
Inhaled siRNA Formulations for Respiratory Diseases: From Basic Research to Clinical Application


Abstract
:1. Introduction
2. Respiratory Diseases Treated with Inhaled siRNA Formulations
2.1. Lung Cancer
2.2. Respiratory Infection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Target | Administration | Delivery System | Ref. |
|---|---|---|---|---|
| Influenza | Nucleoprotein | Intranasal administration | Chitosan nanoparticles | [31] |
| Influenza | Nucleoprotein, acidic polymerase | Intranasal administration | Oligofectamine | [32] |
| H1N1 | Nucleoprotein, acidic polymerase | Inhalation | PH-responsive peptides | [33] |
| RSV | N-protein | Intranasal administration | Naked siRNA | [34] |
| RSV | RSV-protein | Intranasal administration | Naked siRNA | [35] |
| RSV | NSP1 | Intranasal administration | Chitosan nanoparticles | [36] |
| Pneumonia | TNF-α | Intratracheal delivery | RC-NCs | [30] |
| Tuberculosis | TGFβ1 | Inhalation | Naked siRNA | [37] |
| Tuberculosis | XCL1 | Oro-tracheal administration | Naked siRNA | [29] |
2.3. Chronic Respiratory Disease
| Disease | Target | Administration | Delivery System | Ref. |
|---|---|---|---|---|
| Asthma | GATA3 | Inhalation | Tf-Mel-PEI | [40] |
| Asthma | Chil3, Chil4 | Intratracheal Administration | HMG-OR | [43] |
| Asthma | VDBP | Intra-tracheal instillation | DEXA-PEI | [44] |
| Asthma | c-Kit | Intranasal administration | Modified siRNA | [45] |
| Asthma | SOCS3 | Intranasal administration | Naked siRNA | [46] |
| Asthma | Syk | Intranasal administration | Naked siRNA | [47] |
| COPD | RIP2 | Intratracheal administration | Naked siRNA | [48] |
| COPD | RPS3 | Intratracheal administration | Naked siRNA | [49] |
| COPD/ILD | Cytokine Signaling | Nasal instillation | CaP-PLGA | [41] |
| COPD | MAP3K19 | Intratracheal administration | Naked siRNA | [50] |
3. Inhaled siRNA Barriers
3.1. Pulmonary Barriers
3.1.1. Airway Defense
3.1.2. Alveolar Defense
3.2. Cell Barriers
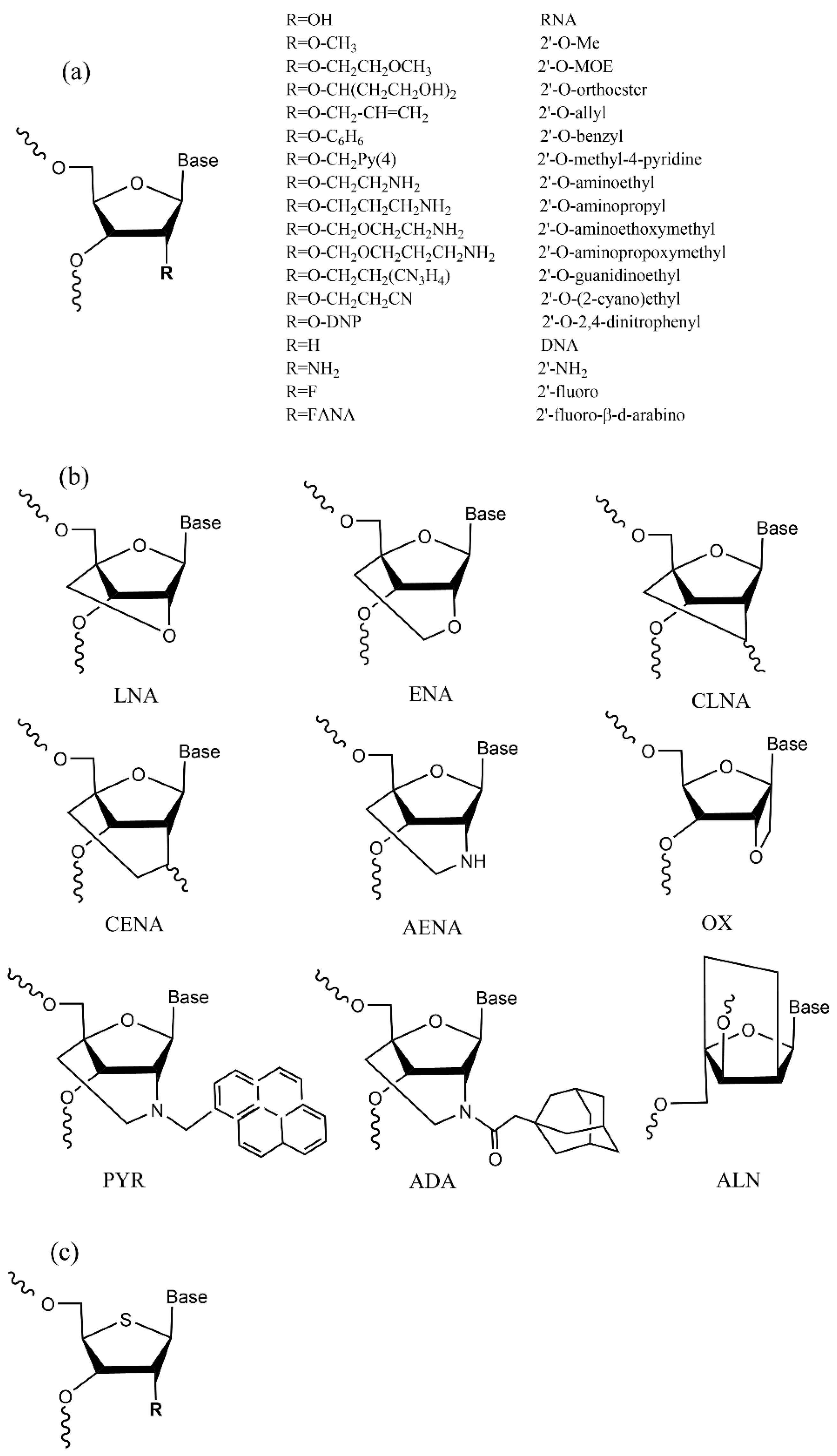
4. siRNA Structural Modification
4.1. Ribose Modification
4.2. Phosphate Backbone Modification
4.3. Base Modification
5. siRNA Inhalation Delivery Systems
5.1. Lipid Nanoparticles
5.1.1. Liposomes
5.1.2. Solid Lipid Nanoparticles
5.1.3. Nanostructured Lipid Carriers
5.1.4. Nanoemulsions
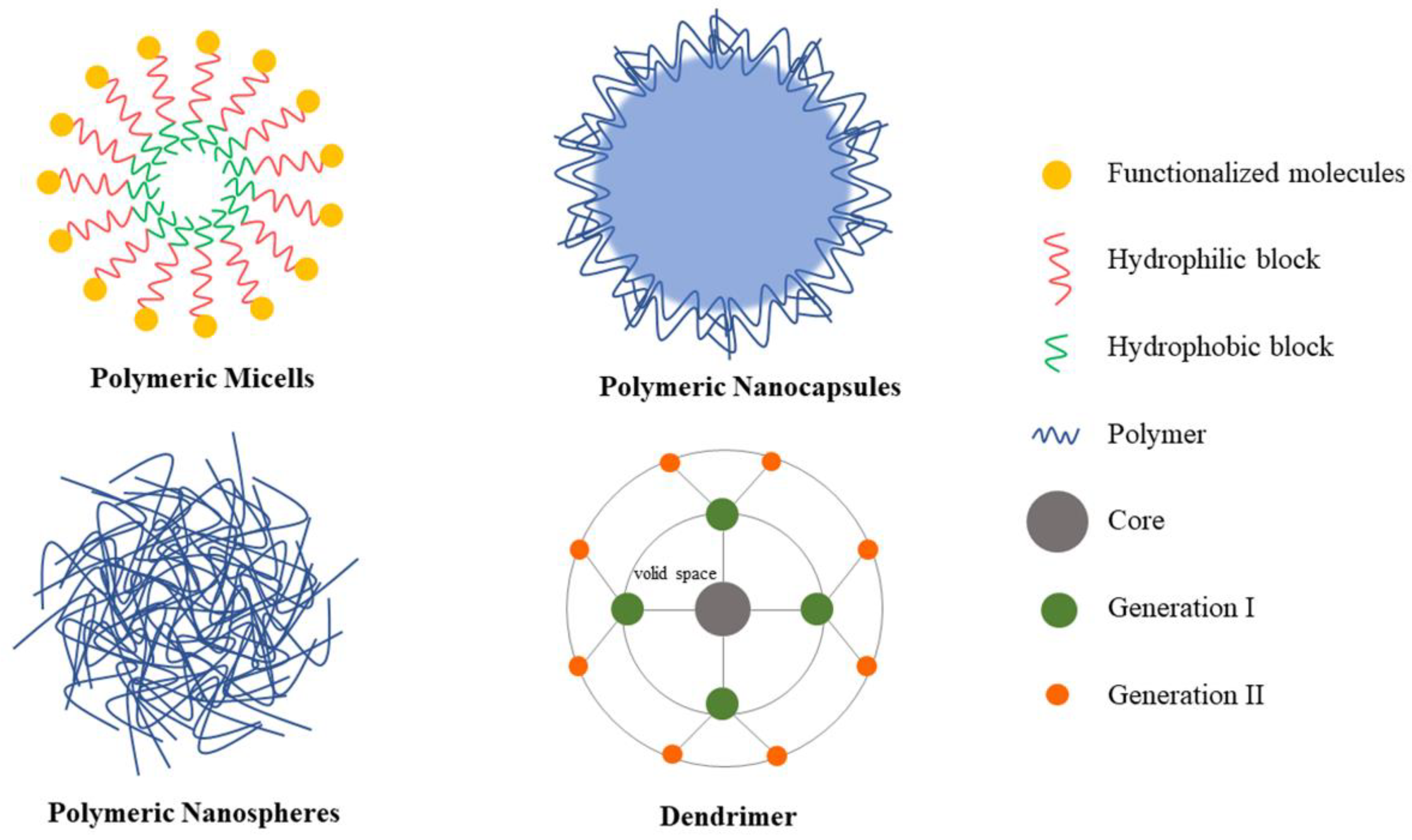
5.2. Polymeric Nanoparticles
5.2.1. Polymeric Micelles
5.2.2. Polymer Nanocapsules
5.2.3. Nanospheres
5.2.4. Dendrimers
5.3. Pulmonary Surfactants
6. Clinical Application
| Year | Therapeutic Name | Disease | Delivery Route | Phase Stage | Target | NCT ID | Ref. |
|---|---|---|---|---|---|---|---|
| 2020 | ALN-HSD | NASH | Subcutaneous | I | HSD17B13 | NCT04565717 | [153] |
| 2020 | DCR-PHXC | PH3 | Subcutaneous | I | LDHA | NCT04555486 | [154] |
| 2019 | Vutrisiran | ATTR With Cardiomyopathy | Subcutaneous | III | TTR | NCT04153149 | [155] |
| 2019 | DCR-PHXC | PH1, PH2, Kidney Diseases | Subcutaneous | II | LDHA | NCT03847909 | [154] |
| 2018 | Lumasiran | PH1 | Subcutaneous | II | Glycolate oxidase | NCT03350451 | [156] |
| 2018 | DCR-HBVS | Chronic Hepatitis B | Subcutaneous | I | HBV transcripts | NCT03772249 | [157] |
| 2018 | siG12D-LODER | Pancreatic Cancer | Intravenous | II | KRAS G12D, all additional G12X mutations | NCT01676259 | [6] |
| 2018 | Inclisiran | ACD | Intravenous | III | PCSK9 | NCT03705234 | [158] |
| 2016 | ARC-AAT | AATD | Intravenous | II | Z-AAT | NCT02900183 | [159] |
| 2016 | ALN-HBV | HBV | Subcutaneous | I | HBV RNA | NCT02826018 | [160] |
| 2016 | DCR-PH1 | PH1 | Intravenous | I | GO | NCT02795325 | [161] |
| 2016 | ALN-TTRSC02 | ATTR Amyloidosis | Subcutaneous | I | TTR | NCT02797847 | [162] |
| 2015 | ARC-520 | Chronic Hepatitis B | Intravenous | II | HBV DNA | NCT02349126 | [163] |
| 2015 | ALN-CC5 | PNH | Subcutaneous | I, II | C5 | NCT02352493 | [164] |
| 2015 | Fitusiran | Hemophilia A, B | Subcutaneous | I, II | AT | NCT02554773 | [165] |
| 2015 | ALN-AS1 | AIP | Subcutaneous | I | ALAS1 | NCT02452372 | [166] |
| 2014 | SYL040012 | Open Angle Glaucoma | Ocular topical | II | ADRβ2 | NCT02250612 | [68] |
| 2014 | ALN-TTR02 | TTR-Mediated Amyloidosis | Intravenous | I | TTR | NCT02053454 | [167] |
| 2014 | ALN-PCSSC | Hypercholesterolemia | Subcutaneous | I | PCSK9 | NCT02314442 | [168] |
| 2013 | ALN-TTRSC | TTR-Mediated Amyloidosis | Subcutaneous | II | TTR | NCT01981837 | [169] |
| 2012 | SYL1001 | Ocular Pain | Ocular topical | I, II | TRPV1 | NCT01776658 | [170] |
| 2012 | AVI-7100 | Influenza | Intravenous | I | Influenza A M1/M2 | NCT01747148 | [171] |
| Approval Date | Company | Therapeutic Name | Disease | Target | Delivery Route | Chemical Modification | Delivery System |
|---|---|---|---|---|---|---|---|
| August 2018 | Alnylam | Patisiran | hATTR | TTR | Intravenous | 2′-OMe modification | Second-generation LNPs |
| November 2019 | Alnylam | Givosiran | AHP | ALAS1 | Subcutaneous | PS linkages, 2′-OMe, 2′-F modification | GalNAc ligand conjugate |
| November 2020 | Alnylam | Lumasiran | PH1 | HAO1 | Subcutaneous | PS linkages, 2′-OMe, 2′-F modification | GalNAc ligand conjugate |
| December 2020 | Alnylam Novartis | Inclisiran | ACD | PCSK9 | Subcutaneous | PS linkages, 2′-OMe, 2′-OMOE, 2′-F modification | GalNAc ligand conjugate |
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ford, L.P.; Toloue, M.M. Delivery of RNAi mediators. Wiley Interdiscip. Rev. RNA 2010, 1, 341–350. [Google Scholar] [CrossRef]
- Castanotto, D.; Rossi, J.J. The promises and pitfalls of RNA-interference-based therapeutics. Nature 2009, 457, 426–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Zhong, L.; Weng, Y.; Peng, L.; Huang, Y.; Zhao, Y.; Liang, X. Therapeutic siRNA: State of the art. Signal Transduct. Target. Ther. 2020, 5, 101. [Google Scholar] [CrossRef]
- McAnuff, M.A.; Rettig, G.R.; Rice, K.G. Potency of siRNA versus shRNA mediated knockdown in vivo. J. Pharm. Sci. 2007, 96, 2922–2930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.M.; Bahal, R.; Rasmussen, T.P.; Manautou, J.E.; Zhong, X.-B. The growth of siRNA-based therapeutics: Updated clinical studies. Biochem. Pharmacol. 2021, 189, 114432. [Google Scholar] [CrossRef] [PubMed]
