

Gossypol and Its Natural Derivatives: Multitargeted Phytochemicals as Potential Drug Candidates for Oncologic Diseases



Abstract
:1. Introduction
2. Sources and Distribution of Gossypol
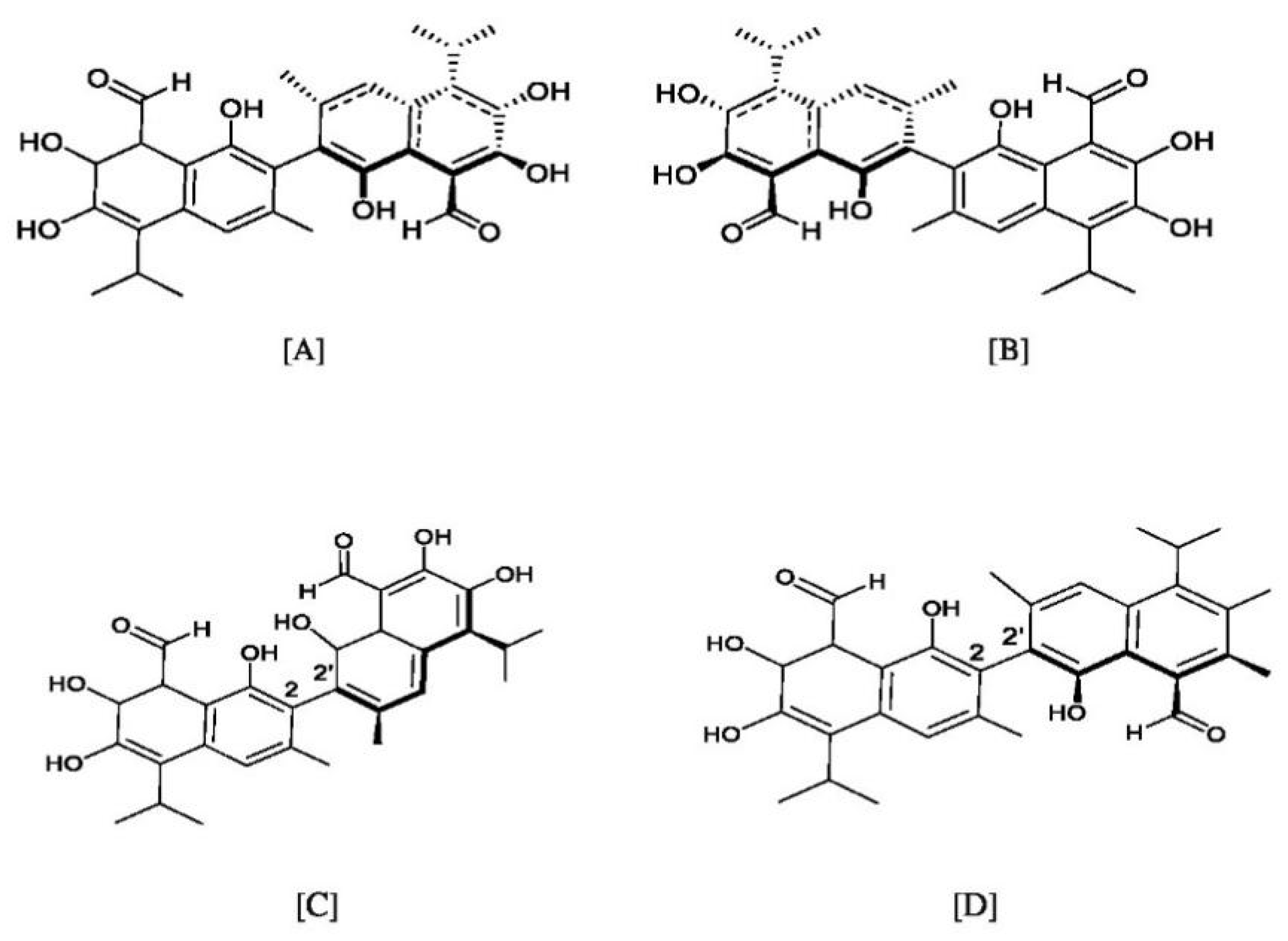
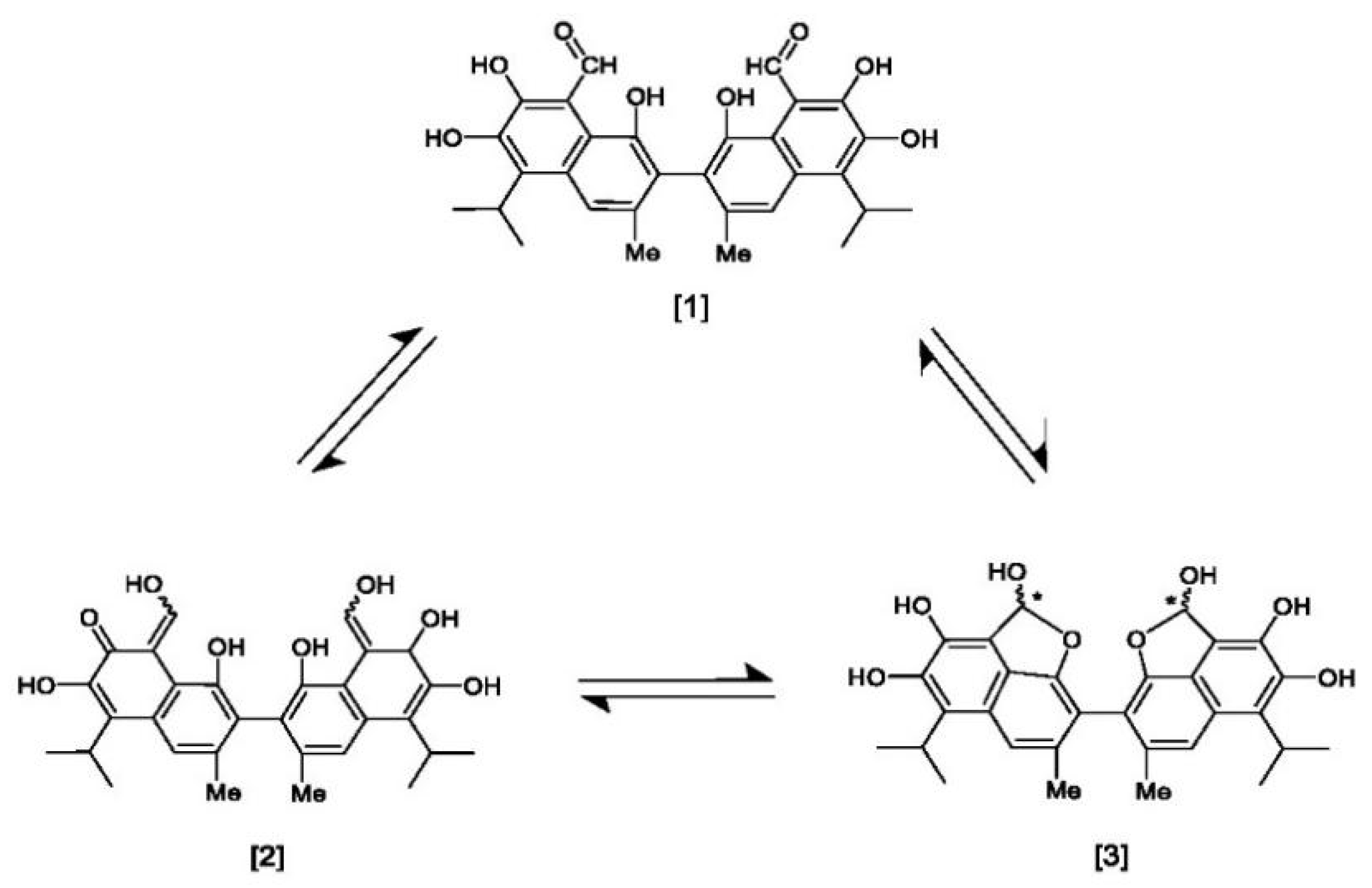
3. Chemistry of Gossypol: Structure, Atropisomerism, Stereochemistry, and Tautomerism
3.1. Atropisomerism
3.2. Tautomerism
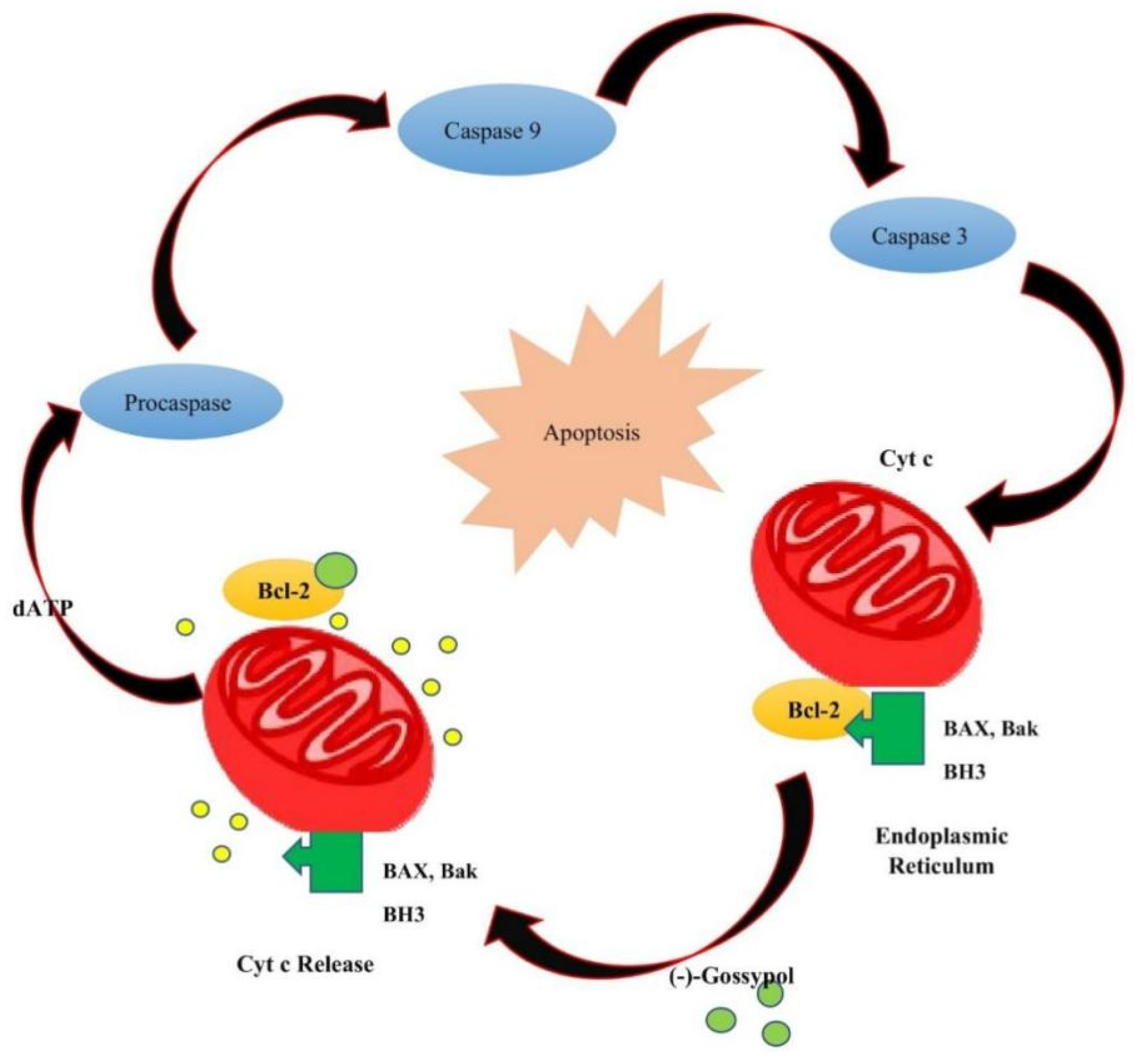
4. Toxicity of Gossypol
5. Pharmacokinetic Profile of Gossypol
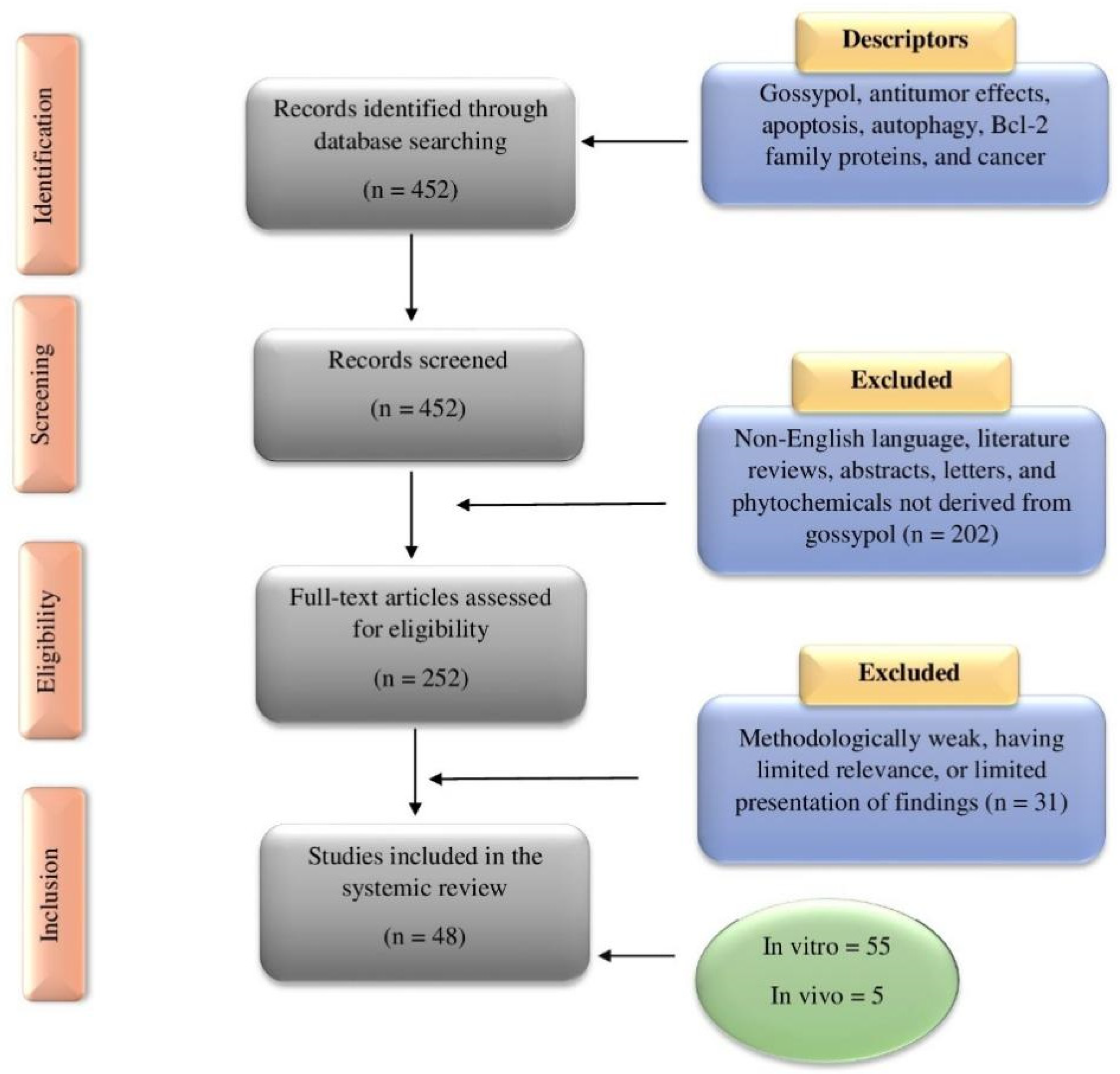
6. Methodology for Literature Search and Selection of Anticancer Studies
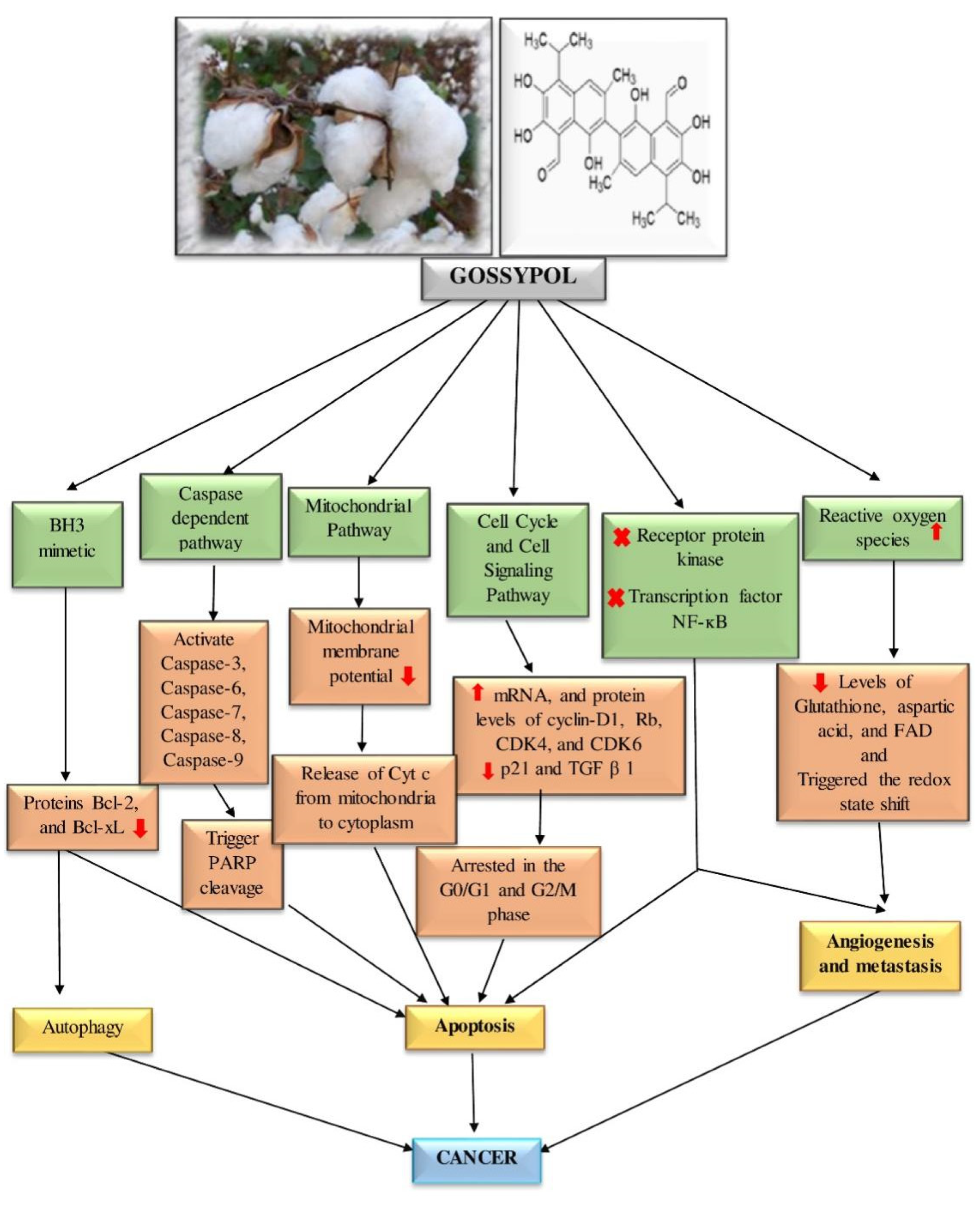
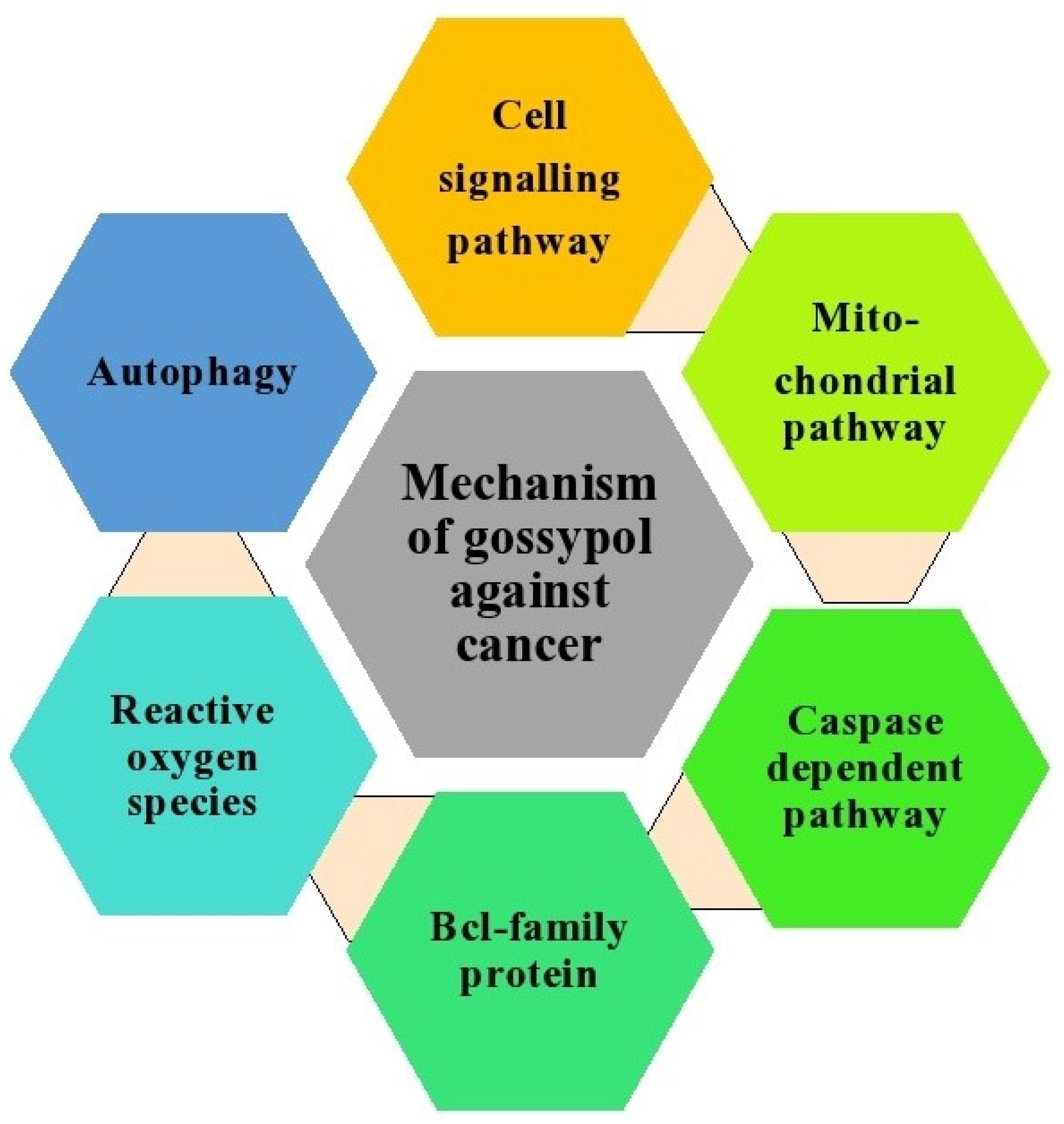
7. Anticancer Activities of Gossypol
7.1. Adrenal Cancer
7.2. Bladder Cancer
7.3. Breast Cancer
7.4. Colorectal Cancer
7.5. Gastric Cancer
7.6. Head and Neck Cancers
7.7. Lung Cancer
7.8. Multiple Myeloma
7.9. Prostate Cancer
8. Anticancer Activities of Gossypol Derivatives
