
Rapid and Specific Action of Methylene Blue against Plasmodium Transmission Stages

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection for Ex Vivo Study
2.2. Mosquito Species
- Anopheles gambiae s. s. (Kisumu strain) were reared at the Laboratory of Entomology at CERMEL in Lambaréné (Gabon). Adult mosquitoes were kept at standard insectary conditions (26 ± 0.5 °C and 80% ± 10 relative humidity) in heating cabinets with a 12:12 h (h) light and dark cycle. They were fed with a 10% sugar solution. Larvae were hatched in room temperature water and fed with a small amount of Vitakraft® Premium VITA Flake-Mix fish provided daily;
- Anopheles stephensi were reared at the Insectary at the Center for Immunology and Infectious Diseases (CIMI) in Paris under similar conditions.
2.3. Parasites
- Plasmodium berghei ANKA GFP, a parasite line constitutively expressing GFP as previously described [26];
- Plasmodium yoelii (17XNL strain), a parasite line expressing both GFP and Luciferase reporters (GFP-luc) as previously described [26];
- Plasmodium falciparum (NF54 strain) sporozoites were obtained from infected salivary glands of A. stephensi 14–21 days after an infective blood meal (Department of Medical Microbiology, Radboud University Medical Center, Nijmegen, The Netherlands).
2.4. Methylene Blue
2.5. Ex Vivo Transmission Blocking Assessment of MB
2.6. Assessment of Anti-Mosquito Stage Activity of MB by Secondary In Vivo Feeding
2.7. In Vitro Sporozoite Invasion Assay
2.8. In Vitro Liver Stage Development Assay
2.9. In Vivo Sporozoite Invasion/Development Assay
2.10. Statistical Analysis
3. Results
3.1. Short-Term Exposure of Fresh P. falciparum Isolates to MB Efficiently Blocks Mosquito Infections
3.2. Limited Effect of MB on Plasmodium berghei Mosquito Stage Development
3.3. Pre-Exposure of Sporozoites with MB Inhibits Hepatocyte Invasion
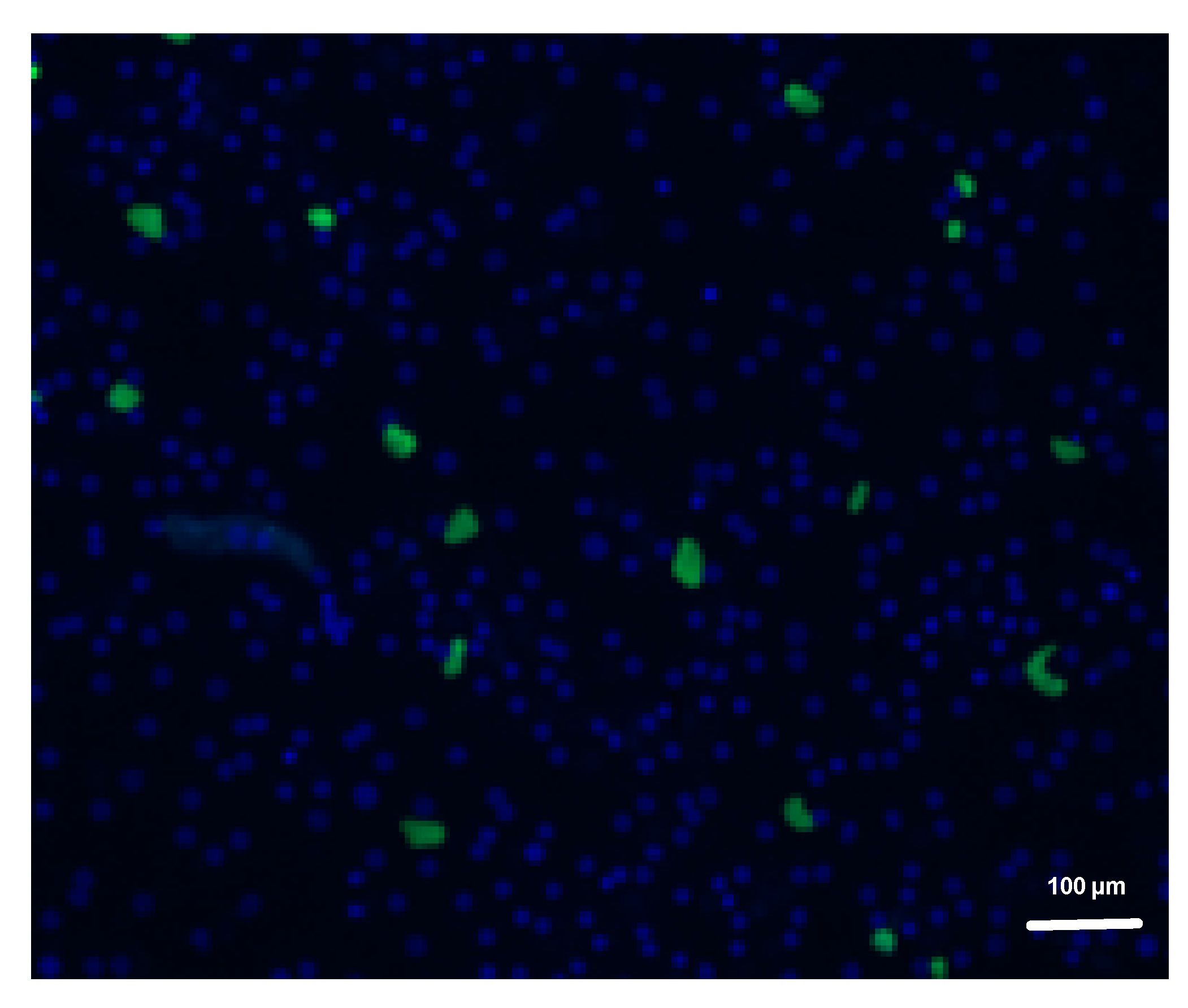
3.4. MB Inhibits Early In Vitro Liver Stage Development
3.5. Reduction of In Vivo Liver Stage Development by MB
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Age | T (°C) | Parasitaemia | Gametocytaemia | Infection Rate in Control (%) | Mean Oocyst |