- Varghese, A.M.; Ang, C.; Dimaio, C.J.; Javle, M.M.; Gutierrez, M.; Yarom, N.; Stemmer, S.M.; Golan, T.; Geva, R.; Semenisty, V.; et al. A phase II study of siG12D-LODER in combination with chemotherapy in patients with locally advanced pancreatic cancer (PROTACT). J. Clin. Oncol. 2020, 38, TPS4672. [Google Scholar] [CrossRef]
- Burnett, J.C.; Rossi, J.J.; Tiemann, K. Current progress of siRNA/shRNA therapeutics in clinical trials. Biotechnol. J. 2011, 6, 1130–1146. [Google Scholar] [CrossRef] [Green Version]
- Cryan, S.A.; Sivadas, N.; Contreras, L.G. In vivo animal models for drug delivery across the lung mucosal barrier. Adv. Drug Deliv. Rev. 2007, 59, 1133–1151. [Google Scholar] [CrossRef]
- de Fougerolles, A.; Novobrantseva, T. siRNA and the lung: Research tool or therapeutic drug? Curr. Opin. Pharmacol. 2008, 8, 280–285. [Google Scholar] [CrossRef]
- Merkel, O.M.; Rubinstein, I.; Kissel, T. siRNA delivery to the lung: What’s new? Adv. Drug. Deliv Rev 2014, 75, 112–128. [Google Scholar] [CrossRef] [Green Version]
- Lam, J.K.-W.; Liang, W.; Chan, H.-K. Pulmonary delivery of therapeutic siRNA. Adv. Drug Deliv. Rev. 2012, 64, 1–15. [Google Scholar] [CrossRef]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA A Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Travis, W.D.; Brambilla, E.; Burke, A.P.; Marx, A.; Nicholson, A.G. Introduction to the 2015 World Health Organization classification of tumors of the lung, pleura, thymus, and heart. J. Thorac. Oncol. 2015, 10, 1240–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, W.A.; Lam, D.C.L.; O’Toole, S.A.; Minna, J.D. Molecular biology of lung cancer. J. Thorac. Dis. 2013, 5 (Suppl. S5), S479–S490. [Google Scholar] [PubMed]
- Garbuzenko, O.B.; Kuzmov, A.; Taratula, O.; Pine, S.R.; Minko, T. Strategy to enhance lung cancer treatment by five essential elements: Inhalation delivery, nanotechnology, tumor-receptor targeting, chemo- and gene therapy. Theranostics 2019, 9, 8362–8376. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.; Lalani, R.; Vhora, I.; Bardoliwala, D.; Patel, A.; Ghosh, S.; Misra, A. Co-delivery of cisplatin and siRNA through hybrid nanocarrier platform for masking resistance to chemotherapy in lung cancer. Drug Deliv. Transl. Res. 2021, 11, 2052–2071. [Google Scholar] [CrossRef]
- Patel, V.; Bardoliwala, D.; Lalani, R.; Patil, S.; Ghosh, S.; Javia, A.; Misra, A. Development of a dry powder for inhalation of nanoparticles codelivering cisplatin and ABCC3 siRNA in lung cancer. Ther. Deliv. 2021, 12, 651–670. [Google Scholar] [CrossRef]
- Han, Y.; Yang, Y.; Sun, Q.; Li, B.; Yue, C.; Liu, Y.; de la Fuente, J.M.; Cui, D. Dual-targeted lung cancer therapy via inhalation delivery of UCNP-siRNA-AS1411 nanocages. Cancer Biol. Med. 2021, 18, 1–5. [Google Scholar] [CrossRef]
- Conte, C.; Monteiro, P.F.; Gurnani, P.; Stolnik, S.; Ungaro, F.; Quaglia, F.; Clarke, P.; Grabowska, A.; Kavallaris, M.; Alexander, C. Multi-component bioresponsive nanoparticles for synchronous delivery of docetaxel and TUBB3 siRNA to lung cancer cells. Nanoscale 2021, 13, 11414–11426. [Google Scholar] [CrossRef]
- Taratula, O.; Kuzmov, A.; Shah, M.; Garbuzenko, O.B.; Minko, T. Nanostructured lipid carriers as multifunctional nanomedicine platform for pulmonary co-delivery of anticancer drugs and siRNA. J. Control. Release 2013, 171, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.-X.; Jere, D.; Jin, H.; Chang, S.-H.; Chung, Y.-S.; Shin, J.-Y.; Kim, J.-E.; Park, S.-J.; Lee, Y.-H.; Chae, C.-H.; et al. Poly(ester amine)-mediated, aerosol-delivered Akt1 small interfering RNA suppresses lung tumorigenesis. Am. J. Respir. Crit. Care Med. 2008, 178, 60–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, G.; Choi, H.-w.; Lee, S.; Choi, J.; Yu, Y.H.; Park, D.-E.; Choi, Y.; Kim, C.-W.; Oh, Y.-K. Enhanced intrapulmonary delivery of anticancer siRNA for lung cancer therapy using cationic ethylphosphocholine-based nanolipoplexes. Mol. Ther. 2013, 21, 816–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Chen, G.; Ding, L.; Wang, Y.; Zhu, C.; Wang, K.; Li, J.; Sun, M.; Oupicky, D. Increased survival by pulmonary treatment of established lung metastases with dual STAT3/CXCR4 inhibition by siRNA nanoemulsions. Mol. Ther. 2019, 27, 2100–2110. [Google Scholar] [CrossRef] [PubMed]
- Taratula, O.; Garbuzenko, O.B.; Chen, A.M.; Minko, T. Innovative strategy for treatment of lung cancer: Targeted nanotechnology-based inhalation co-delivery of anticancer drugs and siRNA. J. Drug Target. 2011, 19, 900–914. [Google Scholar] [CrossRef]
- Templeton, K.E. Why diagnose respiratory viral infection? J. Clin. Virol. 2007, 40, S2–S4. [Google Scholar] [CrossRef]
- Hanada, S.; Pirzadeh, M.; Carver, K.Y.; Deng, J.C. Respiratory viral infection-induced microbiome alterations and secondary bacterial pneumonia. Front. Immunol. 2018, 9, 2640–2655. [Google Scholar] [CrossRef] [Green Version]
- Bitko, V.; Musiyenko, A.; Shulyayeva, O.; Barik, S. Inhibition of respiratory viruses by nasally administered siRNA. Nat. Med. 2005, 11, 50–55. [Google Scholar] [CrossRef]
- Man, D.K.; Chow, M.Y.; Casettari, L.; Gonzalez-Juarrero, M.; Lam, J.K. Potential and development of inhaled RNAi therapeutics for the treatment of pulmonary tuberculosis. Adv. Drug Deliv. Rev. 2016, 102, 21–32. [Google Scholar] [CrossRef]
- Rosas-Taraco, A.G.; Higgins, D.M.; Sanchez-Campillo, J.; Lee, E.J.; Orme, I.M.; Gonzalez-Juarrero, M. Intrapulmonary delivery of XCL1-targeting small interfering RNA in mice chronically infected with Mycobacterium tuberculosis. Am. J. Respir. Cell Mol. Biol. 2009, 41, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Duan, S.; Ye, H.; Ge, C.; Piao, C.; Chen, Y.; Lee, M.; Yin, L. Pro-peptide-reinforced, mucus-penetrating pulmonary siRNA delivery mitigates cytokine storm in pneumonia. Adv. Funct. Mater. 2021, 31, 2008960:1–2008960:13. [Google Scholar] [CrossRef]
