
Common Structural Pattern for Flecainide Binding in Atrial-Selective Kv1.5 and Nav1.5 Channels: A Computational Approach

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
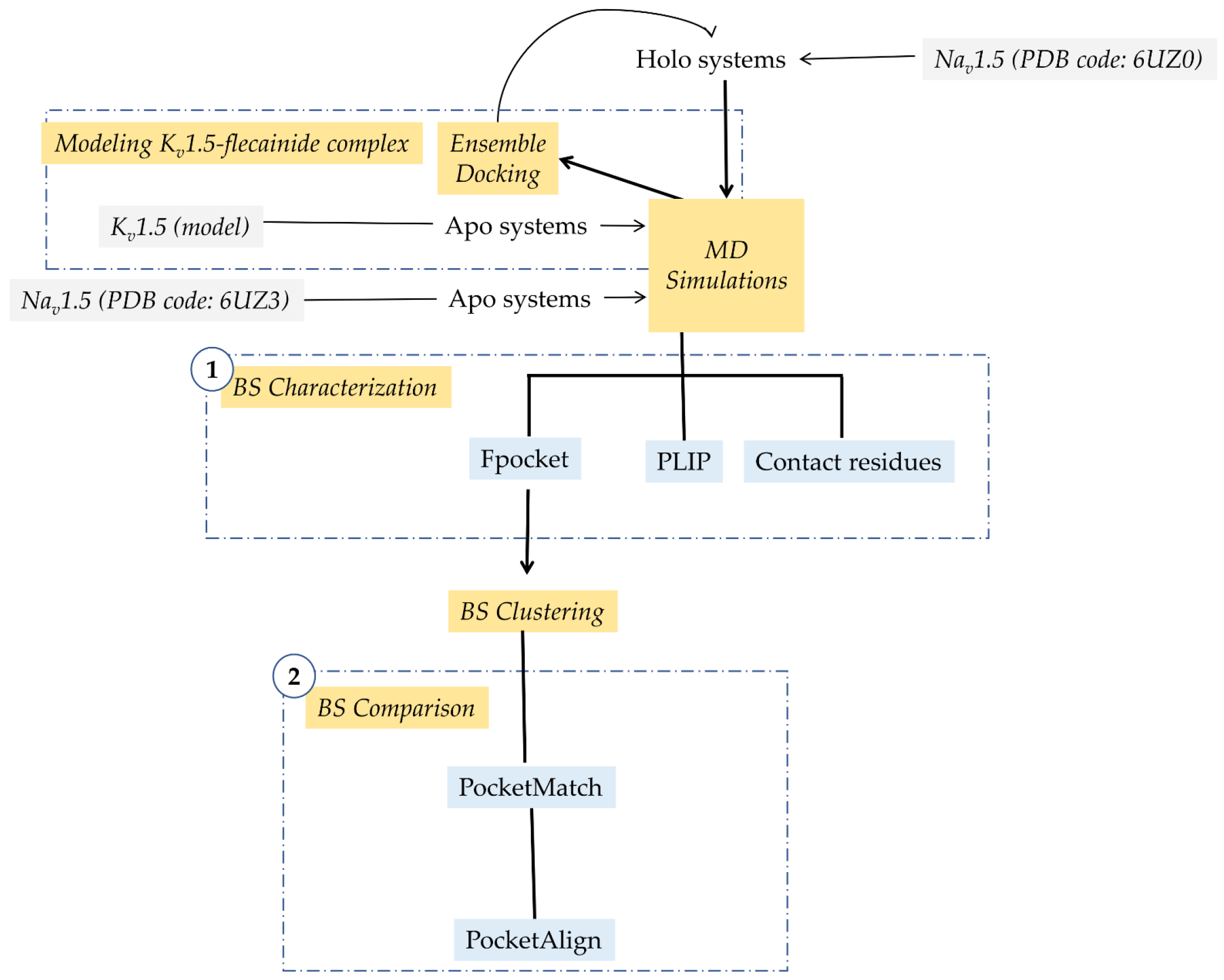
2. Materials and Methods
2.1. Modeling Flecainide-Kv1.5 Complex
2.2. Setting up Ion Channel Systems
2.3. Flecainide Binding Site Characterization
2.4. Flecainide Binding Site Comparison
2.5. Statistical Tests
3. Results
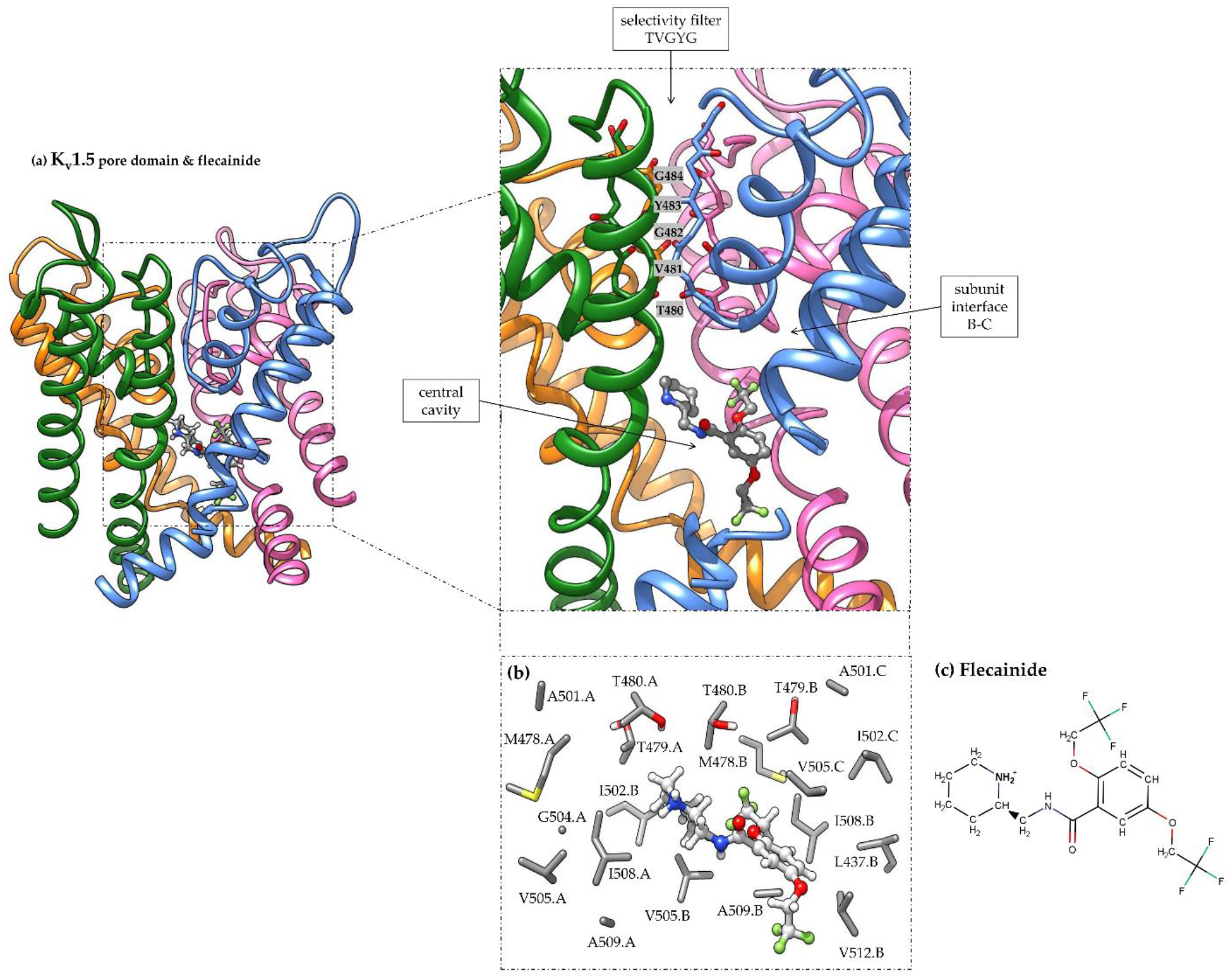
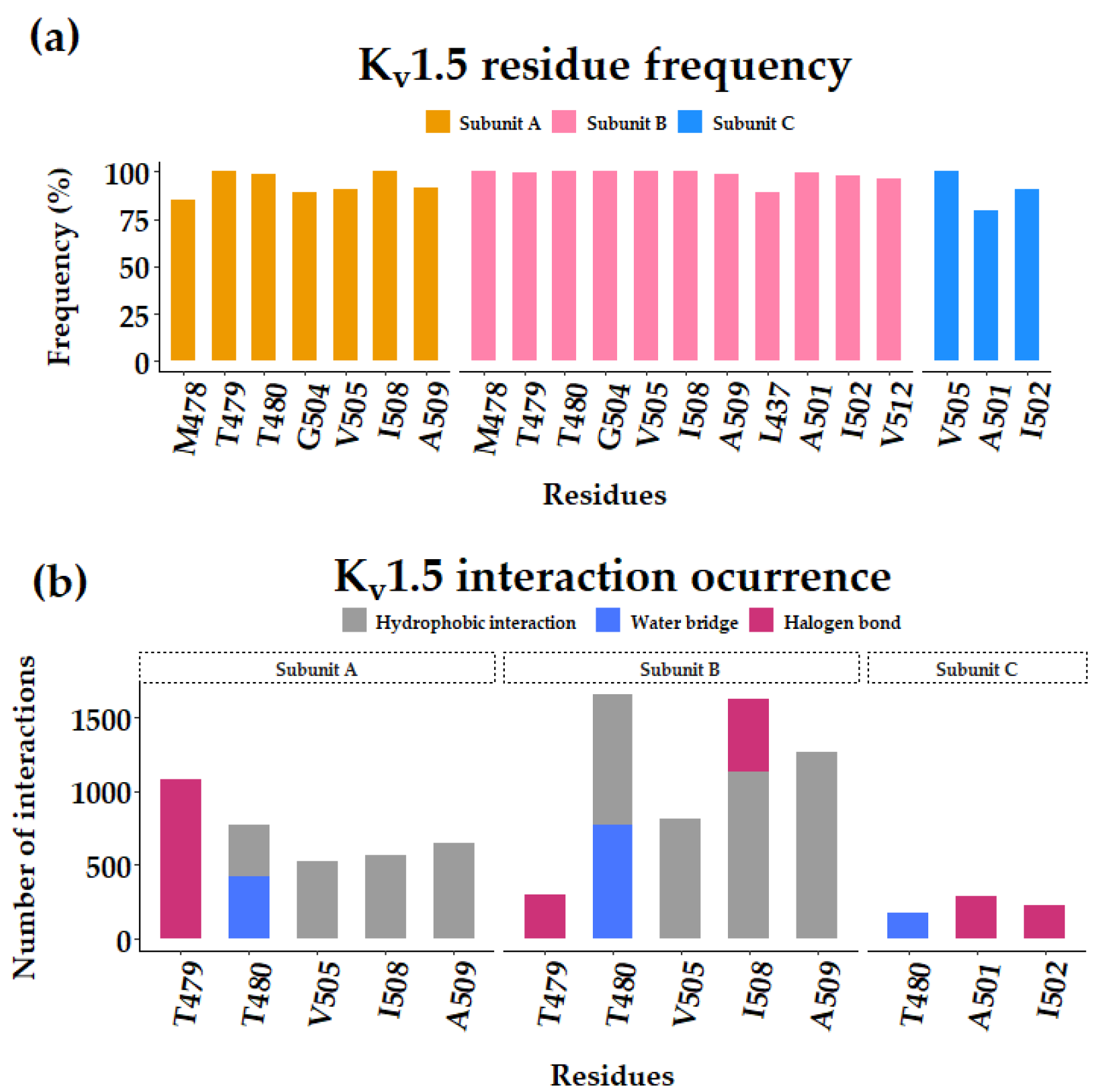
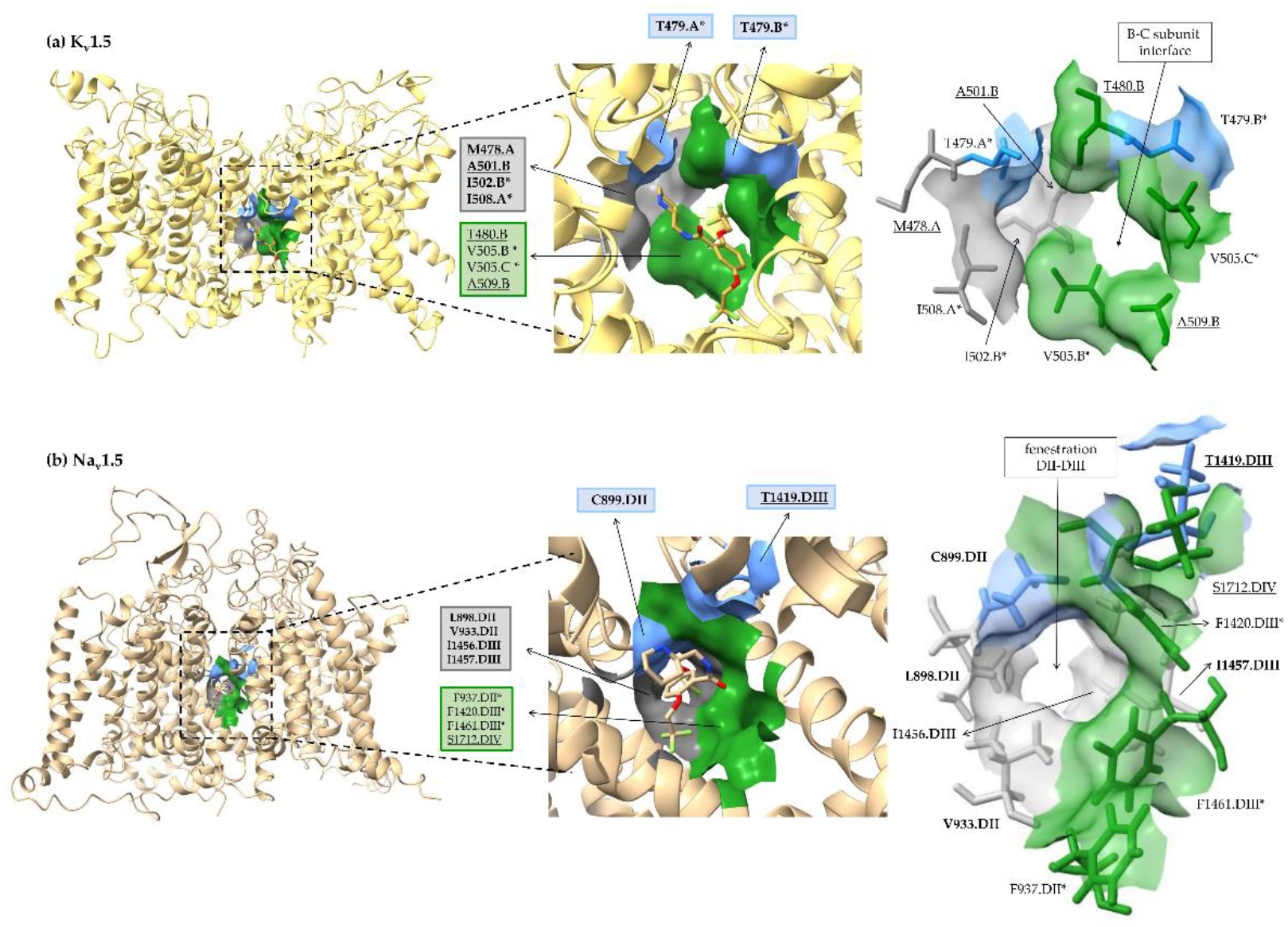
3.1. Flecainide Binding Mode and Interactions in Kv1.5
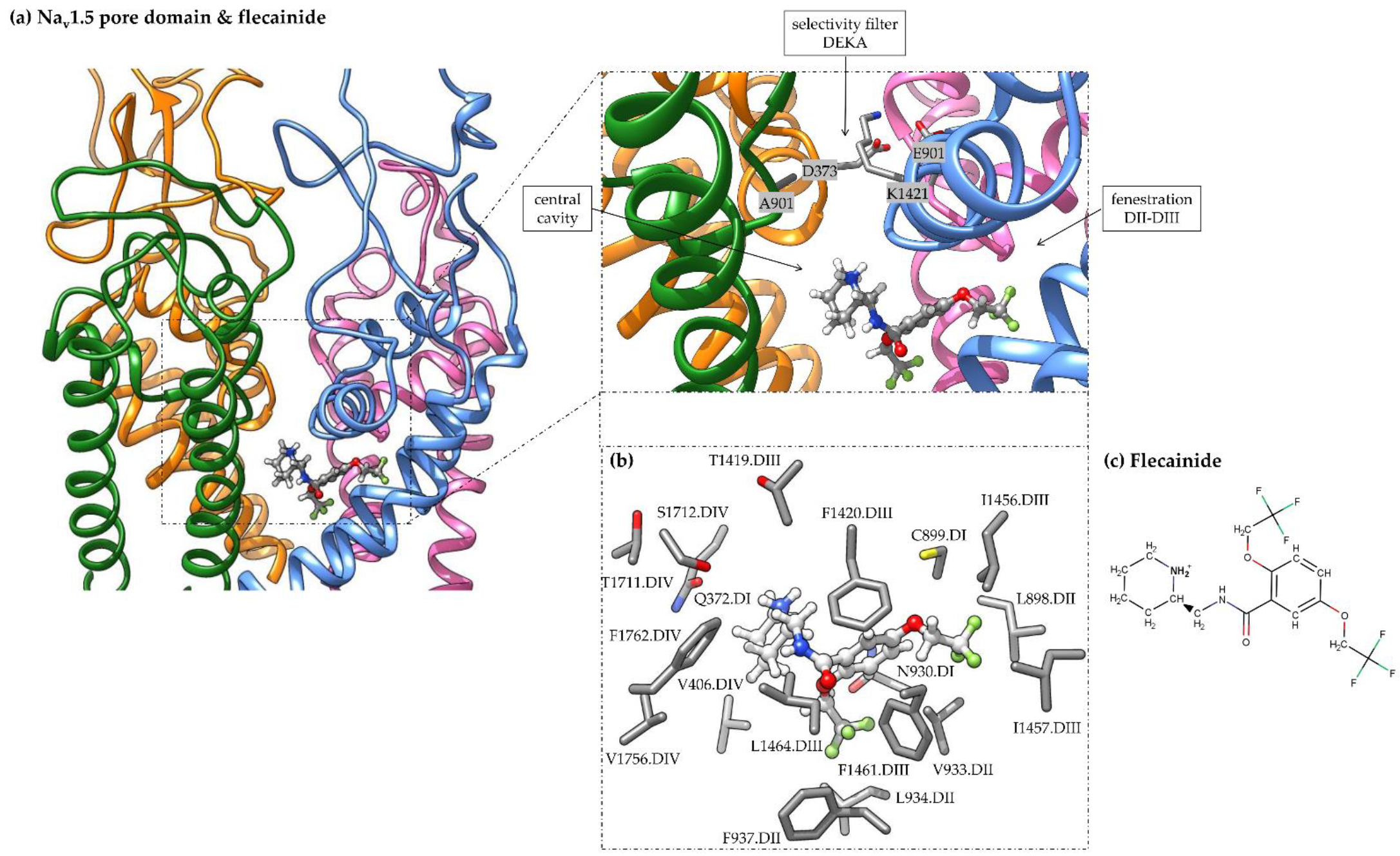
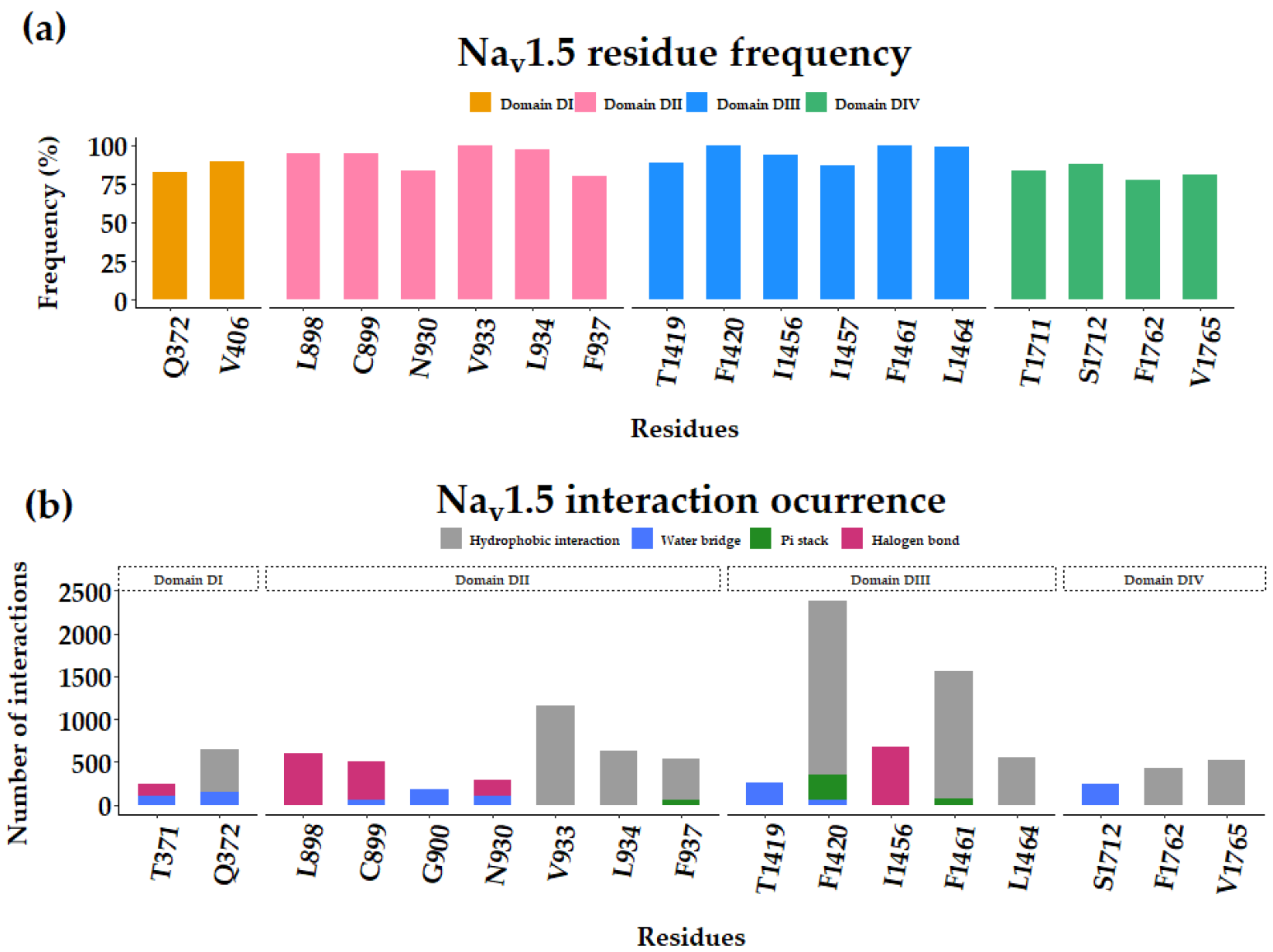
3.2. Flecainide Binding Mode and Interactions in Nav1.5
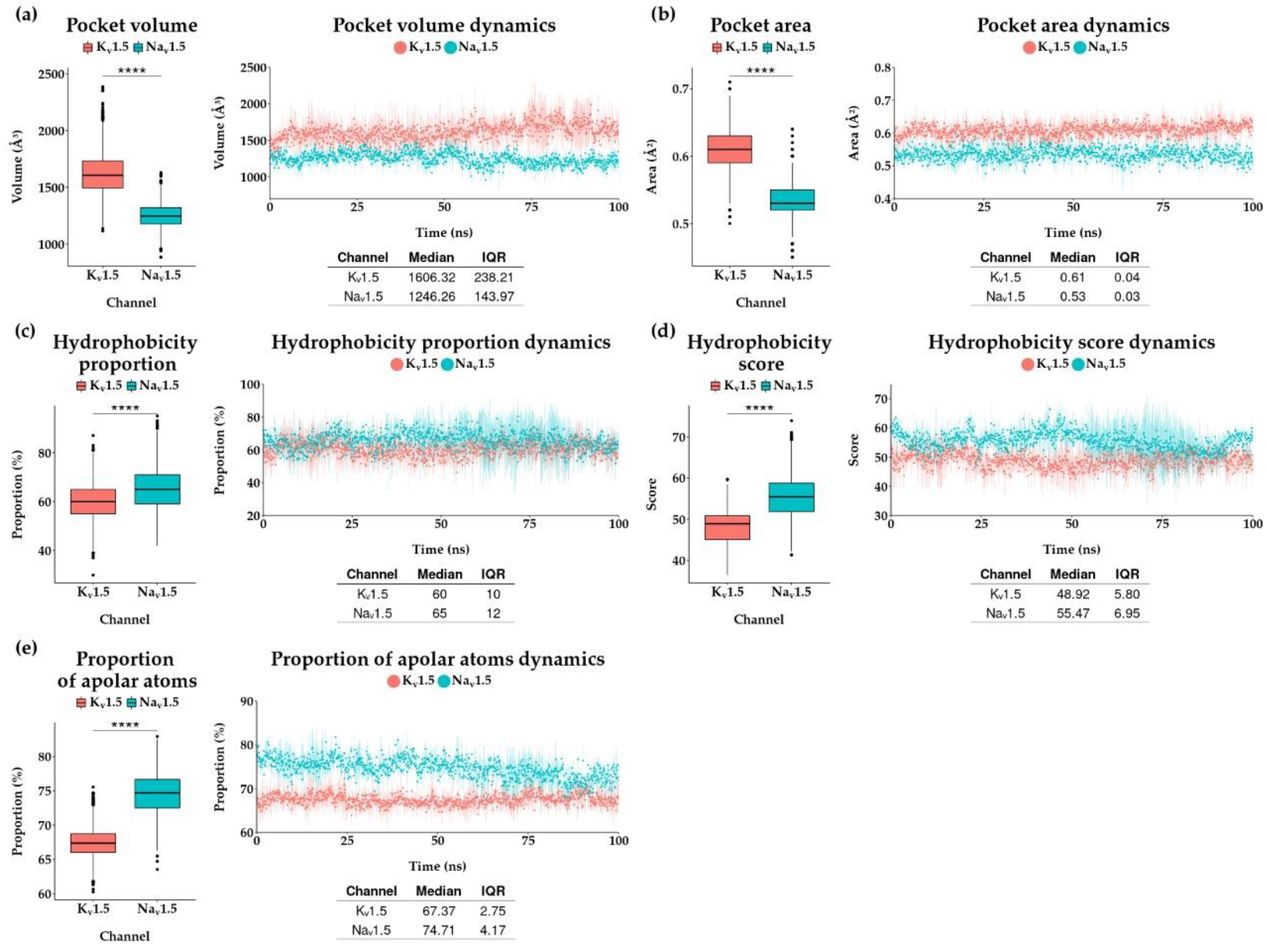
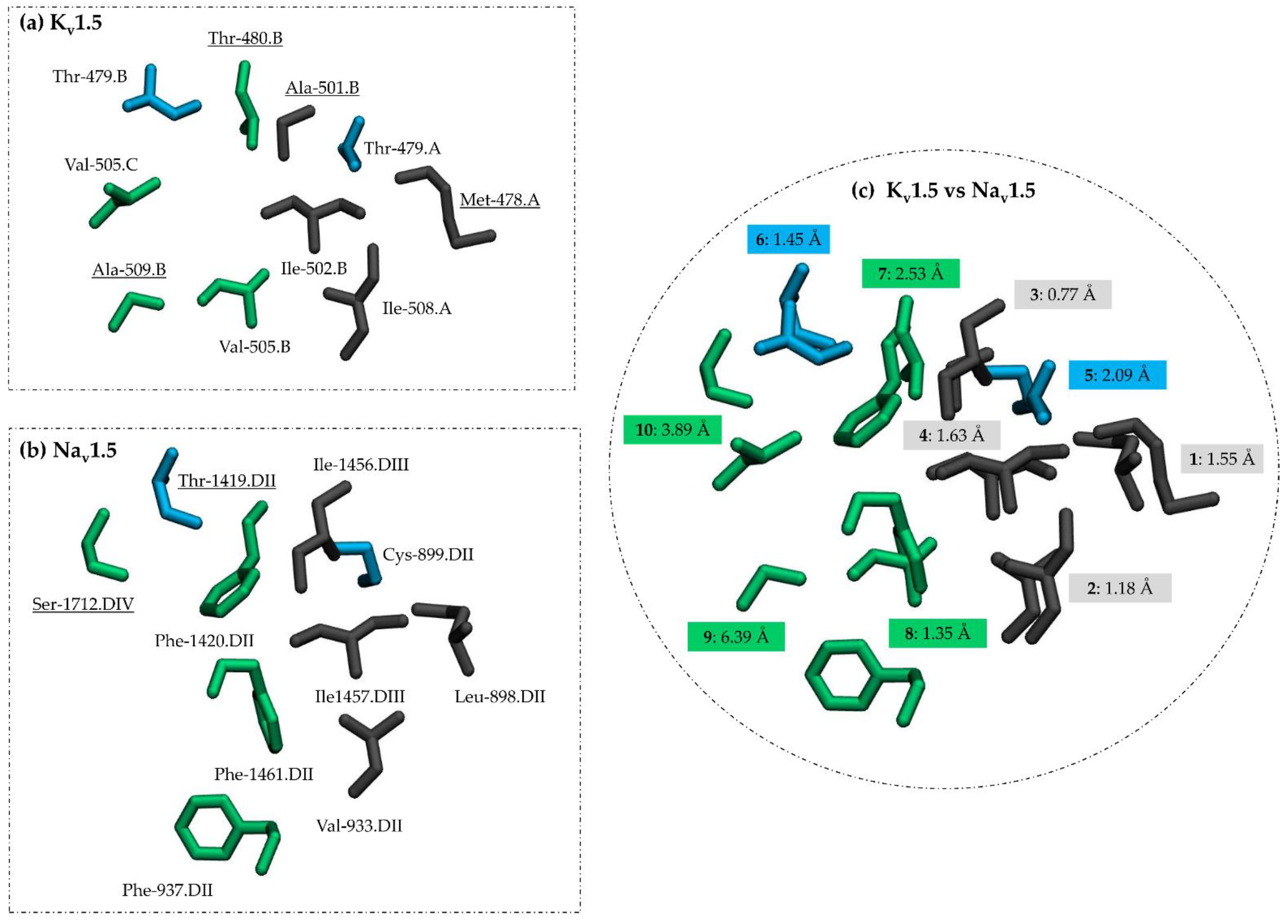
3.3. Comparing Flecainide Binding Site
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Chugh, S.S.; Havmoeller, R.; Narayanan, K.; Singh, D.; Rienstra, M.; Benjamin, E.J.; Gillum, R.F.; Kim, Y.-H.; McAnulty, J.H.; Zheng, Z.-J.; et al. Worldwide Epidemiology of Atrial Fibrillation: A Global Burden of Disease 2010 Study. Circulation 2014, 129, 837–847. [Google Scholar] [CrossRef] [Green Version]
- Geng, M.; Lin, A.; Nguyen, T.P. Revisiting Antiarrhythmic Drug Therapy for Atrial Fibrillation: Reviewing Lessons Learned and Redefining Therapeutic Paradigms. Front. Pharmacol. 2020, 11, 581837. [Google Scholar] [CrossRef] [PubMed]
- Peyronnet, R.; Ravens, U. Atria-Selective Antiarrhythmic Drugs in Need of Alliance Partners. Pharmacol. Res. 2019, 145, 104262. [Google Scholar] [CrossRef] [PubMed]
