
Polylactide Nanocapsules Attenuate Adverse Cardiac Cellular Effects of Lyso-7, a Pan-PPAR Agonist/Anti-Inflammatory New Thiazolidinedione

 , ,
, ,  and
and 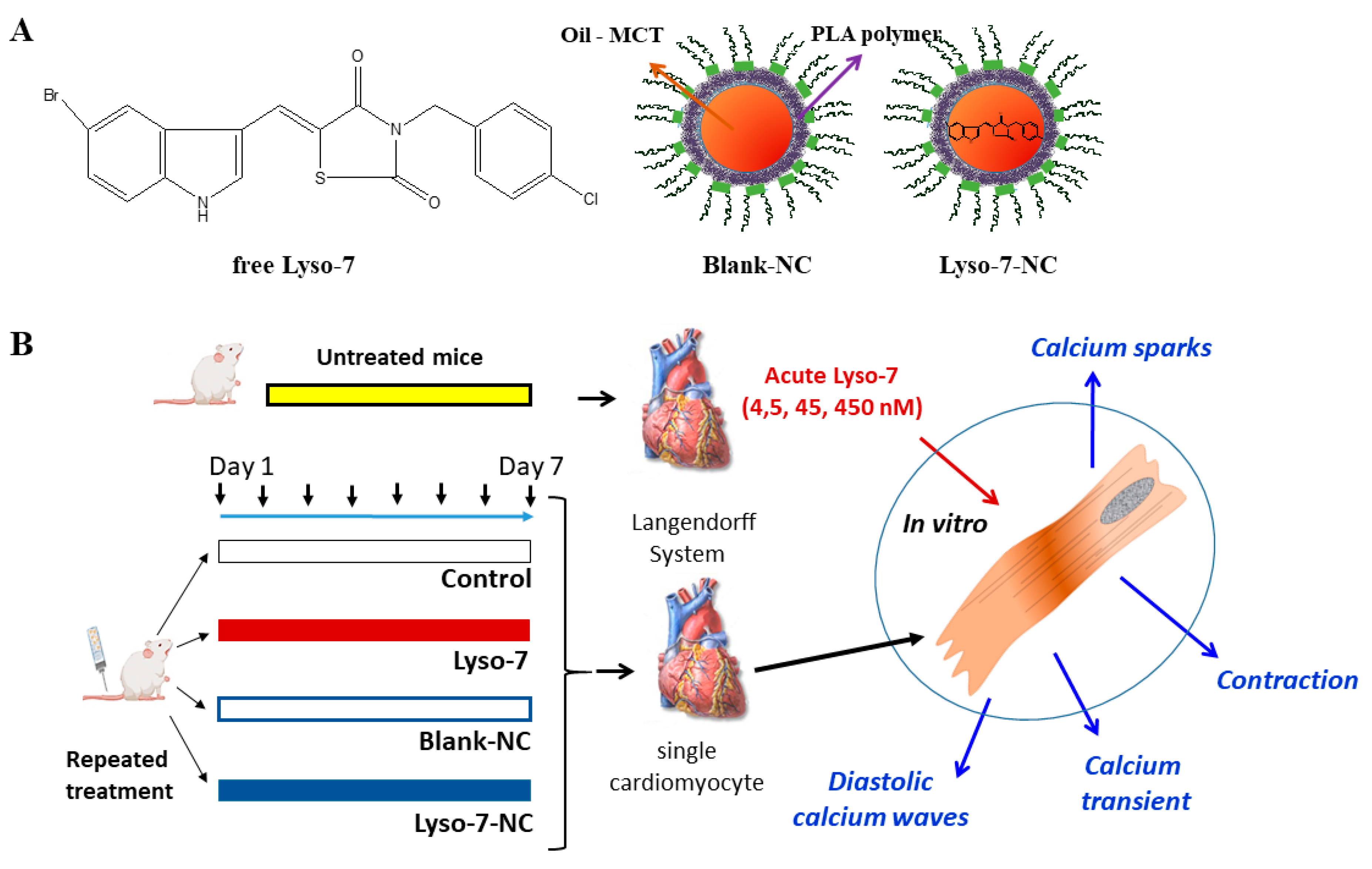{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Lyso-7 Solution and Lyso-7-Loaded Nanocapsules
2.3. Animals and Cardiomyocytes
2.4. Measurements of Contraction and Intracellular Ca2+ in Single Cardiomyocytes
2.5. Sparks Confocal
2.6. Statistical Analysis
3. Results
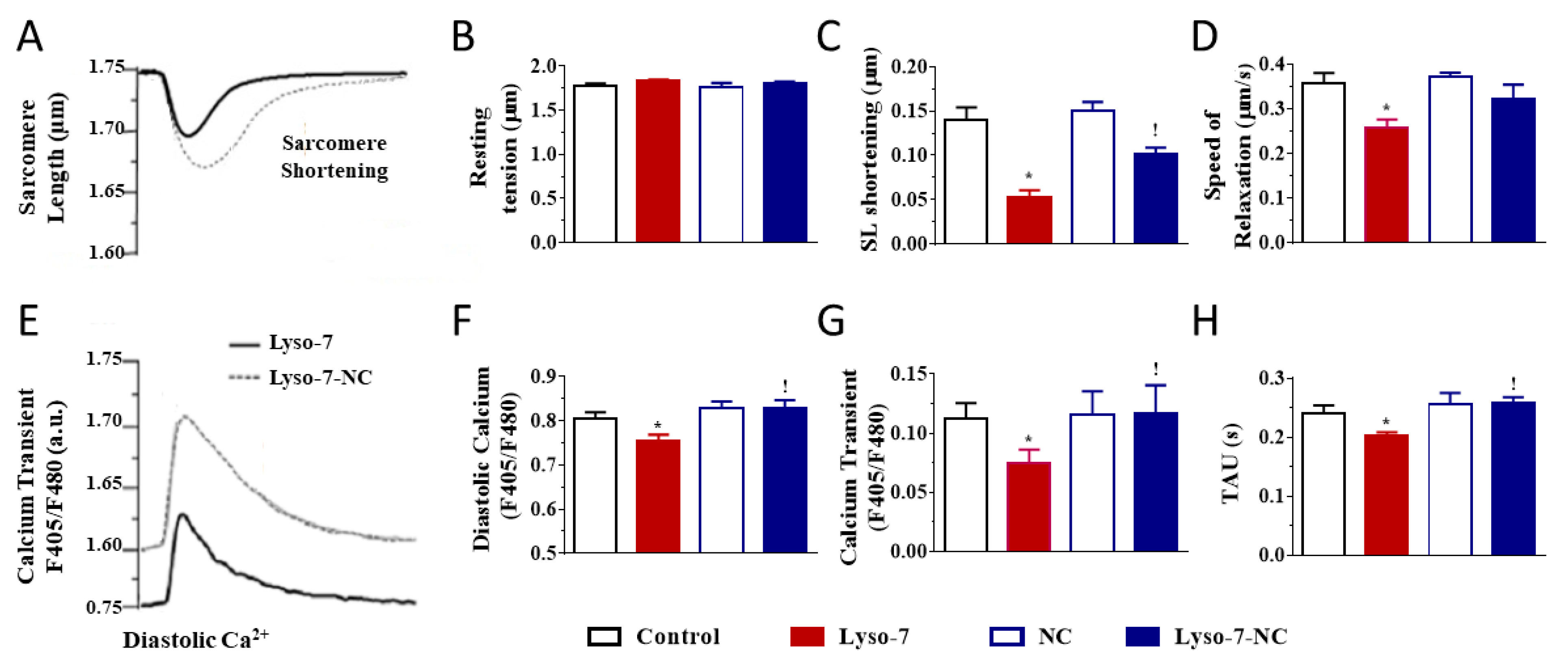
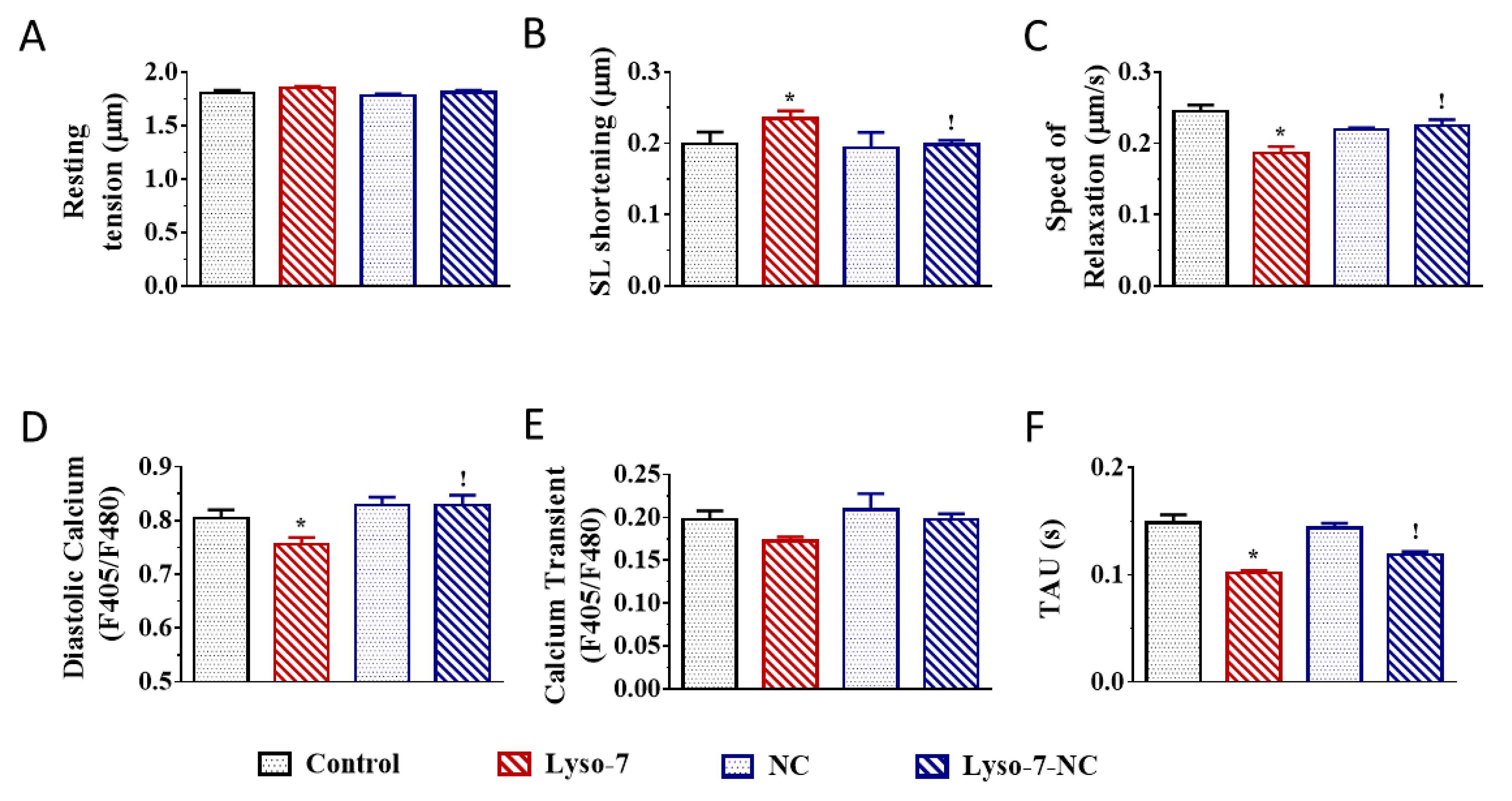
3.1. Effect of In Vivo Repeated Dose Administration of Lyso-7 on Single Myocytes
3.1.1. Differential Effects of Lyso-7 and Lyso-7-NC on Cellular Contraction and Ca2+
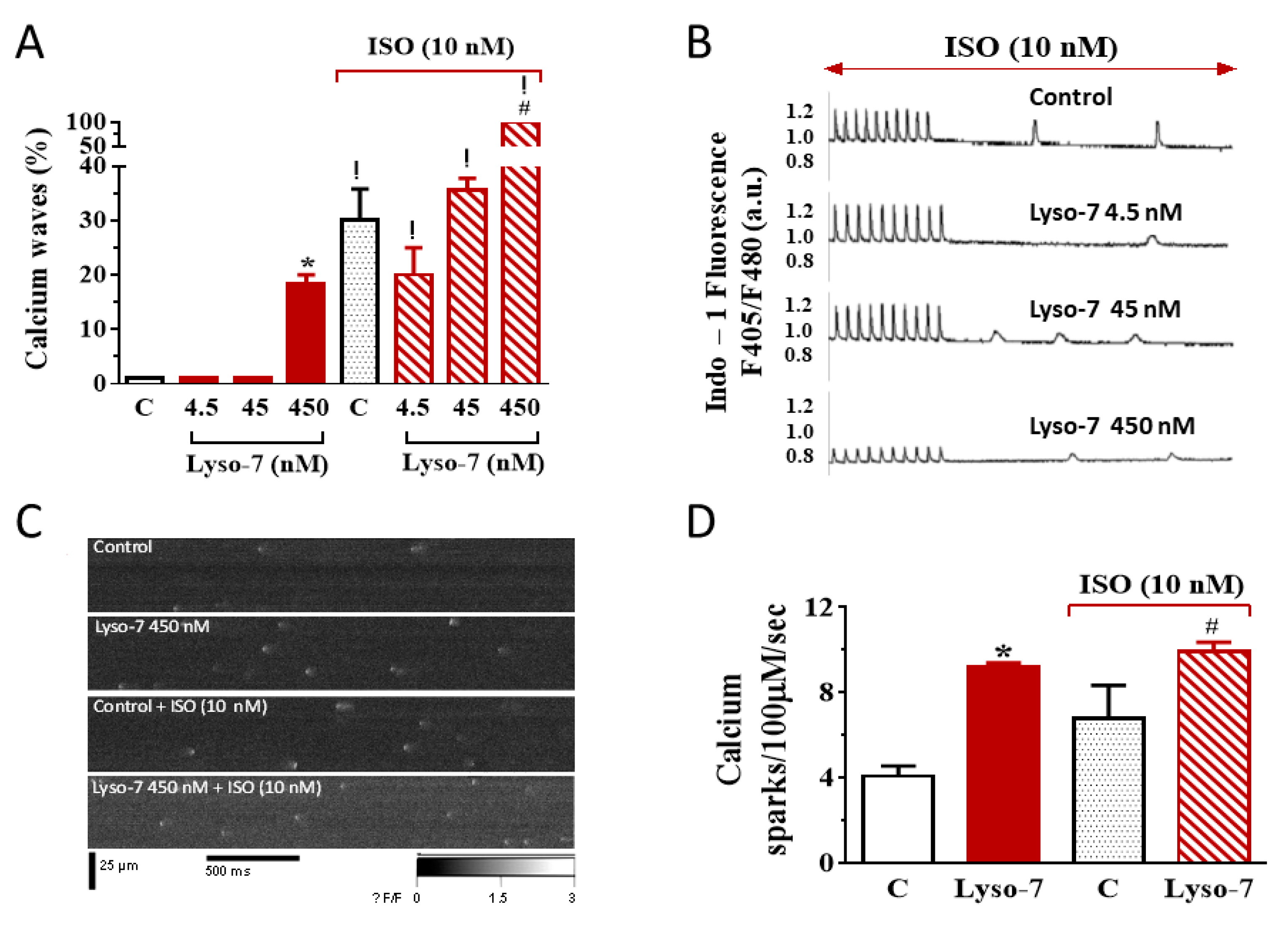
3.1.2. Differential Effects of Lyso-7 and Lyso-7-NC on Ectopic Ca2+ Events at Rest
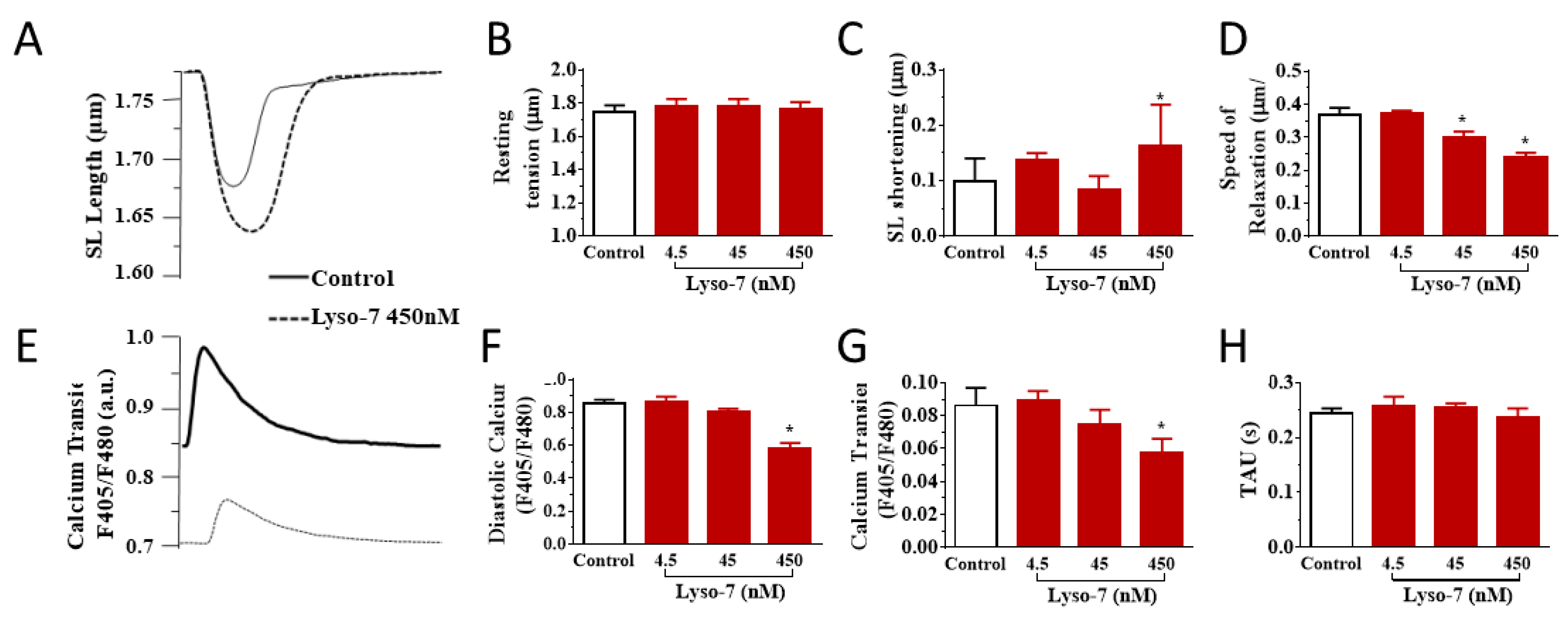
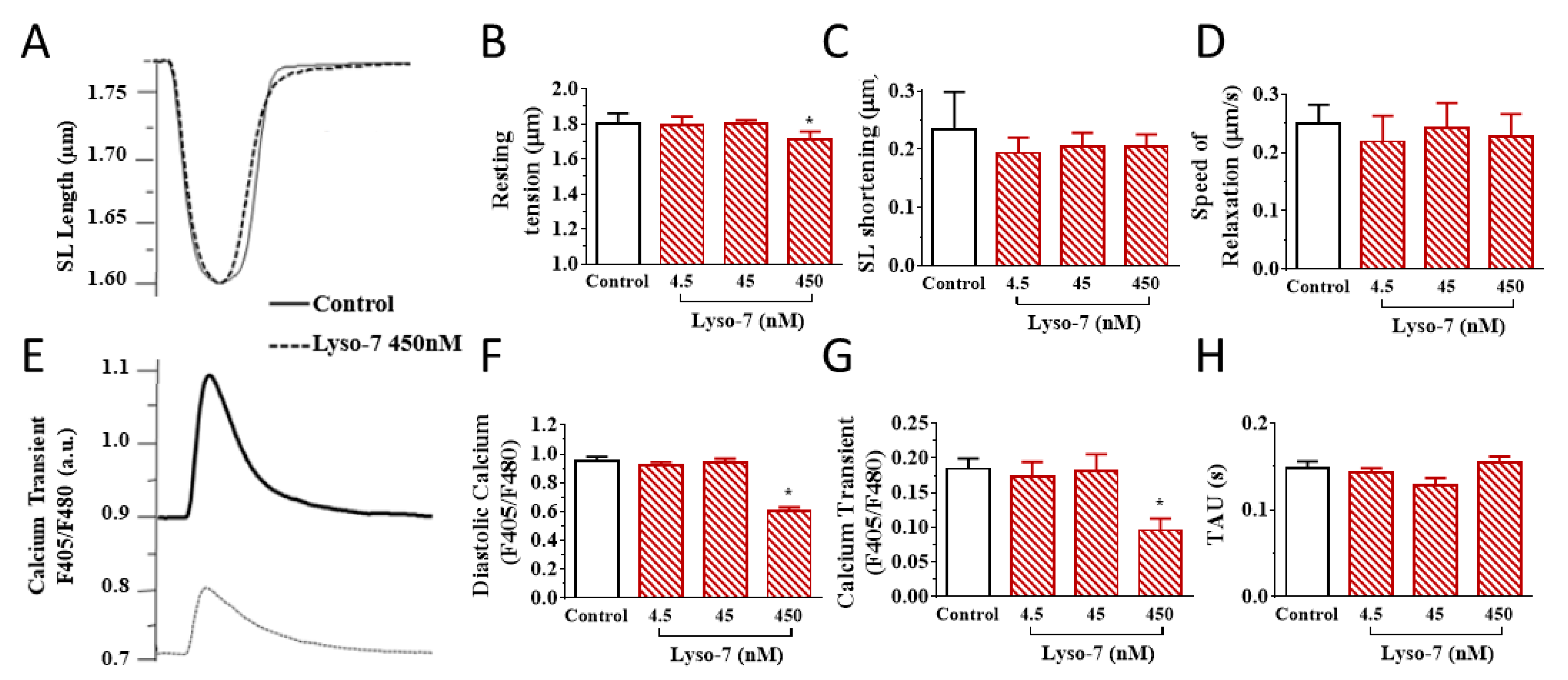
3.2. Acute Effects of Free Lyso-7 on Single Cardiomyocytes In Vitro
3.2.1. Lyso-7 Impairs Contraction and Ca2+ Transient
3.2.2. Lyso-7 Promotes Abnormal Ca2+ Events at Rest
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Michalik, L.; Auwerx, J.; Berger, J.P.; Chatterjee, V.K.; Glass, C.K.; Gonzalez, F.J.; Grimaldi, P.A.; Kadowaki, T.; Lazar, M.A.; O’Rahilly, S.; et al. International Union of Pharmacology. LXI. Peroxisome Proliferator-Activated Receptors. Pharmacol. Rev. 2006, 58, 726–741. [Google Scholar] [CrossRef]