|---|---|---|---|---|---|---|
| CML 0570 | 8 | 36.8 | 589 | 288 | 55 | 1.15 |
| CML 0897 | 3 | 36.3 | 1072 | 716 | 60 | 10.8 |
| CML 1036 | 10 | 36.5 | 0 | 212 | 67 | 3.14 |
| CML 1016 | 20 | 36.6 | 0 | 637 | 80 | 5.55 |
| CML 1099 | 5 | 37.1 | 0 | 208 | 40 | 4.15 |

Appendix B
| DayPost-Infection | Methylene Blue Dose (mg/kg) | Ni (n) | Mean ± SEM | Range | Prevalence (%) |
|---|---|---|---|---|---|
| DPI 3 | 0 | 43 | 97 ± 15.5 | 0–388 | 84 |
| 50 | 41 | 74 ± 14.8 | 0–345 | 71 | |
| 100 | 41 | 27.17 ± 7.7 | 0–189 | 54 | |
| DPI 6 | 0 | 24 | 737.7 ± 90.9 | 17–1684 | 91 |
| 50 | 28 | 476 ± 111.6 | 49–1821 | 75 | |
| 100 | 26 | 508.5 ± 89.1 | 1–1879 | 76 |
| Date of the MB Feeding | Methylene Blue Dose (mg/kg) | Ni (n) | Arithmetic Mean ± SE |
|---|---|---|---|
| DPI 6 | 0 | 29 | 63,065 ± 23,007 |
| 50 | 30 | 48,522 ± 4666 | |
| 100 | 14 | 35,643 ± 10,404 | |
| DP 15 | 0 | 30 | 37,325 ± 3653 |
| 50 | 31 | 35,747 ± 25,444 |


References
- World Health Organization. World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Uwimana, A.; Legrand, E.; Stokes, B.H.; Ndikumana, J.M.; Warsame, M.; Umulisa, N.; Ngamije, D.; Munyaneza, T.; Mazarati, J.B.; Munguti, K.; et al. Emergence and clonal expansion of in vitro artemisinin-resistant Plasmodium falciparum kelch13 R561H mutant parasites in Rwanda. Nat. Med. 2020, 26, 1602–1608. [Google Scholar] [CrossRef]
- Balikagala, B.; Fukuda, N.; Ikeda, M.; Katuro, O.T.; Tachibana, S.I.; Yamauchi, M.; Opio, W.; Emoto, S.; Anywar, D.A.; Kimura, E.; et al. Evidence of Artemisinin-Resistant Malaria in Africa. N. Engl. J. Med. 2021, 385, 1163–1171. [Google Scholar] [CrossRef] [PubMed]
- Maiga, F.O.; Wele, M.; Toure, S.M.; Keita, M.; Tangara, C.O.; Refeld, R.R.; Thiero, O.; Kayentao, K.; Diakite, M.; Dara, A.; et al. Artemisinin-based combination therapy for uncomplicated Plasmodium falciparum malaria in Mali: A systematic review and meta-analysis. Malar. J. 2021, 20, 356. [Google Scholar] [CrossRef] [PubMed]