- Jamali, A.; Mottaghitalab, F.; Abdoli, A.; Dinarvand, M.; Esmailie, A.; Kheiri, M.T.; Atyabi, F. Inhibiting influenza virus replication and inducing protection against lethal influenza virus challenge through chitosan nanoparticles loaded by siRNA. Drug Deliv. Transl. Res. 2018, 8, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, S.M.; Lo, C.-Y.; Tumpey, T.M.; Epstein, S.L. Protection against lethal influenza virus challenge by RNA interference in vivo. Proc. Natl. Acad. Sci. USA 2004, 101, 8682–8686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, W.; Chow, M.Y.T.; Lau, P.N.; Zhou, Q.T.; Kwok, P.C.L.; Leung, G.P.H.; Mason, A.J.; Chan, H.-K.; Poon, L.L.M.; Lam, J.K.W. Inhalable dry powder formulations of siRNA and pH-responsive peptides with antiviral activity against H1N1 influenza virus. Mol. Pharm. 2015, 12, 910–921. [Google Scholar] [CrossRef] [PubMed]
- Zamora, M.R.; Budev, M.; Rolfe, M.; Gottlieb, J.; Humar, A.; Devincenzo, J.; Vaishnaw, A.; Cehelsky, J.; Albert, G.; Nochur, S.; et al. RNA interference therapy in lung transplant patients infected with respiratory syncytial virus. Am. J. Respir. Crit. Care Med. 2011, 183, 531–538. [Google Scholar] [CrossRef]
- Zhang, W.; Tripp, R.A. RNA interference inhibits respiratory syncytial virus replication and disease pathogenesis without inhibiting priming of the memory immune response. J. Virol. 2008, 82, 12221–12231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Yang, H.; Kong, X.; Mohapatra, S.; Juan-Vergara, H.S.; Hellermann, G.; Behera, S.; Singam, R.; Lockey, R.F.; Mohapatra, S.S. Inhibition of respiratory syncytial virus infection with intranasal siRNA nanoparticles targeting the viral NS1 gene. Nat. Med. 2005, 11, 56–62. [Google Scholar] [CrossRef]
- Rosas-Taraco, A.G.; Higgins, D.M.; Sánchez-Campillo, J.; Lee, E.J.; Orme, I.M.; González-Juarrero, M. Local pulmonary immunotherapy with siRNA targeting TGFβ1 enhances antimicrobial capacity in Mycobacterium tuberculosis infected mice. Tuberculosis 2011, 91, 98–106. [Google Scholar] [CrossRef]
- Gayle, A.V.; Axson, E.L.; Bloom, C.I.; Navaratnam, V.; Quint, J.K. Changing causes of death for patients with chronic respiratory disease in England, 2005–2015. Thorax 2019, 74, 483–491. [Google Scholar] [CrossRef]
- Dua, K.; Wadhwa, R.; Singhvi, G.; Rapalli, V.; Shukla, S.D.; Shastri, M.D.; Gupta, G.; Satija, S.; Mehta, M.; Khurana, N.; et al. The potential of siRNA based drug delivery in respiratory disorders: Recent advances and progress. Drug Dev. Res. 2019, 80, 714–730. [Google Scholar] [CrossRef]
- Keil, T.W.M.; Baldassi, D.; Merkel, O.M. T-cell targeted pulmonary siRNA delivery for the treatment of asthma. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2020, 12, e1634:1–e1634:13. [Google Scholar] [CrossRef] [Green Version]
- Frede, A.; Neuhaus, B.; Knuschke, T.; Wadwa, M.; Kollenda, S.; Klopfleisch, R.; Hansen, W.; Buer, J.; Bruder, D.; Epple, M.; et al. Local delivery of siRNA-loaded calcium phosphate nanoparticles abates pulmonary inflammation. Nanomedicine 2017, 13, 2395–2403. [Google Scholar] [CrossRef] [PubMed]
- Agustí, A.; Edwards, L.D.; Rennard, S.I.; MacNee, W.; Tal-Singer, R.; Miller, B.E.; Vestbo, J.; Lomas, D.A.; Calverley, P.M.A.; Wouters, E.; et al. Persistent systemic inflammation is associated with poor clinical outcomes in COPD: A novel phenotype. PLoS ONE 2012, 7, e37483. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.; Jeong, H.; Kim, S.; Kim, M.; Lee, M.; Rhim, T. Targeted delivery of Chil3/Chil4 siRNA to alveolar macrophages using ternary complexes composed of HMG and oligoarginine micelles. Nanoscale 2020, 12, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.; Gu, J.; Lee, M.; Rhim, T. A new combination therapy for asthma using dual-function dexamethasone-conjugated polyethylenimine and vitamin D binding protein siRNA. Gene Ther. 2017, 24, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Chen, H.; Li, Y.-M.; Wang, S.-Y.; Diao, X.; Liu, K.-G. Intranasal siRNA targeting c-Kit reduces airway inflammation in experimental allergic asthma. Int. J. Clin. Exp. Pathol. 2014, 7, 5505–5514. [Google Scholar]
- Zafra, M.P.; Mazzeo, C.; Gámez, C.; Marco, A.R.; de Zulueta, A.; Sanz, V.; Bilbao, I.; Ruiz-Cabello, J.; Zubeldia, J.M.; del Pozo, V. Gene silencing of SOCS3 by siRNA intranasal delivery inhibits asthma phenotype in mice. PLoS ONE 2014, 9, e91996. [Google Scholar]
- Huang, Z.Y.; Kim, M.K.; Han, T.H.K.; Indik, Z.K.; Schreiber, A.D. Effect of locally administered Syk siRNA on allergen-induced arthritis and asthma. Mol. Immunol. 2013, 53, 52–59. [Google Scholar] [CrossRef]
- Dong, J.; Liao, W.; Tan, L.H.; Yong, A.; Peh, W.Y.; Wong, W.S.F. Gene silencing of receptor-interacting protein 2 protects against cigarette smoke-induced acute lung injury. Pharmacol. Res. 2019, 139, 560–568. [Google Scholar] [CrossRef]
- Dong, J.; Liao, W.; Peh, H.Y.; Tan, W.S.D.; Zhou, S.; Wong, W.S.F. Ribosomal protein S3 gene silencing protects against cigarette smoke-induced acute lung injury. Mol. Ther.-Nucleic Acids 2018, 12, 370–380. [Google Scholar] [CrossRef] [Green Version]
- Boehme, S.A.; Franz-Bacon, K.; Ludka, J.; DiTirro, D.N.; Ly, T.W.; Bacon, K.B. MAP3K19 is overexpressed in COPD and is a central mediator of cigarette smoke-induced pulmonary inflammation and lower airway destruction. PLoS ONE 2016, 11, e0167169:1–e0167169:26. [Google Scholar] [CrossRef]
- Wang, J.; Lu, Z.; Wientjes, M.G.; Au, J.L.-S. Delivery of siRNA therapeutics: Barriers and carriers. AAPS J. 2010, 12, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Sou, T.; Bergström, C.A.S. Contemporary formulation development for inhaled pharmaceuticals. J. Pharm. Sci. 2021, 110, 66–86. [Google Scholar] [CrossRef] [PubMed]
- Youngren-Ortiz, S.R.; Gandhi, N.S.; Espana-Serrano, L.; Chougule, M.B. Aerosol delivery of siRNA to the lungs. Part 1: Rationale for gene delivery systems. KONA Powder Part. J. 2016, 33, 63–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munkholm, M.; Mortensen, J. Mucociliary clearance: Pathophysiological aspects. Clin. Physiol. Funct. Imaging 2014, 34, 171–177. [Google Scholar] [CrossRef]
- Chen, D.; Liu, J.; Wu, J.; Suk, J.S. Enhancing nanoparticle penetration through airway mucus to improve drug delivery efficacy in the lung. Expert Opin. Drug Deliv. 2021, 18, 595–606. [Google Scholar] [CrossRef]
