southeast Asia, reliable to monitor resistance surveillance in Africa? Malar. J. 2019, 18, 285. [Google Scholar] [CrossRef] [Green Version]
- Uwimana, A.; Legrand, E.; Stokes, B.H.; Mangala Ndikumana, J.L.; Warsame, M.; Umulisa, N.; Ngamije, D.; Munyaneza, T.; Mazarati, J.B.; Munguti, K.; et al. Emergence and clonal expansion of in vitro artemisinin-resistant Plasmodium falciparum kelch13 R561H mutant parasites in Rwanda. Nat. Med. 2020, 26, 1602–1608. [Google Scholar] [CrossRef]
- Bergmann, C.; van Loon, W.; Habarugira, F.; Tacoli, C.; Jäger, J.C.; Savelsberg, D.; Nshimiyimana, F.; Rwamugema, E.; Mbarushimana, D.; Ndoli, J.; et al. Increase in Kelch 13 polymorphisms in Plasmodium falciparum, Southern Rwanda. Emerg. Infect. Dis. 2021, 27, 294–296. [Google Scholar] [CrossRef]
- Straimer, J.; Gandhi, P.; Czermak Renner, K.; Schmitt, E.K. High prevalence of P. falciparum K13 mutations in Rwanda is associated with slow clearance after treatment with artemether-lumefantrine. J. Infect. Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Uwimana, A.; Umulisa, N.; Venkatesan, M.; Svigel, S.S.; Zhou, Z.; Munyanesa, T.; Habimana, R.M.; Rucogoza, A.; Moriarty, L.F.; Sandford, R.; et al. Association of Plasmodium falciparum kelch13 R561H genotypes with delayed parasite clearance in Rwanda: An open-label, single-arm, multicentre, therapeutic efficacy study. Lancet Infect. Dis. 2021, 21, 1120–1128. [Google Scholar] [CrossRef]
- Demas, A.R.; Sharma, A.I.; Wong, W.; Early, A.M.; Redmond, S.; Bopp, S.; Neafsey, D.E.; Volkman, S.K.; Hartl, D.L.; Wirth, D.F. Mutations in Plasmodium falciparum actin-binding protein coronin confer reduced artemisinin susceptibility. Proc. Natl. Acad. Sci. USA 2018, 115, 12799–12804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.I.; Shin, S.H.; Bopp, S.; Volkman, S.K.; Hartl, D.L.; Wirth, D.F. Genetic background and PfKelch13 affect artemisinin susceptibility of PfCoronin mutants in Plasmodium falciparum. PLoS Genet. 2020, 16, e1009266. [Google Scholar] [CrossRef] [PubMed]
- Velavan, T.P.; Nderu, D.; Agbenyega, T.; Ntoumi, F.; Kremsner, P.G. An alternative dogma on reduced artemisinin susceptibility: A new shadow from east to west. Proc. Natl. Acad. Sci. USA 2019, 116, 12611–12612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.Q.; Pernaute-Lau, L.; Khattak, A.A.; Luijcx, S.; Aydin-Schmidt, B.; Hussain, M.; Khan, T.A.; Mufti, F.U.; Morris, U. Surveillance of genetic markers associated with Plasmodium falciparum resistance to artemisinin-based combination therapy in Pakistan, 2018–2019. Malar. J. 2020, 19, 206. [Google Scholar] [CrossRef]