- Ravens, U.; Poulet, C.; Wettwer, E.; Knaut, M. Atrial Selectivity of Antiarrhythmic Drugs. J. Physiol. 2013, 591, 4087–4097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antzelevitch, C.; Burashnikov, A. Atrial-Selective Sodium Channel Block as a Novel Strategy for the Management of Atrial Fibrillation. J. Electrocardiol. 2009, 42, 543–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravens, U. Atrial-Selective K+ Channel Blockers: Potential Antiarrhythmic Drugs in Atrial Fibrillation? Can. J. Physiol. Pharmacol. 2017, 95, 1313–1318. [Google Scholar] [CrossRef] [Green Version]
- Burashnikov, A.; Di Diego, J.M.; Zygmunt, A.C.; Belardinelli, L.; Antzelevitch, C. Atrial-Selective Sodium Channel Block as a Strategy for Suppression of Atrial Fibrillation. Ann. N. Y. Acad. Sci. 2008, 1123, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Ford, J.W.; Milnes, J.T. New Drugs Targeting the Cardiac Ultra-Rapid Delayed-Rectifier Current (I Kur): Rationale, Pharmacology and Evidence for Potential Therapeutic Value. J. Cardiovasc. Pharmacol. 2008, 52, 105–120. [Google Scholar] [CrossRef] [Green Version]
- Ford, J.; Milnes, J.; Wettwer, E.; Christ, T.; Rogers, M.; Sutton, K.; Madge, D.; Virag, L.; Jost, N.; Horvath, Z.; et al. Human Electrophysiological and Pharmacological Properties of XEN-D0101: A Novel Atrial-Selective Kv1.5/IKur Inhibitor. J. Cardiovasc. Pharmacol. 2013, 61, 408–415. [Google Scholar] [CrossRef] [Green Version]
- Pavri, B.B.; Greenberg, H.E.; Kraft, W.K.; Lazarus, N.; Lynch, J.J.; Salata, J.J.; Bilodeau, M.T.; Regan, C.P.; Stump, G.; Fan, L.; et al. MK-0448, a Specific Kv1.5 Inhibitor: Safety, Pharmacokinetics, and Pharmacodynamic Electrophysiology in Experimental Animal Models and Humans. Circ. Arrhythm. Electrophysiol. 2012, 5, 1193–1201. [Google Scholar] [CrossRef] [Green Version]
- Van Wagoner, D.R. Multi-Channel Blockers for Treatment of Atrial Fibrillation: An Effective Strategy? Cardiovasc. Res. 2013, 98, 5–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiffel, J.A.; Camm, A.J.; Belardinelli, L.; Zeng, D.; Karwatowska-Prokopczuk, E.; Olmsted, A.; Zareba, W.; Rosero, S.; Kowey, P. HARMONY Investigators The HARMONY Trial: Combined Ranolazine and Dronedarone in the Management of Paroxysmal Atrial Fibrillation: Mechanistic and Therapeutic Synergism. Circ. Arrhythm. Electrophysiol. 2015, 8, 1048–1056. [Google Scholar] [CrossRef]