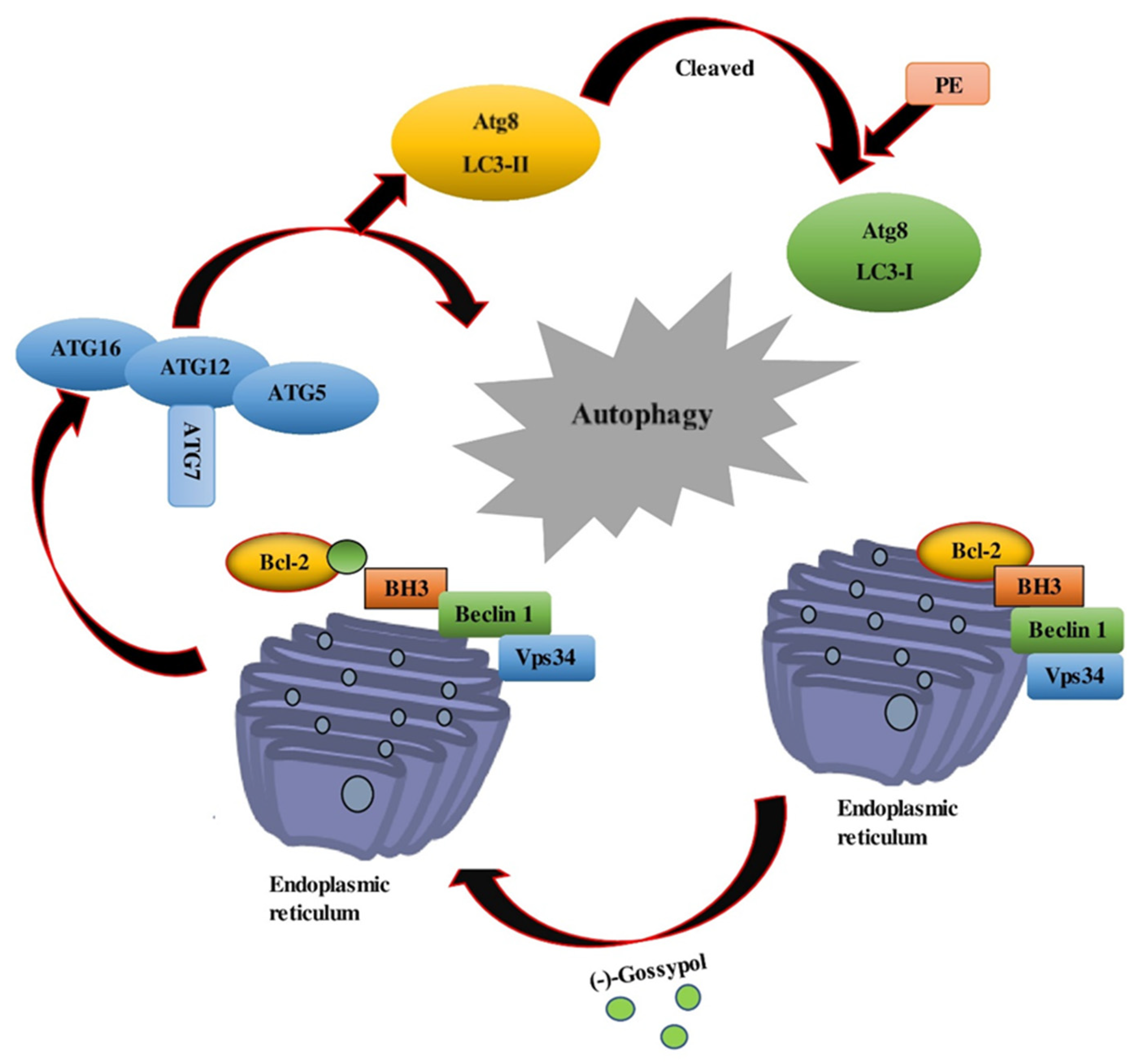
8.1. Apogossypol and ApoG2
8.2. Gossypolone
8.3. Gossypol Schiff Base
8.4. Miscellaneous Gossypol Derivatives
9. Synergistic Studies of Gossypol against Cancer Cells
10. Clinical Studies
11. Future Perspectives
12. Conclusions and Current Challenges/Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| ACC | adrenocortical carcinoma |
| Adr | adriamycin |
| ApoG2 | apogossypolone |
| AUC | area under the curve |
| Bax | Bcl-2-associated X protein |
| Bcl-2 | B-cell lymphoma 2 |
| Bcl-xL | B-cell lymphoma-extra-large |
| Bim | Bcl-2-like protein 11 |
| BNIP3 | BCL2 interacting protein 3 |
| CAD | caspase-activated deoxyribonuclease |
| CDK | cyclin-dependent kinase 4 |
| CHOP | cyclophosphamide-adriamycin-vincristine-prednisolone |
| DR5 | death receptor 5 |
| DT | doubling time |
| ER | Endoplasmic reticulum |
| EGFR | epidermal growth factor receptor |
| GADD45α | growth arrest and DNA damage-inducible 45α |
| HCC | hepatocellular carcinoma |
| HNSCC | head and neck squamous cell carcinoma |
| ICAD | inhibitor of CAD |
| IL | interleukin |
| i.v. | intravenous |
| JNK | Jun N-terminal kinase |
| Mcl-1 | myeloid cell leukemia 1 |
| MM | multiple myeloma |
| NSCLC | non-small cell lung cancer |
| NO | nitric oxide |
| PARP | poly (ADP-ribose) polymerase |
| PCNA | Proliferating cell nuclear antigen |
| p-Rb | phospho-retinoblastoma |
| PRISMA | Preferred Reporting Items for Systematic Reviews and Meta-Analysis |
| PUMA | p53 upregulated modulator of apoptosis |
| RCT | randomized controlled trial |
| ROS | reactive oxygen species |
| TGF-1 | transforming growth factor 1 |
References
- Agarwal, D.K.; Singh, P.; Chakrabarty, M.; Shaikh, A.J.; Gayal, S.G. Cottonseed oil quality, utilization and processing. CICR Technic. Bull. 2003, 1–16. Available online: https://www.cicr.org.in/pdf/cottonseed_oil.pdf (accessed on 24 August 2022).