- Olsson, B.; Bondesson, E.; Borgström, L.; Edsbäcker, S.; Eirefelt, S.; Ekelund, K.; Gustavsson, L.; Hegelund-Myrbäck, T. Pulmonary Drug Metabolism, Clearance, and Absorption. In Controlled Pulmonary Drug Delivery. Advances in Delivery Science and Technology; Springer: New York, NY, USA, 2011; pp. 21–50. [Google Scholar]
- Chang, R.Y.K.; Kwok, P.C.L.; Ghassabian, S.; Brannan, J.D.; Koskela, H.O.; Chan, H.-K. Cough as an adverse effect on inhalation pharmaceutical products. Br. J. Pharmacol. 2020, 177, 4096–4112. [Google Scholar] [CrossRef]
- Edsbäcker, S.; Wollmer, P.; Selroos, O.; Borgström, L.; Olsson, B.; Ingelf, J. Do airway clearance mechanisms influence the local and systemic effects of inhaled corticosteroids? Pulm. Pharmacol. Ther. 2008, 21, 247–258. [Google Scholar] [CrossRef]
- Zuo, Y.Y.; Veldhuizen, R.A.W.; Neumann, A.W.; Petersen, N.O.; Possmayer, F. Current perspectives in pulmonary surfactant—Inhibition, enhancement and evaluation. Biochim. Biophys. Acta Biomembr. 2008, 1778, 1947–1977. [Google Scholar] [CrossRef] [Green Version]
- Backer, L.D.; Cerrada, A.; Pérez-Gil, J.; Smedt, S.C.D.; Raemdonck, K. Bio-inspired materials in drug delivery: Exploring the role of pulmonary surfactant in siRNA inhalation therapy. J. Control. Release 2015, 220, 642–650. [Google Scholar] [CrossRef] [Green Version]
- Joshi, N.; Walter, J.M.; Misharin, A.V. Alveolar macrophages. Cell. Immunol. 2018, 330, 86–90. [Google Scholar] [CrossRef]
- Rubins, J.B. Alveolar macrophages: Wielding the double-edged sword of inflammation. Am. J. Respir. Crit. Care Med. 2003, 167, 103–104. [Google Scholar] [CrossRef] [PubMed]
- Juliano, R.L.; Carver, K.; Cao, C.; Ming, X. Receptors, endocytosis, and trafficking: The biological basis of targeted delivery of antisense and siRNA oligonucleotides. J. Drug Target. 2013, 21, 27–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominska, M.; Dykxhoorn, D.M. Breaking down the barriers: siRNA delivery and endosome escape. J. Cell Sci. 2010, 123, 1183–1189. [Google Scholar] [CrossRef] [Green Version]
- Shim, M.S.; Kwon, Y.J. Efficient and targeted delivery of siRNA in vivo. FEBS J. 2010, 277, 4814–4827. [Google Scholar] [CrossRef]
- Deleavey, G.F.; Watts, J.K.; Damha, M.J. Chemical modification of siRNA. Curr. Protoc. Nucleic Acid Chem. 2009, 39, 16.3.1–16.3.22. [Google Scholar] [CrossRef] [PubMed]
- Nawrot, B.; Sipa, K. Chemical and structural diversity of siRNA molecules. Curr. Top. Med. Chem. 2006, 6, 913–925. [Google Scholar] [CrossRef]
- Chen, C.; Yang, Z.; Tang, X. Chemical modifications of nucleic acid drugs and their delivery systems for gene-based therapy. Med. Res. Rev. 2018, 38, 829–869. [Google Scholar] [CrossRef]
- Prakash, T.P.; Allerson, C.R.; Dande, P.; Vickers, T.A.; Sioufi, N.; Jarres, R.; Baker, B.F.; Swayze, E.E.; Griffey, R.H.; Bhat, B. Positional effect of chemical modifications on short interference RNA activity in mammalian cells. J. Med. Chem. 2005, 48, 4247–4553. [Google Scholar] [CrossRef]
- Bramsen, J.B.; Grünweller, A.; Hartmann, R.K.; Kjems, J. Using Chemical Modification to Enhance siRNA Performance. In Handbook of RNA Biochemistry: Second, Completely Revised and Enlarged Edition; Hartmann, R.K., Bindereif, A., Schön, A., Westhof, E., Eds.; Wiley: Hoboken, NJ, USA, 2014; pp. 1243–1278. ISBN 9783527327645. [Google Scholar]
- Kenski, D.M.; Butora, G.; Willingham, A.T.; Cooper, A.J.; Fu, W.; Qi, N.; Soriano, F.; Davies, I.W.; Flanagan, W.M. siRNA-optimized modifications for enhanced in vivo activity. Mol. Ther. Nucleic Acids 2012, 1, e5:1–e5:8. [Google Scholar] [CrossRef]
- Odadzic, D.; Bramsen, J.B.; Smicius, R.; Bus, C.; Kjems, J.; Engels, J.W. Synthesis of 2’-O-modified adenosine building blocks and application for RNA interference. Bioorg. Med. Chem. 2008, 16, 518–529. [Google Scholar] [CrossRef]
- Layzer, J.M.; McCaffrey, A.P.; Tanner, A.K.; Huang, Z.; Kay, M.A.; Sullenger, B.A. In vivo activity of nuclease-resistant siRNAs. RNA 2004, 10, 766–771. [Google Scholar] [CrossRef] [Green Version]
- Watts, J.K.; Deleavey, G.F.; Damha, M.J. Chemically modified siRNA: Tools and applications. Drug Discov. Today 2008, 13, 842–855. [Google Scholar] [CrossRef] [PubMed]
- Bramsen, J.B.; Laursen, M.B.; Nielsen, A.F.; Hansen, T.B.; Bus, C.; Langkjaer, N.; Babu, B.R.; Hojland, T.; Abramov, M.; Van Aerschot, A.; et al. A large-scale chemical modification screen identifies design rules to generate siRNAs with high activity, high stability and low toxicity. Nucleic Acids Res. 2009, 37, 2867–2881. [Google Scholar] [CrossRef]
- Zhou, C.; Chattopadhyaya, J. Challenges in the Chemistry of Small Interfering RNA as Potential Therapeutics to Inhibit Cellular mRNA Expression. In Reference Module in Chemistry, Molecular Sciences and Chemical Engineering; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Fluiter, K.; Mook, O.R.F.; Baas, F. The Therapeutic Potential of LNA-Modified siRNAs: Reduction of Off-Target Effects by Chemical Modification of the siRNA Sequence; Humana Press: Totowa, NJ, USA, 2009; pp. 1–15. ISBN 978-1-60327-546-0. [Google Scholar]
- Elmén, J.; Thonberg, H.; Ljungberg, K.; Frieden, M.; Westergaard, M.; Xu, Y.; Wahren, B.; Liang, Z.; Ørum, H.; Koch, T.; et al. Locked nucleic acid (LNA) mediated improvements in siRNA stability and functionality. Nucleic Acids Res. 2005, 33, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Sumaria, C.S.; Pradeepkumar, P.I. Exploring chemical modifications for siRNA therapeutics: A structural and functional outlook. Chemmedchem 2010, 5, 328–349. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, M.; Takagi-Sato, M.; Okuyama, R.; Araki, K.; Sun, W.; Nakai, D.; Tsutsumi, S.; Kawai, k. Direct comparison of in vivo antisense activity of ENA oligonucleotides targeting PTP1B mRNA with that of 2′-O-(2-methoxy) ethyl-modified oligonucleotides. Oligonucleotides 2006, 16, 253–262. [Google Scholar] [CrossRef]
- Wan, W.B.; Seth, P.P. The medicinal chemistry of therapeutic oligonucleotides. J. Med. Chem. 2016, 59, 9645–9667. [Google Scholar] [CrossRef]
- Dande, P.; Prakash, T.P.; Sioufi, N.; Gaus, H.; Jarres, R.; Berdeja, A.; Swayze, E.E.; Griffey, R.H.; Bhat, B. Improving RNA interference in mammalian cells by 4‘-thio-modified small interfering RNA (siRNA) effect on siRNA activity and nuclease stability when used in combination with 2-O-alkyl modifications. J. Med. Chem. 2006, 49, 1624–1634. [Google Scholar] [CrossRef]