- Tan, C.K.; Zhuang, Y.; Wahli, W. Synthetic and natural Peroxisome Proliferator-Activated Receptor (PPAR) agonists as candidates for the therapy of the metabolic syndrome. Expert Opin. Ther. Targets 2017, 21, 333–348. [Google Scholar] [CrossRef]
- Hamblin, M.; Chang, L.; Fan, Y.; Zhang, J.; Chen, Y.E. PPARs and the Cardiovascular System. Antioxid. Redox Signal. 2009, 11, 1415–1452. [Google Scholar] [CrossRef]
- Orasanu, G.; Ziouzenkova, O.; Devchand, P.R.; Nehra, V.; Hamdy, O.; Horton, E.S.; Plutzky, J. The Peroxisome Proliferator-Activated Receptor-γ Agonist Pioglitazone Represses Inflammation in a Peroxisome Proliferator-Activated Receptor-α–Dependent Manner In Vitro and In Vivo in Mice. J. Am. Coll. Cardiol. 2008, 52, 869–881. [Google Scholar] [CrossRef] [Green Version]
- Diamant, M.; Heine, R.J. Thiazolidinediones in Type 2 Diabetes Mellitus. Drugs 2003, 63, 1373–1405. [Google Scholar] [CrossRef] [PubMed]
- Lincoff, A.M.; Wolski, K.; Nicholls, S.; Nissen, S.E. Pioglitazone and Risk of Cardiovascular Events in Patients with Type 2 Diabetes Mellitus. JAMA 2007, 298, 1180–1188. [Google Scholar] [CrossRef]
- DeLea, T.E.; Edelsberg, J.S.; Hagiwara, M.; Oster, G.; Phillips, L.S. Use of thiazolidinediones and risk of heart failure in people with type 2 diabetes: A retrospective cohort study. Diabetes Care 2003, 26, 2983–2989. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.Y.; Fantus, I.G. Thiazolidinedione-Induced Congestive Heart Failure. Ann. Pharmacother. 2004, 38, 817–820. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, A.V.; Usmani, A.; Rajamanickam, A.; Moheet, A. Thiazolidinediones and Risk of Heart Failure in Patients with or at High Risk of Type 2 Diabetes Mellitus. Am. J. Cardiovasc. Drugs 2011, 11, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.J.; Ouellet-Hellstrom, R.; MaCurdy, T.E.; Ali, F.; Sholley, C.; Worrall, C.; Kelman, J.A. Risk of Acute Myocardial Infarction, Stroke, Heart Failure, and Death in Elderly Medicare Patients Treated with Rosiglitazone or Pioglitazone. JAMA 2010, 304, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Santin, J.R.; Uchôa, F.D.; Lima, M.D.C.A.; Rabello, M.; Machado, I.D.; Hernandes, M.Z.; Amato, A.A.; Milton, F.A.; Webb, P.; Neves, F.D.A.R.; et al. Chemical synthesis, docking studies and biological effects of a pan peroxisome proliferator-activated receptor agonist and cyclooxygenase inhibitor. Eur. J. Pharm. Sci. 2013, 48, 689–697. [Google Scholar] [CrossRef]
- Santin, J.R.; Machado, I.D.; Rodrigues, S.F.P.; Teixeira, S.; Muscará, M.N.; Galdino, S.L.; Pitta, I.D.R.; Farsky, S.H.P. Role of an Indole-Thiazolidine Molecule PPAR Pan-Agonist and COX Inhibitor on Inflammation and Microcirculatory Damage in Acute Gastric Lesions. PLoS ONE 2013, 8, e76894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- César, F.A.; Rudnicki, M.; Heras, B.D.L.; Boscá, L.; Lima, M.C.; Pitta, I.R.; Abdalla, D.S. New indole-thiazolidine attenuates atherosclerosis in LDLr−/− mice. Vasc. Pharmacol. 