- Bane, K.S.; Lepper, S.; Kehrer, J.; Sattler, J.M.; Singer, M.; Reining, M.; Klug, D.; Heiss, K.; Baum, J.; Mueller, A.K.; et al. The actin filament-binding protein coronin regulates motility in Plasmodium sporozoites. PLoS Pathog. 2016, 12, e1005710. [Google Scholar] [CrossRef] [Green Version]
- Olshina, M.A.; Angrisano, F.; Marapana, D.S.; Riglar, D.T.; Bane, K.; Wong, W.; Catimel, B.; Yin, M.X.; Holmes, A.B.; Frischknecht, F.; et al. Plasmodium falciparum coronin organizes arrays of parallel actin filaments potentially guiding directional motility in invasive malaria parasites. Malar. J. 2015, 14, 280. [Google Scholar] [CrossRef] [Green Version]
- Wurtz, N.; Fall, B.; Bui, K.; Pascual, A.; Fall, M.; Camara, C.; Diatta, B.; Fall, K.B.; Mbaye, P.S.; Diémé, Y.; et al. Pfhrp2 and Pfhrp3 polymorphisms in Plasmodium falciparum isolates from Dakar, Senegal: Impact on rapid malaria diagnosis. Malar. J. 2013, 12, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gendrot, M.; Delandre, O.; Robert, M.G.; Foguim, F.T.; Benoit, N.; Amalvict, R.; Fonta, I.; Mosnier, J.; Madamet, M.; Pradines, B. Absence of association between methylene blue reduced susceptibility and polymorphisms in 12 genes involved in antimalarial drug resistance in African Plasmodium falciparum. Pharmaceuticals 2021, 14, 351. [Google Scholar] [CrossRef] [PubMed]
- Gendrot, M.; Foguim, F.T.; Robert, M.G.; Amalvict, R.; Mosnier, J.; Benoit, N.; Madamet, M.; Pradines, B. The D113N mutation in the RING E3 ubiquitin protein ligase gene is not associated with ex vivo susceptibility to common anti-malarial drugs in African Plasmodium falciparum isolates. Malar. J. 2018, 17, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ICEstimator Version 1.2. Available online: https://www.antimalarial-icestimator.net (accessed on 13 July 2021).
- Pascual, A.; Madamet, M.; Briolant, S.; Gaillard, T.; Amalvict, R.; Benoit, N.; Travers, D.; Pradines, B. Multinormal in vitro distribution of Plasmodium falciparum susceptibility to piperaquine and pyronaridine. Malar. J. 2015, 14, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradines, B.; Bertaux, L.; Pomares, C.; Delaunay, P.; Marty, P. Reduced in vitro susceptibility to artemisinin derivatives associated with multi-resistance in a traveller returning from South-East Asia. Malar. J. 2011, 10, 268. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Areas | Isolates No and (%) | P76S Mutation No and % a | V62M Mutation No and % a |
|---|---|---|---|
| West Africa | 357 (40.8%) | 74 (8.5%) | 3 (0.3%) |
| Central Africa | 335 (38.3%) | 40 (4.5%) | 6 (0.7%) |
| East Africa | 143 (16.3%) | 6 (0.7%) | 0 |
| Unspecified country in Africa | 39 (4.6%) | 5 (0.6%) | 0 |
| Total | 874 | 125 (14.3%) | 9 (1.0%) |
| Country | 2018 No and % | 2019 No and % | 2018–2019 No and % | |||
|---|---|---|---|---|---|---|