- Toure, O.A.; Assi, S.B.; Kiki-Barro, P.M.C.; Yavo, W.; Abba, T.; Tiacoh, L.N.; Konate, A.A.; Angora, E.K.; Bedia, V.A.; Menan, H.; et al. Efficacy and safety of artesuante-amodiaquine and artemether lumefantrine, the first line malaria treatment in six sentinel’s sites of Cote d’Ivoire, West Africa. Ann. Parasitol. 2020, 66, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Sawa, P.; Shekalaghe, S.A.; Drakeley, C.J.; Sutherland, C.J.; Mweresa, C.K.; Baidjoe, A.Y.; Manjurano, A.; Kavishe, R.A.; Beshir, K.B.; Yussuf, R.U.; et al. Malaria transmission after artemether-lumefantrine and dihydroartemisinin-piperaquine: A randomized trial. J. Infect. Dis. 2013, 207, 1637–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adjalley, S.H.; Johnston, G.L.; Li, T.; Eastman, R.T.; Ekland, E.H.; Eappen, A.G.; Richman, A.; Sim, B.K.; Lee, M.C.; Hoffman, S.L.; et al. Quantitative assessment of Plasmodium falciparum sexual development reveals potent transmission-blocking activity by methylene blue. Proc. Natl. Acad. Sci. USA 2011, 108, E1214–E1223. [Google Scholar] [CrossRef] [Green Version]
- Huho, B.J.; Killeen, G.F.; Ferguson, H.M.; Tami, A.; Lengeler, C.; Charlwood, J.D.; Kihonda, A.; Kihonda, J.; Kachur, S.P.; Smith, T.A.; et al. Artemisinin-based combination therapy does not measurably reduce human infectiousness to vectors in a setting of intense malaria transmission. Malar. J. 2012, 11, 118. [Google Scholar] [CrossRef] [Green Version]
- Eziefula, A.C.; Pett, H.; Grignard, L.; Opus, S.; Kiggundu, M.; Kamya, M.R.; Yeung, S.; Staedke, S.G.; Bousema, T.; Drakeley, C. Glucose-6-phosphate dehydrogenase status and risk of hemolysis in Plasmodium falciparum-infected African children receiving single-dose primaquine. Antimicrob. Agents Chemother. 2014, 58, 4971–4973. [Google Scholar] [CrossRef] [Green Version]
- Dicko, A.; Roh, M.E.; Diawara, H.; Mahamar, A.; Soumare, H.M.; Lanke, K.; Bradley, J.; Sanogo, K.; Kone, D.T.; Diarra, K.; et al. Efficacy and safety of primaquine and methylene blue for prevention of Plasmodium falciparum transmission in Mali: A phase 2, single-blind, randomised controlled trial. Lancet Infect. Dis. 2018, 18, 627–639. [Google Scholar] [CrossRef] [Green Version]
- Updated WHO Policy Recommendation: Single Dose Primaquine as a Gametocytocide in Plasmodium Falciparum Malaria; WHO: Geneva, Switzerland, October 2012.