- Longmore, J. Cotton-seed oil: Its colouring matter and mucilage, and description of a new method of recovering the loss occurring in the refining process. J. Soc. Chem. Ind. 1886, 5, 200–206. [Google Scholar]
- Marchlewski, L. Gossypol, ein Bestandtheil der Baumwoll samen. J. Prakt. Chem. 1899, 60, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Zhong, S.; Leong, J.; Ye, W.; Xu, P.; Lin, S.; Liu, J.; Lin, Y.C. (−)-Gossypol-enriched cottonseed oil inhibits proliferation and adipogenesis of human breast pre-adipocytes. Anticancer Res. 2013, 33, 949–956. [Google Scholar] [PubMed]
- Amini, A.; Kamkar, F. The effects of gossypol on spermatogenesis in NMRI mice. Iran. J. Sci. Technol. 2005, 29, 123–133. [Google Scholar]
- Adams, R.; Geissman, T.A.; Edwards, J.D. Gossypol, A pigment of cottonseed. Chem. Rev. 1960, 60, 555–574. [Google Scholar] [CrossRef]
- Bahraini, Z.; Salari, S.; Sari, M.; Fayazi, J.; Behgar, M. Effect of radiation on chemical composition and protein quality of cottonseed meal. Anim. Sci. J. 2017, 88, 1425–1435. [Google Scholar] [CrossRef]
- Cao, H.; Sethumadhavan, K.; Cao, F.; Wang, T.T.Y. Gossypol decreased cell viability and down—Regulated the expression of a number of genes in human colon cancer cells. Sci. Rep. 2021, 11, 5922. [Google Scholar] [CrossRef]
- Dodou, K. Investigations on gossypol: Past and present developments. Exp. Opin. Investig. Drugs 2005, 14, 1419–1434. [Google Scholar] [CrossRef]
- Zhao, C.; Wu, D. Rapid detection assay for the molecular imprinting of gossypol using a two-layer PMAA/SiO2 bulk structure with a piezoelectric imprinting sensor. Sens. Actuators B Chem. 2013, 181, 104–113. [Google Scholar] [CrossRef]
- Jia, G.; Zhan, Y.; Wu, D.; Meng, Y.; Xu, L. An improved ultrasound-assisted extraction process of Gossypol acetic acid from cottonseed soapstock. AIChE J. 2009, 55, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Morgan, S.E. Gossypol as a toxicant in livestock. Vet. Clin. N. Am. Food. Anim. Pract. 1989, 5, 251–262. [Google Scholar] [CrossRef]
- Prasad, M.R.N.; Diczfalusy, E. Gossypol. Int. J. Androl. 1982, 5, 53–70. [Google Scholar] [CrossRef]
- Qian, S.Z.; Wang, Z.G. Gossypol: A potential antifertility agent for males. Ann. Rev. Pharmacol. Toxicol. 1984, 24, 329–360. [Google Scholar] [CrossRef] [PubMed]
- Randel, R.D.; Chase, C.C.; Wyse, S.J. Effects of Gossy 01 and cottonseed products on reproduction of mammals. J. Anim. Sci. 1992, 70, 1628–1638. [Google Scholar] [CrossRef]
- Coutinho, E.M. Gossypol: A contraceptive for men. Contraception 2002, 65, 259–263. [Google Scholar] [CrossRef]
- Keshmiri-Neghab, H.; Goliaei, B. Therapeutic potential of gossypol: An overview. Pharm. Biol. 2014, 52, 124–128. [Google Scholar] [CrossRef]
- Zeng, Y.; Xu, L.; Wu, D. Natural product Gossypol and its derivatives in precision cancer medicine. Curr. Med. Chem. 2019, 26, 1849–1873. [Google Scholar] [CrossRef] [PubMed]
- Stipanovic, R.; Stoessl, A.; Stothers, J.B.; Altman, D.W.; Bell, A.A.; Heinstein, P. The stereochemistry of the biosynthetic precursor of gossypol. J. Chem. Soc. Chem. Commun. 1986, 2, 100–102. [Google Scholar] [CrossRef]
- Wang, X.; Howell, C.P.; Chen, F.; Yin, J.; Jiang, Y. Chapter 6 Gossypol-A polyphenolic compound from cotton plant. Adv. Food. Nutri. Res. 2009, 58, 215–263. [Google Scholar]
- Khaitbaev, A.K.; Khakberdiev, S.M.; Toshov, K.S. Isolation of Gossypol from the bark of cotton roots. Ann. Rom. Soc. Cell Biol. 2021, 25, 1069–1073. [Google Scholar]
- Kenar, J.A. Reaction chemistry of gossypol and its derivatives. J. Am. Oil Chem. Soc. 2006, 83, 269–302. [Google Scholar] [CrossRef]
- Mohamed, B.A.D. Physiological effects and metabolism of gossypol. Residue Rev. 1976, 61, 125–160. [Google Scholar]
- Beyazit, N.; Çakran, H.S.; Cabir, A.; Akışcan, Y.; Demetgül, C. Synthesis, characterization and antioxidant activity of chitosan Schiff base derivatives bearing (−)-gossypol. Carbohydr. Polym. 2020, 240, 116333. [Google Scholar] [CrossRef] [PubMed]
- Freedman, T.B.; Cao, X.; Oliviera, R.V. Determination of the absolute configuration and solution conformation of gossypol by vibrational circular dichroism. Chirality 2003, 15, 196–200. [Google Scholar] [CrossRef]
- Cass, Q.B.; Oliveira, R.V.; De Pietro, A.C. Determination of Gossypol enantiomer ratio in cotton plants by chiral higher-performance liquid chromatography. J. Agric. Food. Chem. 2004, 52, 5822–5827. [Google Scholar] [CrossRef]
- Cass, Q.B.; Bassi, A.L.; Matlin, S.A. First direct resolution of gossypol enantiomers on a chiral high-performance liquid chromatography phase. Chirality 1999, 11, 46–49. [Google Scholar] [CrossRef]
- Mohammad, R.M.; Wang, S.; Aboukameel, A.; Chen, B.; Wu, X.; Chen, J.; Al-katib, A. Preclinical studies of a nonpeptidic small-molecule inhibitor of Bcl-2 and Bcl-X L [(À) -gossypol] against diffuse large cell lymphoma. Mol. Can. Ther. 2005, 4, 13–22. [Google Scholar] [CrossRef]
- Jaroszewski, J.W.; Strom-Hansen, T.; Hansen, L.L. Optical stability of gossypol. Chirality 1992, 4, 216–221. [Google Scholar] [CrossRef]
- Chamkasem, N. Gossypol analysis in cottonseed oil by HPLC. J. Am. Oil Chem. Soc. 1988, 65, 1601–1604. [Google Scholar] [CrossRef]
- Cass, Q.B.; Oliveira, R.V. Separation of multi-milligram quantities of gossypol enantiomers on polysaccharide-based stationary phases. J. Liq. Chromatogr. Relat. Tec. 2002, 25, 819–829. [Google Scholar] [CrossRef]
- Yin, J.; Jin, L.; Chen, F.; Wang, X.; Kitaygorodskiy, A.; Jiang, Y. Novel O-glycosidic gossypol isomers and their bioactivities. Carbohyd. Res. 2011, 346, 2070–2074. [Google Scholar] [CrossRef] [PubMed]
- Marciniak, B.; Schroeder, G.; Kozubek, H.; Brzezinski, B. Spectroscopic and kinetic studies of the aldehyde—Lactol tautomerisation of gossypol in solution. J. Chem. Soc. Perkin. Trans. 1991, 2, 1359–1362. [Google Scholar] [CrossRef]
- Zhai, G.; Wu, J.U.N.; Zhao, X.; Yu, B.O.; Li, H. A liposomal delivery vehicle for the anticancer agent Gossypol. Anticancer Res. 2008, 28, 2801–2806. [Google Scholar]
- Clément, M.O. Gossypol: A potential promising anticancer agent. SAJP 2017, 6, 320–329. [Google Scholar]
- De Peyster, A.; Wang, Y.Y. Genetic toxicity studies of gossypol. Mutat. Res. Genet. Toxicol. 1993, 297, 293–312. [Google Scholar] [CrossRef]
- Waites, G.M.H.; Wang, C.; Griffin, P.D. Gossypol: Reasons for its failure to be accepted as a safe, reversible male antifertility drug. Int. J. Androl. 1998, 21, 8–12. [Google Scholar] [CrossRef]
- Gadelha, I.C.N.; Fonseca, N.B.S.; Oloris, S.C.S.; Melo, M.M.; Soto-blanco, B. Gossypol Toxicity from Cottonseed Products. Sci. World. J. 2014, 2014, 231635. [Google Scholar] [CrossRef]
- Quintana, P.J.; de Peyster, A.; Klatzke, S.; Park, H.J. Gossypol-induced DNA breaks in rat lymphocytes are secondary to cytotoxicity. Toxicol. Lett. 2000, 117, 85–94. [Google Scholar] [CrossRef]
- Poznak, C.V.; Seidman, A.D.; Reidenberg, M.M.; Moasser, M.M.; Sklarin, N.; Zee, K.V.; Borgen, P.; Gollub, M.; Bacotti, D.; Yao, T.; et al. Oral gossypol in the treatment of patients with refractory metastatic breast cancer: A phase I/II clinical trial. Breast Cancer Res. Treat. 2001, 66, 239–248. [Google Scholar] [CrossRef]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. PLoS. Med. 2009, 6, e1000100. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Ni, Z.; Dai, X.; Wang, B.; Ding, W.; Smith, A.R.; Xu, L.; Wu, D.; He, F.; Lian, J. The novel BH-3 mimetic apogossypolone induces Beclin-1-and ROS-mediated autophagy in human hepatocellular carcinoma cells. Cell Death Dis. 2013, 4, e489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, N.E.; O’Reilly, J.E.; Chang, C.J.G.; Lin, Y.C.; Brueggemeier, R.W. Antiproliferative activity of gossypol and gossypolone on human breast cancer cells. Life Sci. 1995, 57, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Libe, R.; Fratticci, A.; Bertherat, J. Adrenocortical cancer: Pathophysiology and clinical management. Endocr. Relat. Cancer 2007, 14, 13–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzolo, M.; Daffara, F.; Ardito, A.; Zaggia, B.; Basile, V.; Ferrari, L.; Berruti, A. Management of adrenal cancer: A 2013 update. J. Endocrinol. Investig. 2014, 37, 207–217. [Google Scholar] [CrossRef]