2015, 71, 174–180. [Google Scholar] [CrossRef]
- Santin, J.R.; Machado, I.D.; Drewes, C.C.; Kupa, L.D.V.K.; Soares, R.M.; Cavalcanti, D.M.; Pitta, I.D.R.; Farsky, S.H. Role of an indole-thiazolidiene PPAR pan ligand on actions elicited by G-protein coupled receptor activated neutrophils. Biomed. Pharmacother. 2018, 105, 947–955. [Google Scholar] [CrossRef]
- Mosqueira, V.C.; Legrand, P.; Pinto-Alphandary, H.; Puisieux, F.; Barratt, G. Poly(d,l-Lactide) Nanocapsules Prepared by a Solvent Displacement Process: Influence of the Composition on Physicochemical and Structural Properties. J. Pharm. Sci. 2000, 89, 614–626. [Google Scholar] [CrossRef]
- Degobert, G.; Aydin, D. Lyophilization of Nanocapsules: Instability Sources, Formulation and Process Parameters. Pharmaceutics 2021, 13, 1112. [Google Scholar] [CrossRef] [PubMed]
- Leite, E.A.; Grabe-Guimarães, A.; Guimarães, H.N.; Machado-Coelho, G.L.L.; Barratt, G.; Mosqueira, V.C. Cardiotoxicity reduction induced by halofantrine entrapped in nanocapsule devices. Life Sci. 2007, 80, 1327–1334. [Google Scholar] [CrossRef] [Green Version]