- Guidelines for the Treatment of Malaria; World Health Organization: Geneva, Switzerland, 2015.
- Guttmann, P.E.P. Ueber die Wirkung des methylenblau bei Malaria. Berlin Klin. Wochenschr. 1891, 28, 953–956. [Google Scholar]
- Lu, G.; Nagbanshi, M.; Goldau, N.; Mendes Jorge, M.; Meissner, P.; Jahn, A.; Mockenhaupt, F.P.; Muller, O. Efficacy and safety of methylene blue in the treatment of malaria: A systematic review. BMC Med. 2018, 16, 59. [Google Scholar] [CrossRef]
- Anh, C.X.; Chavchich, M.; Birrell, G.W.; Van Breda, K.; Travers, T.; Rowcliffe, K.; Lord, A.R.; Shanks, G.D.; Edstein, M.D. Pharmacokinetics and Ex Vivo Antimalarial Activity of Artesunate-Amodiaquine plus Methylene Blue in Healthy Volunteers. Antimicrob. Agents Chemother. 2020, 64, e01441-19. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, R.H.; Coulibaly, B.; Stich, A.; Scheiwein, M.; Merkle, H.; Eubel, J.; Becker, K.; Becher, H.; Muller, O.; Zich, T.; et al. Methylene blue as an antimalarial agent. Redox Rep. 2003, 8, 272–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, O.; Mockenhaupt, F.P.; Marks, B.; Meissner, P.; Coulibaly, B.; Kuhnert, R.; Buchner, H.; Schirmer, R.H.; Walter-Sack, I.; Sie, A.; et al. Haemolysis risk in methylene blue treatment of G6PD-sufficient and G6PD-deficient West-African children with uncomplicated falciparum malaria: A synopsis of four RCTs. Pharmacoepidemiol. Drug. Saf. 2013, 22, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Bistas, E.; Sanghavi, D. Methylene Blue. In StatPearls; Treasure Island (FL): Tampa, FL, USA, 2022. [Google Scholar]
- Buchholz, K.; Schirmer, R.H.; Eubel, J.K.; Akoachere, M.B.; Dandekar, T.; Becker, K.; Gromer, S. Interactions of methylene blue with human disulfide reductases and their orthologues from Plasmodium falciparum. Antimicrob. Agents Chemother. 2008, 52, 183–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vennerstrom, J.L.; Makler, M.T.; Angerhofer, C.K.; Williams, J.A. Antimalarial dyes revisited: Xanthenes, azines, oxazines, and thiazines. Antimicrob. Agents Chemother. 1995, 39, 2671–2677. [Google Scholar] [CrossRef] [Green Version]
- Bosson-Vanga, H.; Franetich, J.F.; Soulard, V.; Sossau, D.; Tefit, M.; Kane, B.; Vaillant, J.C.; Borrmann, S.; Muller, O.; Dereuddre-Bosquet, N.; et al. Differential activity of methylene blue against erythrocytic and hepatic stages of Plasmodium. Malar. J. 2018, 17, 143. [Google Scholar] [CrossRef]
- Wadi, I.; Pillai, C.R.; Anvikar, A.R.; Sinha, A.; Nath, M.; Valecha, N. Methylene blue induced morphological deformations in Plasmodium falciparum gametocytes: Implications for transmission-blocking. Malar. J. 2018, 17, 11. [Google Scholar] [CrossRef] [Green Version]
- Bradley, J.; Soumare, H.M.; Mahamar, A.; Diawara, H.; Roh, M.; Delves, M.; Drakeley, C.; Churcher, T.S.; Dicko, A.; Gosling, R.; et al. Transmission-blocking Effects of Primaquine and Methylene Blue Suggest Plasmodium falciparum Gametocyte Sterilization Rather Than Effects on Sex Ratio. Clin. Infect. Dis. 2019, 69, 1436–1439. [Google Scholar] [CrossRef] [Green Version]