- Leblanc, M.L.; Russo, J.; Kudelka, A.P.; Smith, J.A. An In vitro study of inhibitory activity of gossypol, a cottonseed extract, in human carcinoma cell lines. Pharmacol. Res. 2002, 46, 551–555. [Google Scholar] [CrossRef]
- Macoska, J.; Adsule, S.; Tantivejkul, K.; Wang, S.; Pienta, K.; Lee, C. (−)-Gossypol promotes the apoptosis of bladder cancer cells In vitro. Pharmacol. Res. 2008, 58, 323–331. [Google Scholar] [CrossRef]
- Kisim, A.; Atmaca, H.; Cakar, B.; Karabulut, B.; Sezgin, C.; Uzunoglu, S.; Uslu, R.; Karaca, B. Pretreatment with AT-101 enhances tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis of breast cancer cells by inducing death receptors 4 and 5 protein levels. J. Cancer Res. Clin. Oncol. 2012, 138, 1155–1163. [Google Scholar] [CrossRef]
- Ye, W.; Chang, H.; Wang, L.; Huang, Y.; Shu, S.; Sugimoto, Y.; Dowd, M.K.; Wan, P.J.; Lin, Y.C. Induction of apoptosis by (-)-gossypol-enriched cottonseed oil in human breast cancer cells. Int. J. Mol. Med. 2010, 26, 113–119. [Google Scholar]
- Dao, V.; Dowd, M.K.; Martin, M.T.; Gaspard, C.; Mayer, M.; Michelot, R.J. Cytotoxicity of enantiomers of gossypol Schiff’s bases and optical stability of gossypolone. Eur. J. Med. Chem. 2004, 39, 619–624. [Google Scholar] [CrossRef]
- Yan, F.; Cao, X.; Jiang, H.; Zhao, X.; Wang, J.; Lin, Y.; Liu, Q.; Zhang, C.; Liang, B.; Guo, F. A novel water-soluble gossypol derivative increases chemotherapeutic sensitivity and promotes growth inhibition in colon cancer. J. Med. Chem. 2010, 53, 5502–5510. [Google Scholar] [CrossRef] [PubMed]
- Messeha, S.S.; Zarmouh, N.O.; Mendonca, P.; Alwagdani, H.; Cotton, C.; Soliman, K.F.A. Effects of gossypol on apoptosis—Related gene expression in racially distinct triple—Negative breast cancer cells. Oncol. Rep. 2019, 42, 467–478. [Google Scholar] [CrossRef]
- Lan, L.; Appelman, C.; Smith, A.R.; Yu, J.; Larsen, S.; Marquez, R.T.; Xu, L. Natural product (−)-gossypol inhibits colon cancer cell growth by targeting RNA-binding protein Musashi-1. Mol. Oncol. 2015, 9, 1406–1420. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, H.; Guo, R.; Ling, Y.; Wu, X.; Li, B.H.; Roller, P.P.; Wang, S.; Yang, D. Molecular mechanism of gossypol-induced cell growth inhibition and cell death of HT-29 human colon carcinoma cells. Biochem. Pharmacol. 2003, 66, 93–103. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, X.; Mu, S.; Chen, R.; An, Q.; Xia, A.; Wu, D. Inhibitory effect of a new gossypol derivative apogossypolone (ApoG2) on xenograft of human prostate cancer cell line PC-3. J. Med. Coll. PLA 2009, 24, 274–282. [Google Scholar]
- Xin, J.; Zhan, Y.H.; Xia, L.M.; Zhu, H.W.; Nie, Y.Z.; Liang, J.M.; Tian, J. ApoG2 as the most potent gossypol derivatives inhibits cell growth and induces apoptosis on gastric cancer cells. Biomed. Pharmacother. 2013, 67, 88–95. [Google Scholar] [CrossRef]
- Xin, J.; Zhan, Y.; Liu, M.; Hu, H.; Xia, L.; Nie, Y.; Tian, J. ApoG2 induces ER stress-dependent apoptosis in gastric cancer cells In vitro and its real-time evaluation by bioluminescence imaging In vivo. Cancer Lett. 2013, 336, 260–269. [Google Scholar] [CrossRef]
- Mi, J.; Wang, G.; Wang, H.; Sun, X.; Ni, X.; Zhang, X. Synergistic antitumoral activity and induction of apoptosis by novel pan Bcl-2 proteins inhibitor apogossypolone with adriamycin in human hepatocellular carcinoma. Acta Pharmacol. Sin. 2008, 29, 1467–1477. [Google Scholar] [CrossRef] [Green Version]
- Dao, V.; Gaspard, C.; Mayer, M.; Werner, G.H.; Nguyen, S.N.; Michelot, R.J. Synthesis and cytotoxicity of gossypol related compounds. Eur. J. Med. Chem. 2000, 35, 805–813. [Google Scholar] [CrossRef]
- Wolter, K.G.; Wang, S.J.; Henson, B.S.; Wang, S.; Griffith, K.A.; Kumar, B.; D’Silva, N.J. (−)-Gossypol inhibits growth and promotes apoptosis of human head and neck squamous cell carcinoma In vivo. Neoplasia 2006, 8, 163–172. [Google Scholar] [CrossRef]
- Oliver, C.L.; Bauer, J.A.; Wolter, K.G.; Ubell, M.L.; Narayan, A.; Connell, K.M.O.; Fisher, S.G.; Wang, S.; Wu, X. In vitro effects of the BH3 Mimetic, (−)-Gossypol, on head and neck squamous cell carcinoma cells. Clin. Cancer Res. 2004, 10, 7757–7763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Zhu, X.; Zhong, Z.; Sun, J.; Wang, J.; Yang, D. ApoG2, a novel inhibitor of antiapoptotic Bcl-2 family proteins, induces apoptosis and suppresses tumor growth in nasopharyngeal carcinoma xenografts. Int. J. Cancer 2008, 123, 2418–2429. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Sun, J.; Zhu, X.; Yang, D.; Zeng, Y. ApoG2 induces cell cycle arrest of nasopharyngeal carcinoma cells by suppressing the c-Myc signaling pathway. J. Transl. Med. 2009, 7, 2418–2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, D.X.; Uto, T.; Tong, X.; Takeshita, T.; Tanigawa, S.; Imamura, I.; Fujii, M. Involvement of reactive oxygen species-independent mitochondrial pathway in gossypol-induced apoptosis. Arch. Biochem. Biophys. 2004, 428, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Oliver, C.L.; Miranda, M.B.; Shangary, S.; Land, S.; Wang, S.; Johnson, D.E. (−)-Gossypol acts directly on the mitochondria to overcome Bcl-2-and Bcl-XLmediated apoptosis resistance. Mol. Cancer Ther. 2005, 4, 23–31. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, S.; Wang, Y.; Mo, Y.; Xiong, F.; Zhang, S.; Zeng, Z.; Xiong, W.; Li, G.; Chen, H.; et al. Gossypol induces apoptosis of multiple myeloma cells through the JUN-JNK pathway. Am. J. Cancer Res. 2020, 10, 870–883. [Google Scholar]
- Yi, G.; Ni, Z.H.; Zhang, X.; Chen, X.H.; Zou, Z.M. Valproic Acid Enhances the Anti-tumor Effect of (-)-gossypol to Burkitt Lymphoma Namalwa Cells. Biomed. Environ. Sci. 2015, 28, 773–777. [Google Scholar]
- Ni, Z.; Dai, X.; Wang, B.; Ding, W.; Cheng, P.; Xu, L.; He, F. Natural Bcl-2 inhibitor (−)-gossypol induces protective autophagy via reactive oxygen species—High mobility group box 1 pathway in Burkitt lymphoma. Leuk. Lymphoma 2013, 54, 2263–2268. [Google Scholar] [CrossRef]
- Kline, M.P.; Rajkumar, S.V.; Timm, M.M.; Kimlinger, T.K.; Haug, J.L.; Lust, J.A. R-(-)-gossypol (AT-101) activates programmed cell death in multiple myeloma cells. Exp. Hematol. 2008, 36, 568–576. [Google Scholar] [CrossRef] [Green Version]