- Branquinho, R.T.; Roy, J.; Farah, C.; Garcia, G.M.; Aimond, F.; Le Guennec, J.-Y.; Saúde-Guimarães, D.; Grabe-Guimaraes, A.; Mosqueira, V.; Lana, M.; et al. Biodegradable Polymeric Nanocapsules Prevent Cardiotoxicity of Anti-Trypanosomal Lychnopholide. Sci. Rep. 2017, 7, srep44998. [Google Scholar] [CrossRef] [Green Version]
- Souza, A.C.M.; Mosqueira, V.C.F.; Silveira, A.P.A.; Antunes, L.R.; Richard, S.; Guimarães, H.N.; Grabe-Guimarães, A. Reduced cardiotoxicity and increased oral efficacy of artemether polymeric nanocapsules in Plasmodium berghei-infected mice. Parasitology 2017, 145, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.C.M.; Grabe-Guimarães, A.; Cruz, J.D.S.; Santos-Miranda, A.; Farah, C.; Oliveira, L.T.; Lucas, A.; Aimond, F.; Sicard, P.; Mosqueira, V.C.F.; et al. Mechanisms of artemether toxicity on single cardiomyocytes and protective effect of nanoencapsulation. Br. J. Pharmacol. 2020. [Google Scholar] [CrossRef]
- Garcia, G.M.; Oliveira, L.T.; Pitta, I.D.R.; De Lima, M.D.C.A.; Vilela, J.M.C.; Andrade, M.S.; Abdalla, D.S.P.; Mosqueira, V.C.F. Improved nonclinical pharmacokinetics and biodistribution of a new PPAR pan-agonist and COX inhibitor in nanocapsule formulation. J. Control. Release 2015, 209, 207–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thireau, J.; Karam, S.; Fauconnier, J.; Roberge, S.; Cassan, C.; Cazorla, O.; Aimond, F.; Lacampagne, A.; Babuty, M.; Richard, S. Functional evidence for an active role of B-type natriuretic peptide in cardiac remodelling and pro-arrhythmogenicity. Cardiovasc. Res. 2012, 95, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Thireau, J.; Karam, S.; Roberge, S.; Roussel, J.; Aimond, F.; Cassan, C.; Gac, A.; Babuty, D.; Le Guennec, J.