- Hoshika, S.; Minakawa, N.; Kamiya, H.; Harashima, H.; Matsuda, A. RNA interference induced by siRNAs modified with 4’-thioribonucleosides in cultured mammalian cells. FEBS Lett. 2005, 579, 3115–3118. [Google Scholar] [CrossRef] [Green Version]
- Hoshika, S.; Minakawa, N.; Shionoya, A.; Imada, K.; Ogawa, N.; Matsuda, A. Study of modification pattern-RNAi activity relationships by using siRNAs modified with 4’-thioribonucleosides. Chembiochem 2007, 8, 2133–2138. [Google Scholar] [CrossRef]
- Prakash, T.P.; Kinberger, G.A.; Murray, H.M.; Chappell, A.; Riney, S.; Graham, M.J.; Lima, W.F.; Swayze, E.E.; Seth, P.P. Synergistic effect of phosphorothioate, 5′-vinylphosphonate and GalNAc modifications for enhancing activity of synthetic siRNA. Bioorg. Med. Chem. Lett. 2016, 26, 2817–2820. [Google Scholar] [CrossRef] [PubMed]
- Jahns, H.; Roos, M.; Imig, J.; Baumann, F.; Wang, Y.; Gilmour, R.; Hall, J. Stereochemical bias introduced during RNA synthesis modulates the activity of phosphorothioate siRNAs. Nat. Commun. 2015, 6, 6317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Sierant, M.; Janicka, M.; Peczek, L.; Martinez, C.; Hassell, T.; Li, N.; Li, X.; Wang, T.; Nawrot, B. Gene silencing activity of siRNA molecules containing phosphorodithioate substitutions. ACS Chem. Biol. 2012, 7, 1214–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meade, B.R.; Gogoi, K.; Hamil, A.S.; Palm-Apergi, C.; Berg, A.v.d.; Hagopian, J.C.; Springer, A.D.; Eguchi, A.; Kacsinta, A.D.; Dowdy, C.F.; et al. Efficient delivery of RNAi prodrugs containing reversible charge-neutralizing phosphotriester backbone modifications. Nat. Biotechnol. 2014, 32, 1256–1261. [Google Scholar] [CrossRef] [Green Version]
- Tsubaki, K.; Hammill, M.L.; Varley, A.J.; Kitamura, M.; Okauchi, T.; Desaulniers, J.-P. Synthesis and evaluation of neutral phosphate triester backbone-modified siRNAs. ACS Med. Chem. Lett. 2020, 11, 1457–1462. [Google Scholar] [CrossRef]
- Threlfall, R.N.; Torres, A.G.; Krivenko, A.; Gait, M.J.; Caruthers, M.H. Synthesis and biological activity of phosphonoacetate-and thiophosphonoacetate-modified 2′-O-methyl oligoribonucleotides. Org. Biomol. Chem. 2012, 10, 746–754. [Google Scholar] [CrossRef]
- Hall, A.H.S.; Wan, J.; Shaughnessy, E.E.; Shaw, B.R.; Alexander, K.A. RNA interference using boranophosphate siRNAs: Structure-activity relationships. Nucleic Acids Res. 2004, 32, 5991–6000. [Google Scholar] [CrossRef] [Green Version]
- Prakash, T.P.; Kraynack, B.; Baker, B.F.; Swayze, E.E.; Bhat, B. RNA interference by 2′, 5′-linked nucleic acid duplexes in mammalian cells. Bioorg. Med. Chem. Lett. 2006, 16, 3238–3240. [Google Scholar] [CrossRef]
- Habibian, M.; Harikrishna, S.; Fakhoury, J.; Barton, M.; Ageely, E.A.; Cencic, R.; Fakih, H.H.; Katolik, A.; Takahashi, M.; Rossi, J.; et al. Effect of 2’-5’/3’-5’ phosphodiester linkage heterogeneity on RNA interference. Nucleic Acids Res. 2020, 48, 4643–4657. [Google Scholar] [CrossRef]
- Peacock, H.; Kannan, A.; Beal, P.A.; Burrows, C.J. Chemical modification of siRNA bases to probe and enhance RNA interference. J. Org. Chem. 2011, 76, 7295–7300. [Google Scholar] [CrossRef] [Green Version]
- Hudson, G.A.; Bloomingdale, R.J.; Znosko, B.M. Thermodynamic contribution and nearest-neighbor parameters of pseudouridine-adenosine base pairs in oligoribonucleotides. RNA 2013, 19, 1474–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sipa, K.; Sochacka, E.; Kazmierczak-Baranska, J.; Maszewska, M.; Janicka, M.; Nowak, G.; Nawrot, B. Effect of base modifications on structure, thermodynamic stability, and gene silencing activity of short interfering RNA. RNA 2007, 13, 1301–1316. [Google Scholar] [CrossRef] [Green Version]
- Imaeda, A.; Tomoike, F.; Hayakawa, M.; Nakamoto, K.; Kimura, Y.; Abe, N.; Abe, H. N(6)-methyl adenosine in siRNA evades immune response without reducing RNAi activity. Nucleosides Nucleotides Nucleic Acids 2019, 38, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.-L.; Rana, T.M. siRNA function in RNAi: A chemical modification analysis. RNA 2003, 9, 1034–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somoza, A.; Silverman, A.P.; Milleri, R.M.; Chelliserrykattil, J.; Kool, E.T. Steric effects in RNA interference: Probing the influence of nucleobase size and shape. Chem.—A Eur. J. 2008, 14, 7978–7987. [Google Scholar] [CrossRef]
- Zoulikha, M.; Xiao, Q.; Boafo, G.F.; Sallam, M.A.; Chen, Z.; He, W. Pulmonary delivery of siRNA against acute lung injury/acute respiratory distress syndrome. Acta Pharm. Sin. B 2021, 12, 600–620. [Google Scholar] [CrossRef]
- Ewert, K.K.; Zidovska, A.; Ahmad, A.; Bouxsein, N.F.; Evans, H.M.; McAllister, C.S.; Samuel, C.E.; Safinya, C.R. Cationic liposome—Nucleic acid complexes for gene delivery and silencing: Pathways and mechanisms for plasmid DNA and siRNA. In Nucleic Acid Transfection; Springer: Berlin/Heidelberg, Germany, 2010; pp. 191–226. [Google Scholar]
- Fukushige, K.; Tagami, T.; Ozeki, T. The offset effect of a hyaluronic acid coating to cationic carriers containing siRNA: Alleviated cytotoxicity and retained gene silencing in vitro. J. Drug Deliv. Sci. Technol. 2017, 39, 435–441. [Google Scholar] [CrossRef]
- Fukushige, K.; Tagami, T.; Naito, M.; Goto, E.; Hirai, S.; Hatayama, N.; Yokota, H.; Yasui, T.; Baba, Y.; Ozeki, T. Developing spray-freeze-dried particles containing a hyaluronic acid-coated liposome-protamine-DNA complex for pulmonary inhalation. Int. J. Pharm. 2020, 583, 119338:1–119338:8. [Google Scholar] [CrossRef]
- Cheng, X.; Lee, R.J. The role of helper lipids in lipid nanoparticles (LNPs) designed for oligonucleotide delivery. Adv. Drug Deliv. Rev. 2016, 99, 129–137. [Google Scholar] [CrossRef]
- Kulkarni, J.A.; Darjuan, M.M.; Mercer, J.E.; Chen, S.; van der Meel, R.; Thewalt, J.L.; Tam, Y.Y.C.; Cullis, P.R. On the formation and morphology of lipid nanoparticles containing Ionizable cationic lipids and siRNA. ACS Nano 2018, 12, 4787–4795. [Google Scholar] [CrossRef] [Green Version]