- Arnold, A.A.; Aboukameel, A.; Chen, J.; Yang, D.; Wang, S.; Al-katib, A.; Mohammad, R.M. Preclinical studies of Apogossypolone: A new nonpeptidic pan small-molecule inhibitor of Bcl-2, Bcl-X L and Mcl-1 proteins in Follicular Small Cleaved Cell Lymphoma model. Mol. Cancer 2008, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Beckham, T.H.; Morris, J.C.; Chen, F.; Gangemi, J.D. Bioactivities of Gossypol, 6-Methoxygossypol, and 6,6′-Dimethoxygossypol. J. Agric. Food. Chem. 2008, 56, 4393–4398. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Lai, E.; Leung, H.; Liu, L.; Yao, X.; Salem, A.F. Gossypol inhibits non-small cell lung cancer cells proliferation by Gossypol inhibits non-small cell lung cancer cells proliferation by targeting EGFRL858R/T790M. Front. Pharmacol. 2018, 9, 728. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Stebbins, J.L.; Kitada, S.; Dash, R.; Placzek, W.; Rega, M.F.; Wu, B.; Cellitti, J.; Zhai, D.; Yang, L.; et al. BI-97C1, an optically pure Apogossypol derivative as pan-active inhibitor of antiapoptotic B-cell lymphoma/leukemia-2 (Bcl-2) family proteins. J. Med. Chem. 2010, 53, 4166–4176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Huang, X.; Mu, S.; An, Q.; Xia, A.; Chen, R.; Wu, D. Inhibition of proliferation of prostate cancer cell line, PC-3, In vitro and In vivo using (−)-gossypol. Asian J. Androl. 2010, 12, 390–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.; Tang, W.; Dai, Y.; Wu, X.; Liu, M.; Ji, Q.; Ji, M.; Pienta, K.; Lawrence, T.; Xu, L. Natural BH3 mimetic (-)-gossypol chemosensitizes human prostate cancer via Bcl-xL inhibition accompanied by increase of Puma and Noxa. Mol. Cancer Ther. 2008, 7, 2192–2202. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Wang, L.; Chang, H.; Ye, W.; Dowd, M.K.; Wan, P.J.; Lin, Y.C. (−)-Gossypol reduces invasiveness in metastatic prostate cancer cells. Anticancer Res. 2009, 29, 2179–2188. [Google Scholar]
- Jiang, J.; Ye, W.; Lin, Y.C. Gossypol inhibits the growth of MAT-LyLu prostate cancer cells by modulation of TGFβ/Akt signaling. Int. J. Mol. Med. 2009, 24, 69–75. [Google Scholar]
- Xu, L.; Yang, D.; Wang, S.; Tang, W.; Liu, M.; Davis, M.; Chen, J.; Rae, J.M.; Lawrence, T.; Lippman, M.E. (−)-Gossypol enhances response to radiation therapy and results in tumor regression of human prostate cancer. Mol. Cancer Ther. 2005, 4, 197–205. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, L.; Chang, H.; Ye, W.; Dowd, M.K.; Wan, P.J.; Lin, Y.C. Molecular mechanisms of (-)-Gossypol-induced apoptosis in human prostate cancer cells. Anticancer Res. 2006, 26, 1925–1934. [Google Scholar]
- Cengiz, E.; Karaca, B.; Kucukzeybek, Y.; Gorumlu, G.; Gul, M.K.; Erten, C.; Atmaca, H.; Uzunoglu, S.; Karabulut, B.; Sanli, U.A.; et al. Overcoming drug resistance in hormone-and drug-refractory prostate cancer cell line, PC-3 by docetaxel and gossypol combination. Mol. Biol. Rep. 2010, 37, 1269–1277. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, H.; Tian, Z.; Griffith, B.N.; Ji, M.; Li, Q.Q. Gossypol induces apoptosis in human PC-3 prostate cancer cells by modulating caspase-dependent and caspase-independent cell death pathways. Life Sci. 2007, 80, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Huang, X.; Hu, X.; Zhan, Y.; An, Q.; Yang, S.; Xia, A.; Yi, J.; Chen, R.; Mu, S.; et al. Apogossypolone, a novel inhibitor of antiapoptotic Bcl-2 family proteins, induces autophagy of PC-3 and LNCaP prostate cancer cells In vitro. Asian J. Androl. 2010, 12, 697–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamat, A.M.; Hahn, N.M.; Efstathiou, J.A.; Lerner, S.P.; Malmström, P.U.; Choi, W.; Guo, C.C.; Lotan, Y.; Kassouf, W. Bladder cancer. Lancet 2016, 388, 2796–2810. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, D.S.; Shipley, W.U.; Feldman, A.S. Bladder cancer. Lancet 2009, 374, 239–249. [Google Scholar] [CrossRef]
- Lenis, A.T.; Lec, P.M.; Chamie, K.; Mshs, M. Bladder Cancer. JAMA 2020, 324, 1980. [Google Scholar] [CrossRef]
- World Health Organization. Breast Cancer. Updated 26 March 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/breast-cancer (accessed on 24 August 2022).
- Harbeck, N.; Gnant, M. Breast cancer. Lancet 2017, 389, 1134–1150. [Google Scholar] [CrossRef]
- Sharma, G.N.; Dave, R.; Sanadya, J.; Sharma, P.; Sharma, K.K. Various types and management of breast cancer: An overview. J. Adv. Pharm. Technol. Res. 2010, 1, 109–126. [Google Scholar]
- Ban, K.A.; Godellas, C.V. Epidemiology of Breast Cancer. Surg. Oncol. Clin. N. Am. 2014, 23, 409–422. [Google Scholar] [CrossRef]
- Sun, Y.S.; Zhao, Z.; Yang, Z.N.; Xu, F.; Lu, H.-J.; Zhu, Z.-Y.; Zhu, H.-P. Risk Factors and Preventions of Breast Cancer. Int. J. Biol. Sci. 2017, 13, 1387–1397. [Google Scholar] [CrossRef] [Green Version]
- Labianca, R.; Beretta, G.D.; Kildani, B.; Milesi, L.; Merlin, F.; Mosconi, S.; Wils, J. Colon Cancer. Crit. Rev. Oncol./Hematol. 2010, 74, 106–133. [Google Scholar] [CrossRef]
- Smyth, E.C.; Nilsson, M.; Grabsch, H.I.; Grieken, N.C.; Lordick, F. Gastric cancer. Lancet 2020, 396, 635–648. [Google Scholar] [CrossRef]
- Van Cutsem, E.; Sagaert, X.; Topal, B.; Haustermans, K.; Prenen, H. Gastric cancer. Lancet 2016, 388, 2654–2664. [Google Scholar] [CrossRef]
- Minna, J.D.; Roth, J.A.; Gazdar, A.F. Focus on lung cancer. Cancer Cell 2002, 1, 49–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raab, M.S.; Podar, K.; Breitkreutz, I.; Richardson, P.G.; Anderson, K.C. Multiple myeloma. Lancet 2009, 374, 324–339. [Google Scholar] [CrossRef] [PubMed]
- Hideshima, T. Advances in biology of multiple myeloma: Clinical applications. Blood 2004, 104, 607–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajkumar, S.; Vincent, K.; Robert, A. Multiple Myeloma: Diagnosis and Treatment. Mayo Clin. Proc. 2005, 80, 1371–1382. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.C.; Alsina, M.; Bensinger, W.; Biermann, J.S.; Chanan-Khan, A.; Cohen, A.D.; Yunus, F. Multiple myeloma. J. Natl. Compr. Cancer Netw. 2009, 7, 908–942. [Google Scholar] [CrossRef]
- Crawford, E.D. Epidemiology of prostate cancer. Urology 2003, 62, 3–12. [Google Scholar] [CrossRef]
- Mohler, J.; Bahnson, R.R.; Boston, B.; Busby, J.E.; D’Amico, A.; Eastham, J.A.; Walsh, P.C. Prostate cancer. J. Natl. Compr. Cancer Netw. 2010, 8, 162–200. [Google Scholar] [CrossRef]
- Pernar, C.H.; Ebot, E.M.; Wilson, K.M.; Mucci, L.A. The epidemiology of prostate cancer. Cold Spring Harb. Lab. Press 2018, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Rawla, P. Epidemiology of prostate cancer. World. J. Oncol. 2019, 10, 63–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Sugimoto, Y.; Liu, S.; Chang, H.L.; Park, K.Y.; Kulp, S.K.; Lin, Y.C. The Inhibitory Effects of Gossypol on Human Prostate Cancer Cells-PC3 are Associated with Transforming Growth Factor Beta1 (TGFβ1) Signal Transduction Pathway. Anticancer Res. 2004, 24, 91–100. [Google Scholar] [PubMed]
- Loberg, R.D.; Mcgregor, N.; Ying, C.; Sargent, E.; Pienta, K.J. In Vivo Evaluation of AT-101 (R-(–)-Gossypol acetic acid) in androgen-independent growth of VCaP prostate cancer cells in combination with surgical castration. Neoplasia 2007, 9, 1030–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, X.; Li, S.; Wei, F.; Huang, J. Apogossypolone induces autophagy and apoptosis in breast cancer MCF-7 cells in vitro and in vivo. Breast Cancer 2014, 21, 223–230. [Google Scholar] [CrossRef]
- Pang, X.; Wu, Y.; Wu, Y.; Lu, B.; Chen, J.; Wang, J.; Yi, Z.; Qu, W.; Liu, M. (−)-Gossypol suppresses the growth of human prostate cancer xenografts via modulating VEGF signaling-mediated angiogenesis. Mol. Cancer Ther. 2011, 10, 795–805. [Google Scholar] [CrossRef] [Green Version]
- Kitada, S.; Kress, C.L.; Krajewska, M.; Jia, L.; Pellecchia, M.; Reed, J.C. Bcl-2 antagonist apogossypol (NSC736630) displays single-agent activity in Bcl-2-transgenic mice and has superior efficacy with less toxicity compared with gossypol (NSC19048). Blood 2008, 111, 3211–3219. [Google Scholar] [CrossRef]
- Zubair, H.; Khan, H.Y.; Ullah, M.; Ahmad, A.; Wu, D.; Hadi, S. Apogossypolone, derivative of gossypol, mobilizes endogenous copper in human peripheral lymphocytes leading to oxidative DNA breakage. Eur. J. Pharm. Sci. 2012, 47, 280–286. [Google Scholar] [CrossRef]