-Y.; Lacampagne, A.; et al. β-Adrenergic blockade combined with subcutaneous B-type natriuretic peptide: A promising approach to reduce ventricular arrhythmia in heart failure? Heart 2014, 100, 833–841. [Google Scholar] [CrossRef] [Green Version]
- Zalvidea, S.; Andre, L.; Loyer, X.; Cassan, C.; Sainte-Marie, Y.; Thireau, J.; Sjaastad, I.; Heymes, C.; Pasquié, J.-L.; Cazorla, O.; et al. ACE Inhibition Prevents Diastolic Ca2+ Overload and Loss of Myofilament Ca2+ Sensitivity after Myocardial Infarction. Curr. Mol. Med. 2012, 12, 206–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, H.E.; Rodrigo, G.C. Inotropic Response of Cardiac Ventricular Myocytes to β-Adrenergic Stimulation with Isoproterenol Exhibits Diurnal Variation. Circ. Res. 2010, 106, 1244–1252. [Google Scholar] [CrossRef] [Green Version]
- Eisner, D.A.; Caldwell, J.L.; Trafford, A.W.; Hutchings, D. The Control of Diastolic Calcium in the Heart. Circ. Res. 2020, 126, 395–412. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Lederer, W.J. Calcium Sparks. Physiol. Rev. 2008, 88, 1491–1545. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Velasco, M.; Rueda, A.; Rizzi, N.; Benitah, J.