- Koskinas, K.C.; Fragakis, N.; Katritsis, D.; Skeberis, V.; Vassilikos, V. Ranolazine Enhances the Efficacy of Amiodarone for Conversion of Recent-Onset Atrial Fibrillation. Europace 2014, 16, 973–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, M.; Xiong, F.; Qi, X.Y.; Comtois, P.; Nattel, S. Potassium Channel Blockade Enhances Atrial Fibrillation-Selective Antiarrhythmic Effects of Optimized State-Dependent Sodium Channel Blockade. Circulation 2015, 132, 2203–2211. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.; Whittaker, D.G.; Wang, W.; Giles, W.R.; Narayan, S.M.; Zhang, H. Synergistic Anti-Arrhythmic Effects in Human Atria with Combined Use of Sodium Blockers and Acacetin. Front. Physiol. 2017, 8, 946. [Google Scholar] [CrossRef] [Green Version]
- Jæger, K.H.; Edwards, A.G.; Giles, W.R.; Tveito, A. A Computational Method for Identifying an Optimal Combination of Existing Drugs to Repair the Action Potentials of SQT1 Ventricular Myocytes. PLoS Comput. Biol. 2021, 17, e1009233. [Google Scholar] [CrossRef]
- Haupt, V.J.; Daminelli, S.; Schroeder, M. Drug Promiscuity in PDB: Protein Binding Site Similarity Is Key. PLOS ONE 2013, 8, e65894. [Google Scholar] [CrossRef]
- Valdés-Jiménez, A.; Jiménez-González, D.; Kiper, A.K.; Rinné, S.; Decher, N.; González, W.; Reyes-Parada, M.; Núñez-Vivanco, G. A New Strategy for Multitarget Drug Discovery/Repositioning through the Identification of Similar 3D Amino Acid Patterns Among Proteins Structures: The Case of Tafluprost and Its Effects on Cardiac Ion Channels. Front. Pharmacol. 2022, 13, 761. [Google Scholar] [CrossRef]