- Habrant, D.; Peuziat, P.; Colombani, T.; Dallet, L.; Gehin, J.; Goudeau, E.; Evrard, B.; Lambert, O.; Haudebourg, T.; Pitard, B. Design of ionizable lipids to overcome the limiting step of endosomal escape: Application in the intracellular delivery of mRNA, DNA, and siRNA. J. Med. Chem. 2016, 59, 3046–3062. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.Y.; Wong, H.L. Tailoring nanostructured solid-lipid carriers for time-controlled intracellular siRNA kinetics to sustain RNAi-mediated chemosensitization. Biomaterials 2011, 32, 2662–2672. [Google Scholar] [CrossRef] [PubMed]
- Hanafy, M.S.; Hufnagel, S.; Trementozzi, A.N.; Sakran, W.; Stachowiak, J.C.; Koleng, J.J.; Cui, Z. PD-1 siRNA-encapsulated solid lipid nanoparticles downregulate PD-1 expression by macrophages and inhibit tumor growth. AAPS PharmSciTech 2021, 22, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-L.; Hanafy, M.S.; Xu, H.; Leal, J.; Zhai, Y.; Ghosh, D.; Williams, R.O.; Smyth, H.D.; Cui, Z. Aerosolizable siRNA-encapsulated solid lipid nanoparticles prepared by thin-film freeze-drying for potential pulmonary delivery. Int. J. Pharm. 2021, 596, 120215:1–120215:9. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, I.; Yasir, M.; Verma, M.; Singh, A.P. Nanostructured lipid carriers: A groundbreaking approach for transdermal drug delivery. Adv. Pharm. Bull. 2020, 10, 150–165. [Google Scholar] [CrossRef] [PubMed]
- Abdulbaqi, I.M.; Assi, R.A.; Yaghmur, A.; Darwis, Y.; Mohtar, N.; Parumasivam, T.; Saqallah, F.G.; Wahab, H.A. Pulmonary delivery of anticancer drugs via lipid-based nanocarriers for the treatment of lung cancer: An update. Pharmaceuticals 2021, 14, 725. [Google Scholar] [CrossRef]
- Patil, T.S.; Deshpande, A.S.; Deshpande, S.; Shende, P. Targeting pulmonary tuberculosis using nanocarrier-based dry powder inhalation: Current status and futuristic need. J. Drug Target. 2019, 27, 12–27. [Google Scholar] [CrossRef]
- Pindiprolu, S.K.; Kumar, C.S.; Golla, V.S.; Likitha, P.; Chandra, S.; Ramachandra, R.K. Pulmonary delivery of nanostructured lipid carriers for effective repurposing of salinomycin as an antiviral agent. Med. Hypotheses 2020, 143, 109858:1–109858:3. [Google Scholar] [CrossRef]
- Nafee, N.; Makled, S.; Boraie, N. Nanostructured lipid carriers versus solid lipid nanoparticles for the potential treatment of pulmonary hypertension via nebulization. Eur. J. Pharm. Sci. 2018, 125, 151–162. [Google Scholar] [CrossRef]
- Almurshedi, A.S.; Aljunaidel, H.A.; Alquadeib, B.; Aldosari, B.N.; Alfagih, I.M.; Almarshidy, S.S.; Eltahir, E.K.D.; Mohamoud, A.Z. Development of inhalable nanostructured lipid carriers for ciprofloxacin for noncystic fibrosis bronchiectasis treatment. Int. J. Nanomed. 2021, 16, 2405–2417. [Google Scholar] [CrossRef]
- Bhatt, P.; Madhav, S. A detailed review on nanoemulsion drug delivery system. Int. J. Pharm. Sci. Res. 2011, 2, 2482–2489. [Google Scholar]
- Jaiswal, M.; Dudhe, R.; Sharma, P.K. Nanoemulsion: An advanced mode of drug delivery system. 3 Biotech 2015, 5, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Wang, Y.; Wu, P.; Zhou, Y.; Yu, F.; Zhu, C.; Li, Z.; Hang, Y.; Wang, K.; Li, J.; et al. Reversibly stabilized polycation nanoparticles for combination treatment of early- and late-stage metastatic breast cancer. ACS Nano 2018, 12, 6620–6636. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Tang, S.; Tang, W.; Mosley, D.D.; Yu, A.; Sil, D.; Romanova, S.; Bailey, K.L.; Knoell, D.L.; Wyatt, T.A.; et al. Perfluorocarbon nanoemulsions enhance therapeutic siRNA delivery in the treatment of pulmonary fibrosis. Adv. Sci. 2022, 9, 2103676:1–2103676:16. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Bansal, M.; Visht, S.; Sharma, P.; Kulkarni, G. Nanoemulsion: A new concept of delivery system. Chron. Young Sci. 2010, 1, 2–6. [Google Scholar]
- Youngren-Ortiz, S.R.; Gandhi, N.S.; España-Serrano, L.; Chougule, M.B. Aerosol delivery of siRNA to the lungs. Part 2: Nanocarrier-based delivery systems. KONA Powder Part. J. 2017, 34, 44–69. [Google Scholar] [CrossRef] [Green Version]
- Aw, M.S.; Kurian, M.; Losic, D. Recent advances in the design, development, and targeting mechanisms of polymeric micelles for delivery of siRNA in cancer therapy. Prog. Polym. Sci. 2017, 64, 154–181. [Google Scholar]
- Jhaveri, A.M.; Torchilin, V.P. Multifunctional polymeric micelles for delivery of drugs and siRNA. Front. Pharmacol. 2014, 5, 77:1–77:26. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-Q.; Sun, C.-Y.; Yang, X.-Z.; Wang, J. Polymeric-micelle-based nanomedicine for siRNA delivery. Part. Part. Syst. Charact. 2013, 30, 211–228. [Google Scholar] [CrossRef]
- Yoon, P.O.; Park, J.W.; Lee, C.-M.; Kim, S.H.; Kim, H.-N.; Ko, Y.; Bae, S.J.; Yun, S.; Park, J.H.; Kwon, T.; et al. Self-assembled micelle interfering RNA for effective and safe targeting of dysregulated genes in pulmonary fibrosis. J. Biol. Chem. 2016, 291, 6433–6446. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, M. Clinical applications of polymeric micelle carrier systems in chemotherapy and image diagnosis of solid tumors. J. Exp. Clin. Med. 2011, 3, 151–158. [Google Scholar] [CrossRef]
- Deng, S.; Gigliobianco, M.R.; Censi, R.; Martino, P.D. Polymeric nanocapsules as nanotechnological alternative for drug delivery system: Current status, challenges and opportunities. Nanomaterials 2020, 10, 847. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Liang, M.; Wen, J.; Liu, Y.; Lu, Y.; Chen, I.S.Y. Single siRNA nanocapsules for enhanced RNAi delivery. J. Am. Chem. Soc. 2012, 134, 13542–13545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora-Huertas, C.E.; Fessi, H.; Elaissari, A. Polymer-based nanocapsules for drug delivery. Int. J. Pharm. 2010, 385, 113–142. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Choi, S.H.; Kim, S.H.; Park, T.G. Thermally sensitive cationic polymer nanocapsules for specific cytosolic delivery and efficient gene silencing of siRNA: Swelling induced physical disruption of endosome by cold shock. J. Control. Release 2008, 125, 25–32. [Google Scholar] [CrossRef]