| 76S Mutation | 62M Mutation | 76S Mutation | 62M Mutation | 76S Mutation | 62M Mutation | |
| West Africa | 38/179 (21.2) | 2/179 (1.1) | 36/178 (20.2) | 1/178 (0.6) | 74/357 (20.7) | 3/357 (0.8) |
| Côte d’Ivoire | 20/72 (27.8) | 0/20 | 14/69 (20.3) | 0/69 | 34/141 (24.1) | 0/141 |
| Guinea | 4/25 (16.0) | 0/25 | 6/29 (20.7) | 0/29 | 10/54 (18.5) | 0/54 |
| Togo | 1/14 (7.1) | 0/14 | 2/20 (10.0) | 0/20 | 3/34 (8.8) | 0/34 |
| Burkina Faso | 4/19 (21.1) | 2/19 (10.5) | 2/10 (20.0) | 0/10 | 6/29 (20.7) | 2/29 (6.9) |
| Benin | 2/13 (15.4) | 0/13 | 4/12 (33.3) | 0/12 | 6/25 (24.0) | 0/25 |
| Mali | 2/8 (25.0) | 0/8 | 2/8 (25.0) | 0/8 | 4/16 (25.0) | 0/16 |
| Nigeria | 2/5 (40.0) | 0/5 | 2/12 (16.7) | 1/12 (8.3) | 4/17 (23.5) | 1/17 (5.9) |
| Senegal | 1/8 (12.5) | 0/8 | 0/3 | 0/3 | 1/11 (9.1) | 0/11 |
| Niger | 0/3 | 0/3 | 3/9 (33.3) | 0/9 | 3/12 (25.0) | 0/12 |
| Ghana | 1/4 (25.0) | 0/4 | 1/5 (20.0) | 0/5 | 2/9 (22.2) | 0/9 |
| Sierra Leone | 1/4 (25.0) | 0/4 | 0 | 0 | 1/4 (25.0) | 0/4 |
| Guinea Conakry | 0/3 | 0/3 | 0/1 | 0/1 | 0/4 | 0/4 |
| Cape Verde | 0/1 | 0/1 | 0 | 0 | 0/1 | 0/1 |
| Central Africa | 20/154 (13.0) | 1/154 (0.6) | 20/181 (11.0) | 5/181 (2.8) | 40/335 (11.9) | 6/335 (1.8) |
| Cameroon | 8/68 (11.8) | 0/68 | 11/79 (13.9) | 2/79 (2.5) | 19/147 (12.9) | 2/147 (1.4) |
| Gabon | 3/28 (10.7) | 0/28 | 2/28 (7.1) | 1/28 (3.6) | 5/56 (8.9) | 1/56 (1.8) |
| Democratic Republic of Congo | 2/22 (9.1) | 0/22 | 1/28 (3.6) | 0/28 | 3/50 (6.0) | 0/50 |
| Central African Republic | 6/19 (3.2) | 0/19 | 4/31 (12.9) | 2/31 (6.5) | 10/50 (20.0) | 2/50 (4.0) |
| Chad | 1/13 (7.7) | 1/13 (7.7) | 2/13 (15.4) | 0/13 | 3/26 (11.5) | 2/26 (3.8) |
| Angola | 0/2 | 0/2 | 0/1 | 0/1 | 0/3 | 0/3 |
| Equatorial Guinea | 0/2 | 0/2 | 0/1 | 0/1 | 0/3 | 0/3 |
| East Africa | 1/56 (1.8) | 0/56 | 5/87 (5.7) | 0/87 (0.0) | 6/143 (4.2) | 0/143 (0.0) |
| Mayotte | 1/3 (33.3) | 0/3 | 1/2 (50.0) | 0/2 | 2/5 (40.0) | 0/5 |
| Eritrea | 0 | 0 | 1/1 (100.0) | 0/1 | 1/1 (100.0) | 0/1 |
| Tanzania | 0/1 | 0/1 | 2/7 (28.6) | 0/7 | 2/8 (25.0) | 0/8 |
| Sudan | 0 | 0 | 1/2 (50.0) | 0/2 | 1/2 (50.0) | 0/2 |
| Kenya | 0 | 0 | 0/1 | 0/1 | 0/1 | 0/1 |
| Zambia | 0 | 0 | 0/1 | 0/1 | 0/1 | 0/1 |
| Uganda | 0 | 0 | 0/2 | 0/2 | 0/2 | 0/2 |
| Mozambique | 0/1 | 0/1 | 0 | 0 | 0/1 | 0/1 |
| Djibouti | 0/6 | 0/6 | 0/6 | 0/6 | 0/12 | 0/12 |
| Comoros | 0/40 | 0/40 | 0/64 | 0/64 | 0/104 | 0/104 |
| Madagascar | 0/5 | 0/5 | 0/1 | 0/1 | 0/6 | 0/6 |
| Unspecified African country | 2/8 (25.0) | 0/8 | 3/31 (9.7) | 0/31 (0.0) | 5/39 (12.8) | 0/39 (0.0) |
| Total | 31/397 (15.4) | 3/397 (0.8) | 64/477 (13.4) | 6/477 (1.3) | 128/874 (14.3) | 9/874 (1.0) |
| Drug (Isolates No) | Isolates Wild-Type (P76) | Isolates with 76S Mutation | p-Value (t-Test) | ||