- Naderi, M.; Lemoine, J.M.; Govindaraj, R.G.; Kana, O.Z.; Feinstein, W.P.; Brylinski, M. Binding Site Matching in Rational Drug Design: Algorithms and Applications. Brief. Bioinform. 2019, 20, 2167–2184. [Google Scholar] [CrossRef]
- Jiang, D.; Shi, H.; Tonggu, L.; Gamal El-Din, T.M.; Lenaeus, M.J.; Zhao, Y.; Yoshioka, C.; Zheng, N.; Catterall, W.A. Structure of the Cardiac Sodium Channel. Cell 2020, 180, 122–134.e10. [Google Scholar] [CrossRef]
- Eldstrom, J.; Wang, Z.; Xu, H.; Pourrier, M.; Ezrin, A.; Gibson, K.; Fedida, D. The Molecular Basis of High-Affinity Binding of the Antiarrhythmic Compound Vernakalant (RSD1235) to Kv1.5 Channels. Mol. Pharmacol. 2007, 72, 1522–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliot, E.; Capucci, A.; Crijns, H.J.; Goette, A.; Tamargo, J. Twenty-Five Years in the Making: Flecainide Is Safe and Effective for the Management of Atrial Fibrillation. Europace 2011, 13, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Echt, D.S.; Ruskin, J.N. Use of Flecainide for the Treatment of Atrial Fibrillation. Am. J. Cardiol. 2020, 125, 1123–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muzzey, M.; Tellor, K.B.; Ramaswamy, K.; Schwarze, M.; Armbruster, A.L. Flecainide Is Well-Tolerated and Effective in Patient with Atrial Fibrillation at 12 Months: A Retrospective Study. Ther. Adv. Cardiovasc. Dis. 2020, 14, 1753944720926824. [Google Scholar] [CrossRef] [PubMed]
- Melgari, D.; Zhang, Y.; El Harchi, A.; Dempsey, C.E.; Hancox, J.C. Molecular Basis of HERG Potassium Channel Blockade by the Class Ic Antiarrhythmic Flecainide. J. Mol. Cell. Cardiol. 2015, 86, 42–53. [Google Scholar] [CrossRef] [Green Version]