- Zubair, H.; Azim, S.; Khan, H.Y.; Ullah, M.F.; Wu, D.; Singh, A.P.; Hadi, S.M.; Ahmad, A. Mobilization of intracellular copper by gossypol and apogossypolone leads to reactive oxygen species-mediated cell death: Putative anticancer mechanism. Int. J. Mol. Sci. 2016, 17, 973–985. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Jiang, H.; Cao, X.; Zhao, H.; Wang, F.; Cui, Y.; Jiang, B. Chiral gossypol derivatives: Evaluation of their anticancer activity and molecular modeling. Eur. J. Med. Chem. 2009, 44, 3961–3972. [Google Scholar] [CrossRef]
- Dao, V.; Dowd, M.K.; Gaspard, C.; Martin, M.T.; Hémez, J.; Laprévote, O.; Mayer, M.; Michelot, R.J. New thioderivatives of gossypol and gossypolone, as prodrugs of cytotoxic agents. Bioorg. Med. Chem. 2003, 11, 2001–2006. [Google Scholar] [CrossRef]
- Tomoda, K.; Chiang, H.C.; Kozak, K.R.; Kwon, G.S. Injectable (-)-gossypol-loaded Pluronic P85 micelles for cancer chemoradiotherapy. Int. J. Radiat. Biol. 2017, 93, 402–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castilla, C.; Congregado, B.; Chinchón, D.; Torrubia, F.J.; Japón, M.A.; Sáez, C. Bcl-xL is overexpressed in hormone-resistant prostate cancer and promotes survival of LNCaP cells via interaction with proapoptotic Bak. Endocrinol 2006, 147, 4960–4967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, F.Y.; Liem, N.; Xie, C.; Yan, F.L.; Wong, W.C.; Wang, L.; Yong, W.P. Combination therapy with gossypol reveals synergism against gemcitabine resistance in cancer cells with high BCL-2 expression. PLoS ONE 2012, 7, e50786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.; Lai, T.C.; Kwon, G.S. Poly(ethylene glycol)-block-poly(ε-caprolactone) micelles for combination drug delivery: Evaluation of paclitaxel, cyclopamine and gossypol in intraperitoneal xenograft models of ovarian cancer. J. Control. Release 2013, 166, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Renner, O.; Mayer, M.; Leischner, C.; Burkard, M.; Berger, A.; Lauer, U.M.; Venturelli, S.; Bischoff, S.C. Systematic review of Gossypol/AT-101 in cancer clinical trials. Pharmaceuticals 2022, 15, 144. [Google Scholar] [CrossRef]
- Song, S.; Chen, Q.; Li, Y.; Lei, G.; Scott, A.; Huo, L.; Li, C.Y.; Estrella, J.S.; Correa, A.; Pizzi, M.P.; et al. Targeting cancer stem cells with a pan-BCL-2 inhibitor in preclinical and clinical settings in patients with gastroesophageal carcinoma. Gut 2021, 70, 2238–2248. [Google Scholar] [CrossRef]
- Liu, G.; Kelly, W.K.; Wilding, G.; Leopold, L.; Brill, K.; Somer, B. An open-label, multicenter, phase I/II study of single-agent AT-101 in men with castrate-resistant prostate cancer. Clin. Cancer Res. 2009, 15, 3172–3176. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Yin, J.; Shah, M.H.; Menefee, M.E.; Bible, K.C.; Reidy-Lagunes, D.; Kane, M.A.; Quinn, D.I.; Gandara, D.R.; Erlichman, C. A phase II study of the orally administered negative enantiomer of gossypol (AT-101), a BH3 mimetic, in patients with advanced adrenal cortical carcinoma. Investig. New Drugs 2019, 37, 755–762. [Google Scholar] [CrossRef]
- Baggstrom, M.Q.; Qi, Y.; Koczywas, M.; Argiris, A.; Johnson, E.A.; Millward, M.J.; Murphy, S.C.; Erlichman, C.; Rudin, C.M.; Govindan, R. A phase II study of AT-101 (Gossypol) in chemotherapy-sensitive recurrent extensive-stage small cell lung cancer. J. Thorac. Oncol. 2011, 6, 1757–1760. [Google Scholar] [CrossRef] [Green Version]
- Bushunow, P.; Reidenberg, M.M.; Wasenko, J.; Winfield, J.; Lorenzo, B.; Lemke, S.; Himpler, B.; Corona, R.; Coyle, T. Gossypol treatment of recurrent adult malignant gliomas. J. Neuro-Oncol. 1999, 43, 79–86. [Google Scholar] [CrossRef]
- Flack, M.R.; Pyle, R.G.; Mullen, N.M.; Lorenzo, B.; Wu, Y.W.; Knazek, R.A.; Nisula, B.C.; Reidenberg, M.M. Oral Gossypol in the treatment of metastatic adrenal cancer. J. Clin. Endocrinol. Metab. 1993, 76, 1019–1024. [Google Scholar] [PubMed]
- Stein, R.C.; Joseph, A.E.A.; Matlin, S.A.; Cunningham, D.C.; Ford, H.T.; Coombes, R.C. A preliminary clinical study of gossypol in advanced human cancer. Cancer Chemother. Pharmacol. 1992, 30, 480–482. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Location of Gossypol in the Plant | Gossypol Enantiomer in Excess |
|---|---|---|
| Gossypium hirsutum | Seeds and roots | (+) |
| Gossypium barbadense | Seeds | (−) |
| Gossypium aboreum | Seeds, stem and roots | (+) |
| Gossypium herbaceum | Seeds and stem | (+) |
| Gossypium mustelinum | Seeds | (+) |
| Thespesia populnea | Wood, leaves and flowers | (+) |
| Gossypol Derivatives | Structure |
|---|---|
| Gossypol |  |
| Apogossypolone |  |
| Apogossypol |  |
| Gossypolone |  |
| 6-Aminopenicillanic acid sodium gossypolone |  |
| BI-97C1 |  |
| Compound Tested | Cell Lines Used | IC50 (μM) (Duration) | References |
|---|---|---|---|
| Adrenal cancer | |||
| Gossypol | SW-13, H295r (human adrenocortical carcinoma) | 1.3–2.9 | [46] |
| Apogossypol hexaacetate | H295r, SW-13 (human adrenocortical carcinoma) | 5.2–6.8 | [46] |
| Bladder cancer | |||
| Gossypol | UM-UC2 and UM-UC-9 (human bladder carcinoma) | 0–20 | [47] |
| Breast cancer | |||
| Gossypol | MDA-MD-231 (human breast carcinoma) | 14.37 | [11] |
| Gossypol | MCF-7, MDA-MD-231 (human breast carcinoma) | 3.99–4.64 (72 h) | [7] |
| Gossypol | MCF-7, MCF-6/ADR, MDA-MD-231 (human breast carcinoma) | None reported | [48] |
| Gossypol | MCF-7 (human breast carcinoma) | None reported | [49] |
| ApoG2 | MCF-7 (human breast carcinoma) | None reported | [39] |
| Gossypolone | MCF-7, MCF-6/ADR, MDA-MD-231 (human breast carcinoma) | None reported | [43] |
| Gossypolone | MCF-7/ADR, MCF-7/WT (human breast carcinoma) | 0.28–0.36 | [50] |
| 6-Aminopenicillanic acid sodium gossypolone | MCF-7, MDA-MB435 (human breast carcinoma) | 29.06–40 | [51] |
| 6-Aminopenicillanic acid sodium gossypolone | 4T1 (mouse mammary carcinoma) | 28.25 | [51] |
| Gossypol-enriched cottonseed oil | MDA-MD-231, MDA-MD-468 (human breast carcinoma) | 21.85–26.39 | [52] |
| Ethylimine of (±)-gossypol | MCF-7/ADR, MCF-7/WT (human breast carcinoma) | 0.40–0.45 | [50] |
| Ethylimine of (±)- gossypolone | MCF-7/ADR, MCF-7/WT (human breast carcinoma) | 0.29–0.35 | [50] |
| Methylimine of (±)-gossypolone | MCF-7/ADR, MCF-7/WT (human breast carcinoma) | 0.14–0.37 | [50] |
| Gastrointestinal and associated cancers | |||
| Gossypol | HCT116 (human colorectal carcinoma) | 8.8 | [53] |
| Gossypol | COLO 225 (human colon carcinoma) | 0.1–100 | [8] |
| Gossypol | HT-29 (human colorectal carcinoma) | 10 | [54] |
| 6-Aminopenicillanic acid sodium gossypolone | HCT116, SW620 (human colorectal carcinoma) | 6.56–17.35 | [51] |
| 6-Aminopenicillanic acid sodium gossypolone | CT26 (mouse colon carcinoma) | 6.59 | [51] |
| Gossypol | M85 (human gastric carcinoma) | 18.4–39.7 | [55] |
| ApoG2 | MKN28, MKN45, AGS (human gastric carcinoma) | 5.6–11.37 (72 h) | [56] |
| ApoG2 | SGC-7901 (human gastric carcinoma) | 18.7 | [57] |
| ApoG2 | SMMC-7721 (human hepatocellular carcinoma) | 17.29–30.63 | [58] |
| ApoG2 | HepG2, Hep3B (human hepatocellular carcinoma) | None reported | [42] |
| Gynecologic cancers | |||
| Gossypol | SKOV-3 (human ovarian carcinoma) | 5.7 | [46] |
| Gossypol | KB (human cervical carcinoma) | 5.7 | [59] |
| Gossypol | RL95-2 (human endometrial carcinoma) | 3.4 | [46] |
| Gossypol | HeLa (human cervical carcinoma) | 17.8–31.3 | [55] |
| Apogossypol hexaacetate | Skov-3 (human ovarian carcinoma) | 9.0 | [46] |
| Apogossypol hexaacetate | RL95-2 (human endometrial carcinoma) | 7.3 | [46] |
| Gossypolone | KB (human cervical carcinoma) | 0.45 | [50] |
| Gossypolone | KB (human cervical carcinoma) | 2.4 | [59] |
| Ethylimine of (±)-gossypol | KB (human cervical carcinoma) | 1.00 | [50] |
| Ethylimine of (±)- gossypolone | KB (human cervical carcinoma) | 0.55 | [50] |
| Methylimine of (±)-gossypolone | KB (human cervical carcinoma) | 0.50 | [50] |
| Head and neck cancers | |||