- Chen, C.-K.; Law, W.-C.; Aalinkeel, R.; Yu, Y.; Nair, B.; Wu, J.; Mahajan, S.; Reynolds, J.L.; Li, Y.; Lai, C.K.; et al. Biodegradable cationic polymeric nanocapsules for overcoming multidrug resistance and enabling drug-gene co-delivery to cancer cells. Nanoscale 2014, 6, 1567–1572. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wang, M.; Guan, S.; Huang, Z.; Liu, S.; Li, X.; Jiang, X.; Luo, Q.; Xu, J.; Liu, J. Cucurbit[8]uril-based supramolecular polymer nanocapsules as an effective siRNA delivery platform for gene therapy. Polym. Chem. 2019, 10, 5659–5664. [Google Scholar] [CrossRef]
- Guterres, S.S.; Alves, M.P.; Pohlmann, A.R. Polymeric nanoparticles, nanospheres and nanocapsules, for cutaneous applications. Drug Target Insights 2007, 2, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Tahara, K.; Yamamoto, H.; Hirashima, N.; Kawashima, Y. Chitosan-modified poly (D, L-lactide-co-glycolide) nanospheres for improving siRNA delivery and gene-silencing effects. Eur. J. Pharm. Biopharm. 2010, 74, 421–426. [Google Scholar] [CrossRef]
- Jensen, D.M.K.; Cun, D.; Maltesen, M.J.; Frokjaer, S.; Nielsen, H.M.; Foged, C. Spray drying of siRNA-containing PLGA nanoparticles intended for inhalation. J. Control. Release 2010, 142, 138–145. [Google Scholar] [CrossRef]
- d’Angelo, I.; Costabile, G.; Durantie, E.; Brocca, P.; Rondelli, V.; Russo, A.; Russo, G.; Miro, A.; Quaglia, F.; Petri-Fink, A.; et al. Hybrid lipid/polymer nanoparticles for pulmonary delivery of siRNA: Development and fate upon in vitro deposition on the human epithelial airway barrier. J. Aerosol Med. Pulm. Drug Deliv. 2018, 31, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Ni, S.; Liu, Y.; Tang, Y.; Chen, J.; Li, S.; Pu, J.; Han, L. GABAB receptor ligand-directed trimethyl chitosan/tripolyphosphate nanoparticles and their pMDI formulation for survivin siRNA pulmonary delivery. Carbohydr. Polym. 2018, 179, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Kesharwani, P.; Jain, K.; Jain, N.K. Dendrimer as nanocarrier for drug delivery. Prog. Polym. Sci. 2014, 39, 268–307. [Google Scholar] [CrossRef]
- Sherje, A.P.; Jadhav, M.; Dravyakar, B.R.; Kadam, D. Dendrimers: A versatile nanocarrier for drug delivery and targeting. Int. J. Pharm. 2018, 548, 707–720. [Google Scholar] [CrossRef]
- Conti, D.S.; Brewer, D.; Grashik, J.; Avasarala, S.; Da Rocha, S.R.P. Poly(amidoamine) dendrimer nanocarriers and their aerosol formulations for siRNA delivery to the lung epithelium. Mol. Pharm. 2014, 11, 1808–1822. [Google Scholar] [CrossRef] [PubMed]
- Bielski, E.; Zhong, Q.; Mirza, H.; Brown, M.; Molla, A.; Carvajal, T.; Da Rocha, S.R.P. TPP-dendrimer nanocarriers for siRNA delivery to the pulmonary epithelium and their dry powder and metered-dose inhaler formulations. Int. J. Pharm. 2017, 527, 171–183. [Google Scholar] [CrossRef]
- Merckx, P.; Lammens, J.; Nuytten, G.; Bogaert, B.; Guagliardo, R.; Maes, T.; Vervaet, C.; Beer, T.D.; Smedt, S.C.D.; Raemdonck, K. Lyophilization and nebulization of pulmonary surfactant-coated nanogels for siRNA inhalation therapy. Eur. J. Pharm. Biopharm. 2020, 157, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Guagliardo, R.; Herman, L.; Penders, J.; Zamborlin, A.; De Keersmaecker, H.; Van de Vyver, T.; Verstraeten, S.; Merckx, P.; Mingeot-Leclercq, M.-P.; Echaide, M.; et al. Surfactant protein B promotes cytosolic SiRNA delivery by adopting a virus-like mechanism of action. ACS Nano 2021, 15, 8095–8109. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Doss, C.G.P.; Lee, S.-S. Therapeutic miRNA and siRNA: Moving from bench to clinic as next generation medicine. Mol. Ther. Nucleic Acids 2017, 8, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, R.; Elbashir, S.; Borland, T.; Toudjarska, I.; Hadwiger, P.; John, M.; Roehl, I.; Morskaya, S.S.; Martinello, R.; Kahn, J.; et al. RNA interference-mediated silencing of the respiratory syncytial virus nucleocapsid defines a potent antiviral strategy. Antimicrob. Agents Chemother. 2009, 53, 3952–3962. [Google Scholar] [CrossRef] [Green Version]
- Xing, Y.; Proesmans, M. New therapies for acute RSV infections: Where are we? Eur. J. Pediatr. 2019, 178, 131–138. [Google Scholar] [CrossRef]
- DeVincenzo, J.; Lambkin-Williams, R.; Wilkinson, T.; Cehelsky, J.; Nochur, S.; Walsh, E.; Meyers, R.; Gollob, J.; Vaishnaw, A. A randomized, double-blind, placebo-controlled study of an RNAi-based therapy directed against respiratory syncytial virus. Proc. Natl. Acad. Sci. USA 2010, 107, 8800–8805. [Google Scholar] [CrossRef] [Green Version]
- Zamora, M.R.; Budev, M.; Rolfe, M.; Gottlieb, J.; DeVincenzo, J.; Cehelsky, J.; Albert, G.; Gollob, J.; Nochur, S.; Vaishnaw, A.; et al. 41: Results of a randomized phase 2 trial of ALN-RSV01, an RNAi therapeutic, lung transplant (LTX) patients infected with respiratory syncytial virus (RSV). J. Heart Lung Transplant. 2010, 29, S20. [Google Scholar] [CrossRef]
- Simon, A.; Karsten, V.; Cehelsky, J.; Shah, S.; Gollob, J.; Meyers, R.; Vaishnaw, A.; Glanville, A.; Zamora, M.; DeVincenzo, J.; et al. Results of a phase 2b multi-center trial of ALN-RSV01 in respiratory syncytial virus (RSV)-infected lung transplant patients. Eur. Respir. J. 2012, 40, P1476. [Google Scholar]
- Sajid, M.I.; Moazzam, M.; Cho, Y.; Kato, S.; Xu, A.; Way, J.J.; Lohan, S.; Tiwari, R.K. siRNA therapeutics for the therapy of COVID-19 and other coronaviruses. Mol. Pharm. 2021, 18, 2105–2121. [Google Scholar] [CrossRef]
- Khaitov, M.; Nikonova, A.; Shilovskiy, I.; Kozhikhova, K.; Kofiadi, I.; Vishnyakova, L.; Nikolskii, A.; Gattinger, P.; Kovchina, V.; Barvinskaia, E.; et al. Silencing of SARS-CoV-2 with modified siRNA-peptide dendrimer formulation. Allergy 2021, 76, 2840–2854. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.P.; Dhapte-Pawar, V.S. Novel and evolving therapies for COVID-19 related pulmonary complications. Am. J. Med. Sci. 2021, 361, 557–566. [Google Scholar] [CrossRef]
- Negi, C.K.; Babica, P.; Bajard, L.; Bienertova-Vasku, J.; Tarantino, G. Insights into the molecular targets and emerging pharmacotherapeutic interventions for nonalcoholic fatty liver disease. Metabolism 2022, 126, 154925:1–154925:18. [Google Scholar] [CrossRef]
- Weigert, A.; Martin-Higueras, C.; Hoppe, B. Novel therapeutic approaches in primary hyperoxaluria. Expert Opin. Emerg. Drugs 2018, 23, 349–357. [Google Scholar] [CrossRef]