- Mendes Jorge, M.; Ouermi, L.; Meissner, P.; Compaore, G.; Coulibaly, B.; Nebie, E.; Krisam, J.; Klose, C.; Kieser, M.; Jahn, A.; et al. Safety and efficacy of artesunate-amodiaquine combined with either methylene blue or primaquine in children with falciparum malaria in Burkina Faso: A randomized controlled trial. PLoS ONE 2019, 14, e0222993. [Google Scholar] [CrossRef]
- Grobusch, M.P.; Lell, B.; Schwarz, N.G.; Gabor, J.; Dornemann, J.; Potschke, M.; Oyakhirome, S.; Kiessling, G.C.; Necek, M.; Langin, M.U.; et al. Intermittent preventive treatment against malaria in infants in Gabon--a randomized, double-blind, placebo-controlled trial. J. Infect. Dis. 2007, 196, 1595–1602. [Google Scholar] [CrossRef] [PubMed]
- Manzoni, G.; Briquet, S.; Risco-Castillo, V.; Gaultier, C.; Topcu, S.; Ivanescu, M.L.; Franetich, J.F.; Hoareau-Coudert, B.; Mazier, D.; Silvie, O. A rapid and robust selection procedure for generating drug-selectable marker-free recombinant malaria parasites. Sci. Rep. 2014, 4, 4760. [Google Scholar] [CrossRef] [Green Version]
- Delves, M.J.; Sinden, R.E. A semi-automated method for counting fluorescent malaria oocysts increases the throughput of transmission blocking studies. Malar. J. 2010, 9, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Churcher, T.S.; Blagborough, A.M.; Delves, M.; Ramakrishnan, C.; Kapulu, M.C.; Williams, A.R.; Biswas, S.; Da, D.F.; Cohuet, A.; Sinden, R.E. Measuring the blockade of malaria transmission—An analysis of the Standard Membrane Feeding Assay. Int. J. Parasitol. 2012, 42, 1037–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Evidence Review Group: The Safety and Effectiveness of Single Dose Primaquine as a P. Falciparum Gametocytocide; Pullman Hotel: Bangkok, Thailand, 13–15 August 2012.
- Pastrana-Mena, R.; Dinglasan, R.R.; Franke-Fayard, B.; Vega-Rodriguez, J.; Fuentes-Caraballo, M.; Baerga-Ortiz, A.; Coppens, I.; Jacobs-Lorena, M.; Janse, C.J.; Serrano, A.E. Glutathione reductase-null malaria parasites have normal blood stage growth but arrest during development in the mosquito. J. Biol. Chem. 2010, 285, 27045–27056. [Google Scholar] [CrossRef] [Green Version]
- Douglas, R.G.; Reinig, M.; Neale, M.; Frischknecht, F. Screening for potential prophylactics targeting sporozoite motility through the skin. Malar. J. 2018, 17, 319. [Google Scholar] [CrossRef] [Green Version]
- Chaccour, C.J.; Kobylinski, K.C.; Bassat, Q.; Bousema, T.; Drakeley, C.; Alonso, P.; Foy, B.D. Ivermectin to reduce malaria transmission: A research agenda for a promising new tool for elimination. Malar. J. 2013, 12, 153. [Google Scholar] [CrossRef] [Green Version]
- Peter, C.; Hongwan, D.; Kupfer, A.; Lauterburg, B.H. Pharmacokinetics and organ distribution of intravenous and oral methylene blue. Eur. J. Clin. Pharmacol. 2000, 56, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Paton, D.G.; Childs, L.M.; Itoe, M.A.; Holmdahl, I.E.; Buckee, C.O.; Catteruccia, F. Exposing Anopheles mosquitoes to antimalarials blocks Plasmodium parasite transmission. Nature 2019, 567, 239–243. [Google Scholar] [CrossRef]