-P.; Colombi, B.; Napolitano, C.; Priori, S.G.; Richard, S.; Gómez, A.M. Increased Ca 2+ Sensitivity of the Ryanodine Receptor Mutant RyR2 R4496C Underlies Catecholaminergic Polymorphic Ventricular Tachycardia. Circ. Res. 2009, 104, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Santiago, D.J.; Ríos, E.; Shannon, T.R. Isoproterenol Increases the Fraction of Spark-Dependent RyR-Mediated Leak in Ventricular Myocytes. Biophys. J. 2013, 104, 976–985. [Google Scholar] [CrossRef] [Green Version]
- Mechmann, S.; Pott, L. Identification of Na-Ca exchange current in single cardiac myocytes. Nature 1986, 319, 597–599. [Google Scholar] [CrossRef]
- Shattock, M.J.; Ottolia, M.; Bers, D.; Blaustein, M.P.; Boguslavskyi, A.; Bossuyt, J.; Bridge, J.H.B.; Chen-Izu, Y.; Clancy, C.E.; Edwards, A.; et al. Na+/Ca2+exchange and Na+/K+-ATPase in the heart. J. Physiol. 2015, 593, 1361–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venetucci, L.A.; Trafford, A.W.; Eisner, D.A. Increasing Ryanodine Receptor Open Probability Alone Does Not Produce Arrhythmogenic Calcium Waves. Circ. Res. 2007, 100, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabiato, A.; Fabiato, F. Calcium and cardiac excitation-contraction coupling. Annu. Rev. Physiol. 1979, 41. [Google Scholar] [CrossRef] [PubMed]