- Paul, A.A.; Witchel, H.J.; Hancox, J.C. Inhibition of the Current of Heterologously Expressed HERG Potassium Channels by Flecainide and Comparison with Quinidine, Propafenone and Lignocaine. Br. J. Pharmacol. 2002, 136, 717–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grissmer, S.; Nguyen, A.N.; Aiyar, J.; Hanson, D.C.; Mather, R.J.; Gutman, G.A.; Karmilowicz, M.J.; Auperin, D.D.; Chandy, K.G. Pharmacological Characterization of Five Cloned Voltage-Gated K+ Channels, Types Kv1.1, 1.2, 1.3, 1.5, and 3.1, Stably Expressed in Mammalian Cell Lines. Mol. Pharmacol. 1994, 45, 1227–1234. [Google Scholar]
- Herrera, D.; Mamarbachi, A.; Simoes, M.; Parent, L.; Sauvé, R.; Wang, Z.; Nattel, S. A Single Residue in the S6 Transmembrane Domain Governs the Differential Flecainide Sensitivity of Voltage-Gated Potassium Channels. Mol. Pharmacol. 2005, 68, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Salvage, S.C.; Chandrasekharan, K.H.; Jeevaratnam, K.; Dulhunty, A.F.; Thompson, A.J.; Jackson, A.P.; Huang, C.L. Multiple Targets for Flecainide Action: Implications for Cardiac Arrhythmogenesis. Br. J. Pharmacol. 2018, 175, 1260–1278. [Google Scholar] [CrossRef] [Green Version]
- Kaminski, G.A.; Friesner, R.A.; Tirado-Rives, J.; Jorgensen, W.L. Evaluation and Reparametrization of the OPLS-AA Force Field for Proteins via Comparison with Accurate Quantum Chemical Calculations on Peptides. J. Phys. Chem. B 2001, 105, 6474–6487. [Google Scholar] [CrossRef]
- Banks, J.L.; Beard, H.S.; Cao, Y.; Cho, A.E.; Damm, W.; Farid, R.; Felts, A.K.; Halgren, T.A.; Mainz, D.T.; Maple, J.R.; et al. Integrated Modeling Program, Applied Chemical Theory (IMPACT). J. Comput. Chem. 2005, 26, 1752–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shivakumar, D.; Williams, J.; Wu, Y.; Damm, W.; Shelley, J.; Sherman, W. Prediction of Absolute Solvation Free Energies Using Molecular Dynamics Free Energy Perturbation and the OPLS Force Field. J. Chem. Theory Comput. 