- Fowler, R.E.; Sinden, R.E.; Pudney, M. Inhibitory activity of the anti-malarial atovaquone (566C80) against ookinetes, oocysts, and sporozoites of Plasmodium berghei. J. Parasitol. 1995, 81, 452–458. [Google Scholar] [CrossRef]
- Kobylinski, K.C.; Ubalee, R.; Ponlawat, A.; Nitatsukprasert, C.; Phasomkulsolsil, S.; Wattanakul, T.; Tarning, J.; Na-Bangchang, K.; McCardle, P.W.; Davidson, S.A.; et al. Ivermectin susceptibility and sporontocidal effect in Greater Mekong Subregion Anopheles. Malar. J. 2017, 16, 280. [Google Scholar] [CrossRef] [PubMed]
- Capela, R.; Cabal, G.G.; Rosenthal, P.J.; Gut, J.; Mota, M.M.; Moreira, R.; Lopes, F.; Prudencio, M. Design and evaluation of primaquine-artemisinin hybrids as a multistage antimalarial strategy. Antimicrob. Agents Chemother. 2011, 55, 4698–4706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitao, R.; Rodriguez, A. Inhibition of Plasmodium sporozoites infection by targeting the host cell. Exp. Parasitol. 2010, 126, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Pinilla, Y.T.; Stefanie, C.P.L.; Vanderson, S.S.; Andrade, F.S.; Melo, G.C.; Orfano, A.S.; Secundino, N.F.C.; Guerra, M.; Lacerda, M.V.G.; Kobylinski, K.C.; et al. Promising approach to reducing Malaria transmission by ivermectin: Sporontocidal effect against Plasmodium vivax in the South American vectors Anopheles aquasalis and Anopheles darlingi. PLoS Negl. Trop. Dis. 2018, 12, e0006221. [Google Scholar] [CrossRef] [PubMed]
- Reprinted with Permission from WHO Global Malaria Programme, Malaria Parasite Counting; Copyright CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2016.






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saison, N.; Franetich, J.-F.; Pinilla, Y.T.; Hoffmann, A.; Boussougou-Sambe, S.T.; Ngossanga, B.; Tefit, M.; Ashraf, K.; Amanzougaghene, N.; Tajeri, S.; et al. Rapid and Specific Action of Methylene Blue against Plasmodium Transmission Stages. Pharmaceutics 2022, 14, 2794. https://doi.org/10.3390/pharmaceutics14122794
Saison N, Franetich J-F, Pinilla YT, Hoffmann A, Boussougou-Sambe ST, Ngossanga B, Tefit M, Ashraf K, Amanzougaghene N, Tajeri S, et al. Rapid and Specific Action of Methylene Blue against Plasmodium Transmission Stages. Pharmaceutics. 2022; 14(12):2794. https://doi.org/10.3390/pharmaceutics14122794
Chicago/Turabian StyleSaison, Nathanaël, Jean-François Franetich, Yudi T. Pinilla, Anton Hoffmann, Stravensky T. Boussougou-Sambe, Barclaye Ngossanga, Maurel Tefit, Kutub Ashraf, Nadia Amanzougaghene, Shahin Tajeri, and et al. 2022. "Rapid and Specific Action of Methylene Blue against Plasmodium Transmission Stages" Pharmaceutics 14, no. 12: 2794. https://doi.org/10.3390/pharmaceutics14122794
APA StyleSaison, N., Franetich, J. -F., Pinilla, Y. T., Hoffmann, A., Boussougou-Sambe, S. T., Ngossanga, B., Tefit, M., Ashraf, K., Amanzougaghene, N., Tajeri, S., Adegnika, A. A., Mazier, D., & Borrmann, S. (2022). Rapid and Specific Action of Methylene Blue against Plasmodium Transmission Stages. Pharmaceutics, 14(12), 2794. https://doi.org/10.3390/pharmaceutics14122794