- Nissen, S.E.; Wolski, K. Effect of Rosiglitazone on the Risk of Myocardial Infarction and Death from Cardiovascular Causes. N. Engl. J. Med. 2007, 356, 2457–2471. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia, G.M.; Roy, J.; Pitta, I.R.; Abdalla, D.S.P.; Grabe-Guimarães, A.; Mosqueira, V.C.F.; Richard, S. Polylactide Nanocapsules Attenuate Adverse Cardiac Cellular Effects of Lyso-7, a Pan-PPAR Agonist/Anti-Inflammatory New Thiazolidinedione. Pharmaceutics 2021, 13, 1521. https://doi.org/10.3390/pharmaceutics13091521
Garcia GM, Roy J, Pitta IR, Abdalla DSP, Grabe-Guimarães A, Mosqueira VCF, Richard S. Polylactide Nanocapsules Attenuate Adverse Cardiac Cellular Effects of Lyso-7, a Pan-PPAR Agonist/Anti-Inflammatory New Thiazolidinedione. Pharmaceutics. 2021; 13(9):1521. https://doi.org/10.3390/pharmaceutics13091521
Chicago/Turabian StyleGarcia, Giani M., Jérôme Roy, Ivan R. Pitta, Dulcinéia S. P. Abdalla, Andrea Grabe-Guimarães, Vanessa C. F. Mosqueira, and Sylvain Richard. 2021. "Polylactide Nanocapsules Attenuate Adverse Cardiac Cellular Effects of Lyso-7, a Pan-PPAR Agonist/Anti-Inflammatory New Thiazolidinedione" Pharmaceutics 13, no. 9: 1521. https://doi.org/10.3390/pharmaceutics13091521
APA StyleGarcia, G. M., Roy, J., Pitta, I. R., Abdalla, D. S. P., Grabe-Guimarães, A., Mosqueira, V. C. F., & Richard, S. (2021). Polylactide Nanocapsules Attenuate Adverse Cardiac Cellular Effects of Lyso-7, a Pan-PPAR Agonist/Anti-Inflammatory New Thiazolidinedione. Pharmaceutics, 13(9), 1521. https://doi.org/10.3390/pharmaceutics13091521