- Habtemariam, B.A.; Karsten, V.; Attarwala, H.; Goel, V.; Melch, M.; Clausen, V.A.; Garg, P.; Vaishnaw, A.K.; Sweetser, M.T.; Robbie, G.J.; et al. Single dose pharmacokinetics and pharmacodynamics of transthyretin targeting GalNAc-siRNA conjugate, vutrisiran, in healthy subjects. Clin. Pharmacol. Ther. 2020, 109, 349–357. [Google Scholar]
- Shah, V.N.; Pyle, L. Lumasiran, an RNAi therapeutic for primary hyperoxaluria type 1. N. Engl. J. Med. 2021, 384, 1216–1226. [Google Scholar]
- Suarez, A.A.R.; Testoni, B.; Zoulim, F. HBV 2021: New therapeutic strategies against an old foe. Liver Int. 2021, 41, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Warden, B.A.; Duell, P.B. Inclisiran: A novel agent for lowering apolipoprotein B–containing lipoproteins. J. Cardiovasc. Pharmacol. 2021, 78, e157–e174. [Google Scholar] [CrossRef] [PubMed]
- Wooddell, C.I.; Blomenkamp, K.; Peterson, R.M.; Subbotin, V.M.; Schwabe, C.; Hamilton, J.; Chu, Q.; Christianson, D.R.; Hegge, J.O.; Kolbe, J.; et al. Development of an RNAi therapeutic for alpha-1-antitrypsin liver disease. JCI Insight 2020, 5, e135348:1–e135348:19. [Google Scholar] [CrossRef]
- Dong, J.; Ying, J.; Qiu, X.; Lu, Y.; Zhang, M. Advanced strategies for eliminating the cccDNA of HBV. Dig. Dis. Sci. 2018, 63, 7–15. [Google Scholar] [CrossRef]
- Weigert, A.; Hoppe, B. Nephrolithiasis and nephrocalcinosis in childhood—Risk factor-related current and future treatment options. Front. Pediatr. 2018, 6, 00098:1–00098:9. [Google Scholar] [CrossRef]
- Adams, D.; Verena, K. Phase 1 study of ALN-TTRsc02, a subcutaneously administered investigational RNAi therapeutic for the treatment of transthyretin-mediated amyloidosis. Rev. Neurol. 2019, 175, S129. [Google Scholar] [CrossRef]
- Yuen, M.-F.; Schiefke, I.; Yoon, J.-H.; Ahn, S.H.; Heo, J.; Kim, J.H.; Chan, H.L.Y.; Yoon, K.T.; Klinker, H.; Manns, M.; et al. RNA interference therapy with ARC-520 results in prolonged hepatitis B surface antigen response in patients with chronic hepatitis B infection. Hepatology 2020, 72, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Badri, P.; Jiang, X.; Borodovsky, A.; Najafian, N.; Kim, J.; Clausen, V.A.; Goel, V.; Habtemariam, B.; Robbie, G.J. Pharmacokinetic and pharmacodynamic properties of cemdisiran, an RNAi therapeutic targeting complement component 5, in healthy subjects and patients with paroxysmal nocturnal hemoglobinuria. Clin. Pharmacokinet. 2021, 60, 365–378. [Google Scholar] [CrossRef]
- JohnPasi, K.; Georgiev, P.; Mant, T.; Lissitchkov, T.; Creagh, M.D.; Bevan, D.; Austin, S.; RHay, C.; Hegemann, I.; Kazmi, R.; et al. Fitusiran, an investigational RNAi therapeutic targeting antithrombin for the treatment of hemophilia: Updated results from a phase 1 and phase 1/2 extension study in patients with inhibitors. Blood 2016, 128, 1397:1–1397:3. [Google Scholar]
- Sardh, E.; Harper, P.; Balwani, M.; Stein, P.; Rees, D.; Bissell, D.M.; Desnick, R.; Parker, C.; Phillips, J.; Bonkovsky, H.L.; et al. Phase 1 trial of an RNA interference therapy for acute intermittent porphyria. N. Engl. J. Med. 2019, 380, 549–558. [Google Scholar] [CrossRef]
- Hoy, S.M. Patisiran: First global approval. Drugs 2018, 78, 1625–1631. [Google Scholar] [CrossRef]
- Fitzgerald, K.; White, S.; Borodovsky, A.; Bettencourt, B.R.; Strahs, A.; Clausen, V.; Wijngaard, P.; Horton, J.D.; Taubel, J.; Brooks, A.; et al. A highly durable RNAi therapeutic inhibitor of PCSK9. N. Engl. J. Med. 2017, 376, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, X.; Gatti, P.; Papoian, T. Safety of antisense oligonucleotide and siRNA-based therapeutics. Drug Discov. Today 2017, 22, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Benitez-Del-Castillo, J.M.; Moreno-Montañés, J.; Jiménez-Alfaro, I.; Muñoz-Negrete, F.J.; Turman, K.; Palumaa, K.; Sádaba, B.; González, M.V.; Ruz, V.; Vargas, B.; et al. Safety and efficacy clinical trials for SYL1001, a novel short interfering RNA for the treatment of dry eye disease. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6447–6454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beigel, J.H.; Voell, J.; Muñoz, P.; Kumar, P.; Brooks, K.M.; Zhang, J.; Iversen, P.; Heald, A.; Wong, M.; Davey, R.T. Safety, tolerability, and pharmacokinetics of radavirsen (AVI-7100), an antisense oligonucleotide targeting influenza a M1/M2 translation. Br. J. Clin. Pharmacol. 2018, 84, 25–34. [Google Scholar] [CrossRef] [Green Version]







| Disease | Target | Administration | Delivery System | Ref. |
|---|---|---|---|---|
| MDR-lung cancer | ABCC3 | Inhalation | LPNs | [17] |
| Lung adenocarcinoma | VEGF | Inhalation | UCNP | [18] |
| Lung cancer | TUBB3 | Oro-tracheal administration | NPs | [19] |
| Lung cancer | MRP1, BCL2 | Inhalation | NLC | [20] |
| Lung cancer | Akt1 | Inhalation | Nanosized polymer | [21] |
| Lung cancer | Mcl1 | Intratracheal Instillation | Nanoliposomes | [22] |
| Lung metastasis | STAT3 | Inhalation | PFC | [23] |
| NSCLC | EGFR-TKs | Inhalation | NLC | [15] |
| NSCLC | MRP1, BCL2 | Inhalation | MSN | [24] |
| Delivery Systems | Characteristics | |
|---|---|---|
| Advantages | Disadvantages | |
| Liposome |
|
|
| Solid lipid nanoparticle |
|
|
| Nanostructured lipid carrier |
|
|
| Nanoemulsion |
|
|
| Polymeric micelle |
|
|
| Polymer nanocapsule |
|
|
| Nanosphere |
|
|
| Dendrimer |
|
|
| Pulmonary surfactant |
| / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Y.; Yang, Z. Inhaled siRNA Formulations for Respiratory Diseases: From Basic Research to Clinical Application. Pharmaceutics 2022, 14, 1193. https://doi.org/10.3390/pharmaceutics14061193
Fan Y, Yang Z. Inhaled siRNA Formulations for Respiratory Diseases: From Basic Research to Clinical Application. Pharmaceutics. 2022; 14(6):1193. https://doi.org/10.3390/pharmaceutics14061193
Chicago/Turabian StyleFan, Yulin, and Zhijun Yang. 2022. "Inhaled siRNA Formulations for Respiratory Diseases: From Basic Research to Clinical Application" Pharmaceutics 14, no. 6: 1193. https://doi.org/10.3390/pharmaceutics14061193
APA StyleFan, Y., & Yang, Z. (2022). Inhaled siRNA Formulations for Respiratory Diseases: From Basic Research to Clinical Application. Pharmaceutics, 14(6), 1193. https://doi.org/10.3390/pharmaceutics14061193