2010, 6, 1509–1519. [Google Scholar] [CrossRef] [PubMed]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 1. Method and Assessment of Docking Accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef] [PubMed]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra Precision Glide: Docking and Scoring Incorporating a Model of Hydrophobic Enclosure for Protein−Ligand Complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madhavi Sastry, G.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and Ligand Preparation: Parameters, Protocols, and Influence on Virtual Screening Enrichments. J. Comput. Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- Bowers, K.; Chow, E.; Xu, H.; Dror, R.O.; Eastwood, M.P. Scalable Algorithms for Molecular Dynamics Simulations on Commodity Clusters. In Proceedings of the 2006 ACM/IEEE Conference on Supercomputing, Tampa, FL, USA, 11–17 November 2006; Volume 6, pp. 7695–7700. [Google Scholar]
- Marzian, S.; Stansfeld, P.J.; Rapedius, M.; Rinné, S.; Nematian-Ardestani, E.; Abbruzzese, J.L.; Steinmeyer, K.; Sansom, M.S.P.; Sanguinetti, M.C.; Baukrowitz, T.; et al. Side Pockets Provide the Basis for a New Mechanism of Kv Channel-Specific Inhibition. Nat. Chem. Biol. 2013, 9, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.; Merz, K.M. Application of the Nosé−Hoover Chain Algorithm to the Study of Protein Dynamics. J. Phys. Chem. 1996, 100, 1927–1937. [Google Scholar] [CrossRef]
- Martyna, G.J.; Tobias, D.J.; Klein, M.L. Constant Pressure Molecular Dynamics Algorithms. J. Chem. Phys. 1994, 101, 4177–4189. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Smart, O.S.; Neduvelil, J.G.; Wang, X.; Wallace, B.A.; Sansom, M.S. HOLE: A Program for the Analysis of the Pore Dimensions of Ion Channel Structural Models. J. Mol. Graph. 1996, 14, 354–360. [Google Scholar] [CrossRef]
- Fpocket: An Open Source Platform for Ligand Pocket Detection—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/19486540/ (accessed on 17 June 2022).