|---|---|---|---|---|---|
| IC50 Geometric Mean (nM) | Min-Max IC50 | IC50 Geometric Mean (nM) | Min-Max IC50 | ||
| Chloroquine (328) | 84.0 | 2.8–764.7 | 95.2 | 8.0–350.9 | 0.463 |
| Quinine (325) | 281.0 | 6.2–757.2 | 268.7 | 23.3–664.7 | 0.617 |
| Lumefantrine (329) | 6.7 | 0.4–68.8 | 6.5 | 0.5–46.0 | 0.878 |
| Desethylamodiaquine (306) | 40.7 | 0.6–775.9 | 34.3 | 2.0–152.5 | 0.343 |
| Mefloquine (321) | 22.9 | 0.7–98.0 | 26.1 | 3.2–87.8 | 0.295 |
| Pyronaridine (319) | 14.0 | 0.2–119.3 | 12.6 | 1.0–56.9 | 0.410 |
| Piperaquine (321) | 25.1 | 5.5–67.1 | 25.2 | 0.7–379.3 | 0.983 |
| Drug (No Isolates) | % of Isolates with Reduced Susceptibility (No of Isolates with Reduced Susceptibility/Total of Isolates) | p-Value (Chi-Squared Test) | |
|---|---|---|---|
| Wild-Type Isolates (P76) | Isolates with 76S Mutation | ||
| Chloroquine (328) | 25.1 (70/279) | 32.7 (16/49) | 0.506 |
| Quinine (325) | 0 (0/276) | 0 (0/49) | 0 |
| Lumefantrine (329) | 0 (0/280) | 0 (0/49) | 0 |
| Desethylamodiaquine (306) | 8.0 (21/262) | 9.1 (4/44) | 1 |
| Mefloquine (321) | 23.4 (64/273) | 29.2 (14/48) | 0.631 |
| Pyronaridine (319) | 1.1 (3/273) | 0 (0/46) | 1 |
| Piperaquine (321) | 0.7 (2/273) | 0 (0/48) | 0.999 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delandre, O.; Gendrot, M.; Fonta, I.; Mosnier, J.; Benoit, N.; Amalvict, R.; Gomez, N.; Madamet, M.; Pradines, B. Prevalence of Mutations in the pfcoronin Gene and Association with Ex Vivo Susceptibility to Common Quinoline Drugs against Plasmodium falciparum. Pharmaceutics 2021, 13, 1273. https://doi.org/10.3390/pharmaceutics13081273
Delandre O, Gendrot M, Fonta I, Mosnier J, Benoit N, Amalvict R, Gomez N, Madamet M, Pradines B. Prevalence of Mutations in the pfcoronin Gene and Association with Ex Vivo Susceptibility to Common Quinoline Drugs against Plasmodium falciparum. Pharmaceutics. 2021; 13(8):1273. https://doi.org/10.3390/pharmaceutics13081273
Chicago/Turabian StyleDelandre, Océane, Mathieu Gendrot, Isabelle Fonta, Joel Mosnier, Nicolas Benoit, Rémy Amalvict, Nicolas Gomez, Marylin Madamet, and Bruno Pradines. 2021. "Prevalence of Mutations in the pfcoronin Gene and Association with Ex Vivo Susceptibility to Common Quinoline Drugs against Plasmodium falciparum" Pharmaceutics 13, no. 8: 1273. https://doi.org/10.3390/pharmaceutics13081273
APA StyleDelandre, O., Gendrot, M., Fonta, I., Mosnier, J., Benoit, N., Amalvict, R., Gomez, N., Madamet, M., & Pradines, B. (2021). Prevalence of Mutations in the pfcoronin Gene and Association with Ex Vivo Susceptibility to Common Quinoline Drugs against Plasmodium falciparum. Pharmaceutics, 13(8), 1273. https://doi.org/10.3390/pharmaceutics13081273