| Gossypol | Um-SCC-1, Um-SCC-17b, UM-SCC-5PT, UM-SCC-10BPT (human HNSCC) | 3–15 | [60] |
| Gossypol | UM-SCC-12, UM-SCC-23, UM-SCC-1, UM-SCC-6, UM-SCC-14A, UM-SCC-74B, UM-SCC-81B, UM-SCC-22A, UM-SCC-17B, UM-SCC-25 (human HNSCC) | 3.75–6.85 | [61] |
| ApoG2 | C666-1, CNE-1, CNE-2, HONE-1 (human nasopharyngeal carcinoma) | 0.908–66.01 | [62] |
| ApoG2 | CNE-2, HONE-1 (human nasopharyngeal carcinoma) | None reported | [63] |
| Hematologic cancers | |||
| Gossypol | HL60 (human acute myeloid leukemia) | 2.0–8.1 (24–48 h) | [64] |
| Gossypol | Vector/Jurkat, Bcl-2/Jurkat, Bcl-xL/Jurkat (human T cell leukemia) | 4.3–26.6 | [65] |
| Gossypol | U266, ARK, MM1, H929, OC1-MY5, OPM2, ARP-1, KMS11, CAG (human multiple myeloma) | < 6 | [66] |
| Gossypol | Namalwa (human Burkitt lymphoma) | None reported | [67] |
| Gossypol | Namalwa (human Burkitt lymphoma) | None reported | [68] |
| Gossypol | KAS-6/1, MM1.S, OPM-1, OPM-2, U266 (human multiple myeloma) | 2.5–7.5 | [69] |
| ApoG2 | WSU-FSCCL (human B cell lymphoma) | 0.109 | [70] |
| Lung cancer | |||
| Gossypol | H1975 (human lung adenocarcinoma) | 10.05–11.73 | [71] |
| Gossypol | H1975 (human lung adenocarcinoma) | 34.17–36.35 | [72] |
| 6-Aminopenicillanic acid sodium gossypolone | A549 (human lung adenocarcinoma) | 8.77 | [51] |
| BI-97C1 | H460 (human large cell lung carcinoma) | 0.33–0.51 | [73] |
| Neural cancers | |||
| Gossypol | U87 (human malignant glioma) | 30.2–59.6 | [55] |
| Prostate cancer | |||
| Gossypol | PC-3 (human prostate carcinoma) | 9.096 | [11] |
| Gossypol | PC-3 (human prostate carcinoma) | 4.74 | [74] |
| Gossypol | PC-3, LNCaP (human prostate carcinoma) | 2.8–3.5 | [75] |
| Gossypol | MAT-lylu (rat prostate carcinoma) | 14.5 | [76] |
| Gossypol | MAT-lylu (rat prostate carcinoma) | None reported | [77] |
| Gossypol | PC-3 (human prostate carcinoma) | None reported | [78] |
| Gossypol | DU-145 (human prostate carcinoma) | None reported | [79] |
| Gossypol | PC-3 (human prostate carcinoma) | 10 (24–72 h) | [80] |
| Gossypol | PC-3 (human prostate carcinoma) | None reported | [81] |
| ApoG2 | PC-3, LNCaP (human prostate carcinoma) | None reported | [82] |
| BI-97C1 | PC-3 (human prostate carcinoma) | 0.11–0.15 | [73] |
| 6-Aminopenicillanic acid sodium gossypolone | PC-3 (human prostate carcinoma) | 29.46 | [51] |
| Skin cancer | |||
| 6-Aminopenicillanic acid sodium gossypolone | B16-F10 (mouse melanoma) | 11.05 | [51] |
| Thyroid cancer | |||
| Apogossypol hexaacetate | TT (human thyroid carcinoma) | 18.9 | [46] |
| Compound Tested | Animal Tumor Models | Anticancer Effects | Mechanisms | Dose (Route) | Duration | References |
|---|---|---|---|---|---|---|
| Breast cancer | ||||||
| Apogossypolone | Athymic nude female mice xenografted with MCF-7 breast carcinoma cells | Inhibited tumor growth | ↑Apoptosis; ↑autophagy | 120 mg/kg/day (i.p.) | 4 weeks | [105] |
| Gastrointestinal and associated cancers | ||||||
| Combination of apogossypolone and adriamycin | BALB/c nu/nu mice xenografted with SMMC-7721 hepatocellular carcinoma cells | Inhibited tumor growth | ↑Apoptosis; ↑DNA fragmentation | 100 or 200 mg/kg/day (intragastric) ± 5.5 mg/kg/week adriamycin (i.v.) | 28 days | [58] |
| Head and neck cancers | ||||||
| Apogossypolone | Athymic nude (nu/nu) mice xenografted with CNE-1 or CNE-2 nasopharyngeal carcinoma cells | Inhibited tumor growth and angiogenesis; enhanced antitumor activity of cisplatin (CNE-2 cells only) | ↑Apoptosis; ↓CD31 | 200 mg/kg/day (intragastric) ± 3 mg/kg; every 2 days (i.p.)cisplatin | [62] | |
| Hematologic cancers | ||||||
| ApoG2 | Female ICR SCID mice xenografted with WSU-FSCCL B cell lymphoma cells | Inhibited tumor growth | ↑Caspase-3; ↑caspase-8; ↑caspase-9; ↑PARP; ↑AIF | 25 mg/kg/day (i.p. or i.v.) | 5 days | [70] |
| Prostate cancer | ||||||
| (−)-Gossypol | Male BALB/c nude mice xenografted with PC-3 prostate carcinoma cells | Inhibited tumor growth and angiogenesis | ↓VEGF | 15 mg/kg/day (intratumoral) | 50 days | [106] |
| (−)-Gossypol | Male BALB/c nude mice xenografted with PC-3 prostate carcinoma cells | Inhibited tumor growth | ↓Bcl-2; ↑caspase-3; ↑caspase-8; ↓CD31; ↓PCNA | 2.5–10 mg/kg/day (i.p.) | 7 days | [74] |
| AT-101(R-(–)-gossypol acetic acid | SCID mice xenografted with VCaP prostate carcinoma cells | Inhibited tumor growth | ↑Apoptosis; ↑Bcl-2; ↑androgen receptor expression | 15 mg/kg; 5 days/week (p.o.) | 6 weeks | [104] |
| Clinical Trial (Date) | Trial Design | Cancer Type (Number of Patients) | Dose, Route, Frequency (Duration) | Adverse Events | Primary Outcome | References |
|---|---|---|---|---|---|---|
| NCT00848016 (2019) | Phase II | Adrenocortical carcinoma (n = 29) | 20 mg AT-101 PO QD (21 days/month × 80 cycles) | Elevated cardiac troponin, hypokalemia, GI upset, elevated AST/ALT and fatigue | PD: 27; PR: 0; SD: 8 Median duration of 3.8 months; median time of progression 1.9 months; mOS: 8.5 months | [119] |
| NCT00773955 (2011) | Phase II | Small cell lung carcinoma (n = 14) | 20 mg AT-101 PO QD (21 days/month × 6 cycles) | GI upset, exhaustion, anorexia and hematologic abnormalities | OR: 0; SD: 3 Median time of progression 1.7 months; mOS: 8.5 months | [120] |
| NCT00286806 (2009) | Phase I/II | Castrate-resistant prostate carcinoma (n = 23) | 30 mg AT-101 (21 days/month) chemotherapy naive patients, ≥eight weeks of therapy | GI upset, elevated AST/ALT and small intestinal obstruction | Modest single-agent activity; treatment was generally well tolerated | [118] |
| (2001) | Phase I/II | Breast carcinoma (n = 20) | 30–50 mg AT-101 QD; Pre-treatment with doxorubicin and paclitaxel | GI upset, fatigue and dysgeusia | MR: 1; SD: 2 Blood gossypol levels are 10-fold lower than in vitro levels; No clear correlation between plasma drug levels and gossypol dose | [40] |
| (1999) | Phase II | Radiation-resistant glial carcinoma (n = 27) | 10 mg racemic gossypol acetic acid p.o. QD or BID | Mild toxicity | PR: 2; SD: 4; PD: 21 No difference in plasma levels between responders and non-responders | [121] |
| (1993) | Phase I | Adrenal carcinoma (n = 21) | 30–70 mg racemic gossypol mitotane, and suramine | No serious adverse events | PR: 3 (50% reduction in tumor volume); MR: 1; PD: 13 No significant decrease in steroid secretion | [122] |
| (1992) | Phase I | Human carcinoma (n = 34) | 30–180 mg gossypol acetic acid; followed by 30 mg twice weekly, QD, or BID | No serious adverse events | SD: 3; PD: 20 No association between serum drug levels and gossypol dose | [123] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pal, D.; Sahu, P.; Sethi, G.; Wallace, C.E.; Bishayee, A. Gossypol and Its Natural Derivatives: Multitargeted Phytochemicals as Potential Drug Candidates for Oncologic Diseases. Pharmaceutics 2022, 14, 2624. https://doi.org/10.3390/pharmaceutics14122624
Pal D, Sahu P, Sethi G, Wallace CE, Bishayee A. Gossypol and Its Natural Derivatives: Multitargeted Phytochemicals as Potential Drug Candidates for Oncologic Diseases. Pharmaceutics. 2022; 14(12):2624. https://doi.org/10.3390/pharmaceutics14122624
Chicago/Turabian StylePal, Dilipkumar, Pooja Sahu, Gautam Sethi, Carly E. Wallace, and Anupam Bishayee. 2022. "Gossypol and Its Natural Derivatives: Multitargeted Phytochemicals as Potential Drug Candidates for Oncologic Diseases" Pharmaceutics 14, no. 12: 2624. https://doi.org/10.3390/pharmaceutics14122624
APA StylePal, D., Sahu, P., Sethi, G., Wallace, C. E., & Bishayee, A. (2022). Gossypol and Its Natural Derivatives: Multitargeted Phytochemicals as Potential Drug Candidates for Oncologic Diseases. Pharmaceutics, 14(12), 2624. https://doi.org/10.3390/pharmaceutics14122624