- Monera, O.D.; Sereda, T.J.; Zhou, N.E.; Kay, C.M.; Hodges, R.S. Relationship of Sidechain Hydrophobicity and Alpha-Helical Propensity on the Stability of the Single-Stranded Amphipathic Alpha-Helix. J. Pept. Sci. 1995, 1, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully Automated Protein-Ligand Interaction Profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef] [PubMed]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP 2021: Expanding the Scope of the Protein-Ligand Interaction Profiler to DNA and RNA. Nucleic Acids Res. 2021, 49, W530–W534. [Google Scholar] [CrossRef] [PubMed]
- Charrad, M.; Ghazzali, N.; Boiteau, V.; Niknafs, A. NbClust: An R Package for Determining the Relevant Number of Clusters in a Data Set. J. Stat. Softw. 2014, 61, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Yeturu, K.; Chandra, N. PocketMatch: A New Algorithm to Compare Binding Sites in Protein Structures. BMC Bioinform. 2008, 9, 543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeturu, K.; Chandra, N. PocketAlign a Novel Algorithm for Aligning Binding Sites in Protein Structures. J. Chem. Inf. Model. 2011, 51, 1725–1736. [Google Scholar] [CrossRef]
- Ragsdale, D.S.; McPhee, J.C.; Scheuer, T.; Catterall, W.A. Common Molecular Determinants of Local Anesthetic, Antiarrhythmic, and Anticonvulsant Block of Voltage-Gated Na+ Channels. Proc. Natl. Acad. Sci. USA 1996, 93, 9270–9275. [Google Scholar] [CrossRef] [Green Version]
- Ragsdale, D.S.; McPhee, J.C.; Scheuer, T.; Catterall, W.A. Molecular Determinants of State-Dependent Block of Na+ Channels by Local Anesthetics. Science 1994, 265, 1724–1728. [Google Scholar] [CrossRef]
- Nguyen, P.T.; DeMarco, K.R.; Vorobyov, I.; Clancy, C.E.; Yarov-Yarovoy, V. Structural Basis for Antiarrhythmic Drug Interactions with the Human Cardiac Sodium Channel. Proc. Natl. Acad. Sci. USA 2019, 116, 2945–2954. [Google Scholar] [CrossRef] [Green Version]
- Kiper, A.K.; Bedoya, M.; Stalke, S.; Marzian, S.; Ramírez, D.; de la Cruz, A.; Peraza, D.A.; Vera-Zambrano, A.; Márquez Montesinos, J.C.E.; Arévalo Ramos, B.A.; et al. Identification of a Critical Binding Site for Local Anaesthetics in the Side Pockets of Kv 1 Channels. Br. J. Pharmacol. 2021, 178, 3034–3048. [Google Scholar] [CrossRef]
- Kiper, A.K.; Rinné, S.; Rolfes, C.; Ramírez, D.; Seebohm, G.; Netter, M.F.; González, W.; Decher, N. Kv1.5 Blockers Preferentially Inhibit TASK-1 Channels: TASK-1 as a Target against Atrial Fibrillation and Obstructive Sleep Apnea? Pflugers Arch. 2015, 467, 1081–1090. [Google Scholar] [CrossRef] [PubMed]
- Decher, N.; Pirard, B.; Bundis, F.; Peukert, S.; Baringhaus, K.-H.; Busch, A.E.; Steinmeyer, K.; Sanguinetti, M.C. Molecular Basis for Kv1.5 Channel Block: Conservation of Drug Binding Sites among Voltage-Gated K+ Channels. J. Biol. Chem. 2004, 279, 394–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heginbotham, L.; Lu, Z.; Abramson, T.; MacKinnon, R. Mutations in the K+ Channel Signature Sequence. Biophys. J. 1994, 66, 1061–1067. [Google Scholar] [CrossRef] [Green Version]
- Yue, L.; Feng, J.L.; Wang, Z.; Nattel, S. Effects of Ambasilide, Quinidine, Flecainide and Verapamil on Ultra-Rapid Delayed Rectifier Potassium Currents in Canine Atrial Myocytes. Cardiovasc. Res. 2000, 46, 151–161. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Atkins, J.; Kass, R.S. Common Molecular Determinants of Flecainide and Lidocaine Block of Heart Na+ Channels: Evidence from Experiments with Neutral and Quaternary Flecainide Analogues. J. Gen. Physiol. 2003, 121, 199–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clustering Molecular Dynamics Trajectories for Optimizing Docking Experiments. Available online: https://www.hindawi.com/journals/cin/2015/916240/ (accessed on 18 February 2022).
- Tikhonov, D.B.; Zhorov, B.S. Homology Modeling of Kv1.5 Channel Block by Cationic and Electroneutral Ligands. Biochim. Biophys. Acta (BBA)-Biomembr. 2014, 1838, 978–987. [Google Scholar] [CrossRef] [Green Version]
- Tikhonov, D.B.; Zhorov, B.S. Mechanism of Sodium Channel Block by Local Anesthetics, Antiarrhythmics, and Anticonvulsants. J. Gen. Physiol. 2017, 149, 465–481. [Google Scholar] [CrossRef]
- Ahern, C.A.; Eastwood, A.L.; Dougherty, D.A.; Horn, R. Electrostatic Contributions of Aromatic Residues in the Local Anesthetic Receptor of Voltage-Gated Sodium Channels. Circ. Res. 2008, 102, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Pless, S.A.; Galpin, J.D.; Frankel, A.; Ahern, C.A. Molecular Basis for Class Ib Anti-Arrhythmic Inhibition of Cardiac Sodium Channels. Nat. Commun. 2011, 2, 351. [Google Scholar] [CrossRef]
- Blunck, R.; Batulan, Z. Mechanism of Electromechanical Coupling in Voltage-Gated Potassium Channels. Front. Pharmacol. 2012, 3, 166. [Google Scholar] [CrossRef] [Green Version]
- Ehrt, C.; Brinkjost, T.; Koch, O. Binding Site Characterization - Similarity, Promiscuity, and Druggability. Medchemcomm 2019, 10, 1145–1159. [Google Scholar] [CrossRef] [PubMed]
- Ferreira de Freitas, R.; Schapira, M. A Systematic Analysis of Atomic Protein-Ligand Interactions in the PDB. Medchemcomm 2017, 8, 1970–1981. [Google Scholar] [CrossRef] [Green Version]
- Darby, J.F.; Hopkins, A.P.; Shimizu, S.; Roberts, S.M.; Brannigan, J.A.; Turkenburg, J.P.; Thomas, G.H.; Hubbard, R.E.; Fischer, M. Water Networks Can Determine the Affinity of Ligand Binding to Proteins. J. Am. Chem. Soc. 2019, 141, 15818–15826. [Google Scholar] [CrossRef] [PubMed]
- Rusinova, R.; Koeppe, R.E.; Andersen, O.S. A General Mechanism for Drug Promiscuity: Studies with Amiodarone and Other Antiarrhythmics. J. Gen. Physiol. 2015, 146, 463–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unique Kv1.5_c1 Residue | Common Residues | Unique Kv1.5_c2 Residues |
|---|---|---|
| PRO513.B | MET478.A | GLY504.A |
| VAL516.B | THR479.A | VAL505.A |
| LEU506.C | THR480.A | ALA509.A |
| ALA509.C | VAL481.A | LEU437.B |
| VAL512.C | ILE508.A | LEU441.B |
| MET478.B | ALA501.C | |
| THR479.B | ILE502.C | |
| THR480.B | VAL512.D | |
| ALA501.B | ||
| ILE502.B | ||
| GLY504.B | ||
| VAL505.B | ||
| ILE508.B | ||
| ALA509.B | ||
| VAL512.B | ||
| VAL505.C |
| Unique Nav1.5_c1 Residues | Common Residues | Unique Nav1.5_c2 Residues |
|---|---|---|
| TRP375.DI | THR371.DI | LEU410A.DI |
| PHE403.DI | GLN372.DI | LEU936.DII |
| SER402.DI | VAL406.DI | MET370.DI |
| ALA1418.DIII | LEU847.DII | PHE895.DII |
| LYS1421.DIII | LEU898.DII | PHE937.DII |
| THR1711.DIV | CYS899.DII | |
| ALA1713.DIV | GLY900.DII | |
| MET926.DII | ||
| ASN930.DII | ||
| VAL933.DII | ||
| LEU934.DII | ||
| THR1419.DIII | ||
| PHE1420.DIII | ||
| ILE1456.DIII | ||
| ILE1457.DIII | ||
| PHE1461.DIII | ||
| LEU1464.DIII | ||
| SER1712.DIV | ||
| PHE1762.DIV | ||
| VAL1765.DIV |
| Centroid A | Centroid B | Similarity Score (%) |
|---|---|---|
| Kv1.5_c2 | Kv1.5_c1 | 95.2075 |
| Kv1.5_c2 | Nav1.5_c1 | 72.2969 |
| Kv1.5_c2 | Nav1.5_c2 | 78.6090 |
| Kv1.5_c1 | Nav1.5_c1 | 60.6061 |
| Kv1.5_c1 | Nav1.5_c2 | 65.1748 |
| Nav1.5_c1 | Nav1.5_c2 | 83.2655 |
| No. | Kv1.5_c2 | Physicochemical Nature/Int | Nav1.5_c2 | Physicochemical Nature/Int |
|---|---|---|---|---|
| 1 | MET478A | a | LEU898.DII * | a/halogen bonds |
| 2 | ILE508A * | a/hydrophobic int. | VAL933.DII * | a/hydrophobic int. |
| 3 | ALA501B | a | ILE1456.DIII * | a/halogen bonds |
| 4 | ILE502B * | a | ILE1457.DIII * | a |
| 5 | THR479A * | b/halogen bonds | CYS899.DII * | b/halogen bonds |
| 6 | THR479B * | b/halogen bonds | THR1419.DII | b/water bridges |
| 7 | THR480B | b/hydrophobic int., water bridges | PHE1420.DII * | c/hydrophobic int., π stacks |
| 8 | VAL505B * | a/hydrophobic int. | PHE1461.DII * | c/hydrophobic int., π stacks |
| 9 | ALA509B | a/hydrophobic int. | PHE937.DII * | c/hydrophobic int., π stacks |
| 10 | VAL505C * | a | SER1712.DIV | b/water bridges |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazola, Y.; Márquez Montesinos, J.C.E.; Ramírez, D.; Zúñiga, L.; Decher, N.; Ravens, U.; Yarov-Yarovoy, V.; González, W. Common Structural Pattern for Flecainide Binding in Atrial-Selective Kv1.5 and Nav1.5 Channels: A Computational Approach. Pharmaceutics 2022, 14, 1356. https://doi.org/10.3390/pharmaceutics14071356
Mazola Y, Márquez Montesinos JCE, Ramírez D, Zúñiga L, Decher N, Ravens U, Yarov-Yarovoy V, González W. Common Structural Pattern for Flecainide Binding in Atrial-Selective Kv1.5 and Nav1.5 Channels: A Computational Approach. Pharmaceutics. 2022; 14(7):1356. https://doi.org/10.3390/pharmaceutics14071356
Chicago/Turabian StyleMazola, Yuliet, José C. E. Márquez Montesinos, David Ramírez, Leandro Zúñiga, Niels Decher, Ursula Ravens, Vladimir Yarov-Yarovoy, and Wendy González. 2022. "Common Structural Pattern for Flecainide Binding in Atrial-Selective Kv1.5 and Nav1.5 Channels: A Computational Approach" Pharmaceutics 14, no. 7: 1356. https://doi.org/10.3390/pharmaceutics14071356
APA StyleMazola, Y., Márquez Montesinos, J. C. E., Ramírez, D., Zúñiga, L., Decher, N., Ravens, U., Yarov-Yarovoy, V., & González, W. (2022). Common Structural Pattern for Flecainide Binding in Atrial-Selective Kv1.5 and Nav1.5 Channels: A Computational Approach. Pharmaceutics, 14(7), 1356. https://doi.org/10.3390/pharmaceutics14071356