
Safety and Prophylactic Efficacy of Liposome-Based Vaccine against the Drug-Resistant Acinetobacter baumannii in Mice





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Mice
2.3. Preparation of Whole Cell Antigens (WCAgs) from A. baumannii
2.4. Preparation and Characterization of Liposomes
2.5. Mice
2.6. Immunization of Mice with Liposome-Encapsulated Acinetobacter baumannii Whole Cell Antigens (WCAgs)
- PBS
- Sham Liposomes
- Free WCAgs
- IFA-WCAgs
- Lip-WCAgs
2.7. Evaluation of the Safety of Vaccine Formulations
2.8. Determination of the IgG Titer and IgG Isotyping
2.9. Determination of the Proliferation of Lymphocytes
2.10. Analysis of Cytokines
2.11. Determination of the Opsonophagocytic Killing Activity of Serum
2.12. Determination of the Effect of Antisera on the Biofilm Formation by A. baumannii
2.13. Mouse Model of A. baumannii Infection
2.14. Assessment of the Prophylactic Potential of Vaccine Formulations against A. baumannii
2.15. Determination of Inflammation Markers in the Systemic Circulation
2.16. Statistical Analyses
3. Results
3.1. PDI, Size, and Zeta Potential of Liposomes
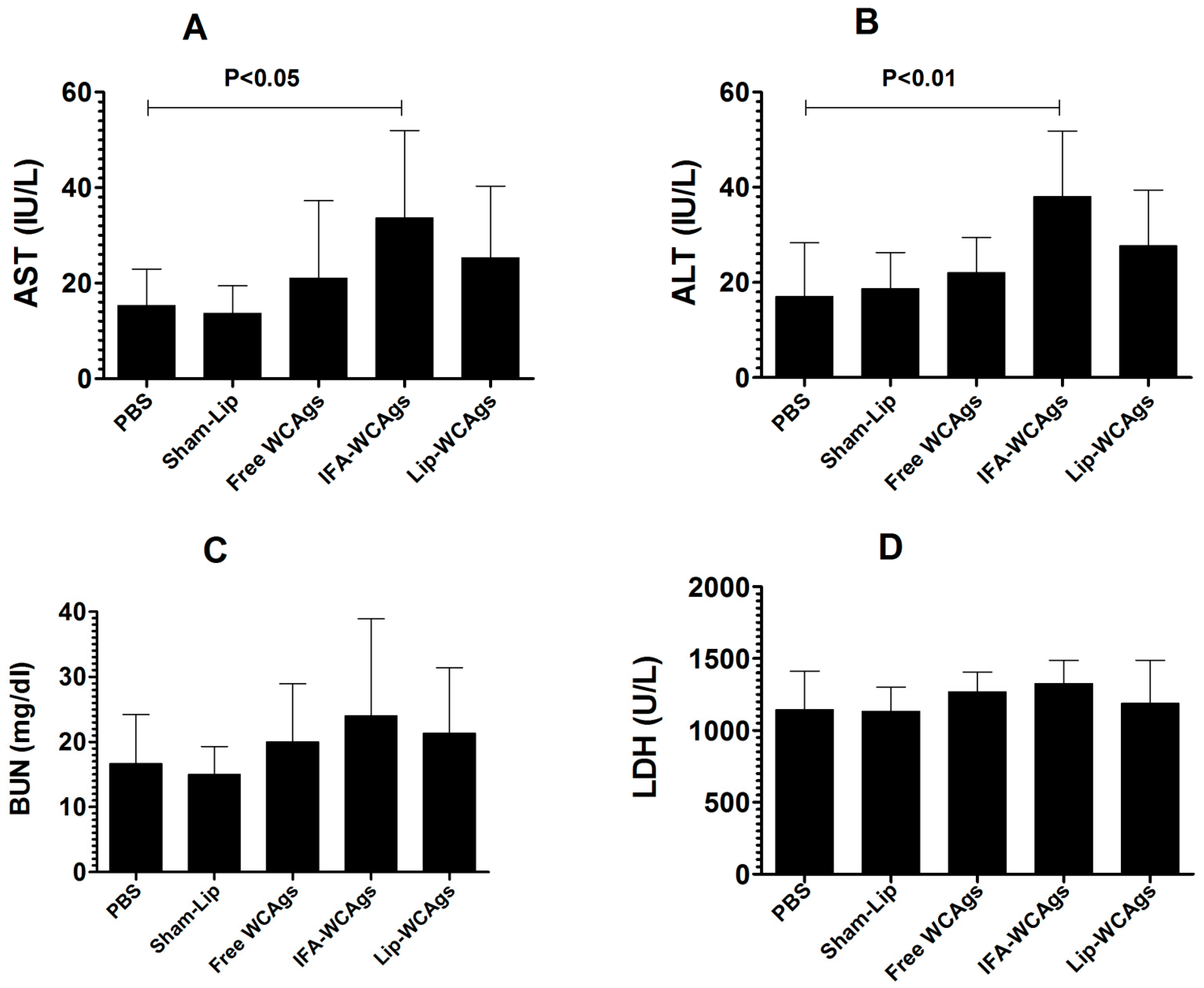
3.2. Immunization with Lip-WCAgs Did Not Induce Any Remarkable Toxicity in Mice
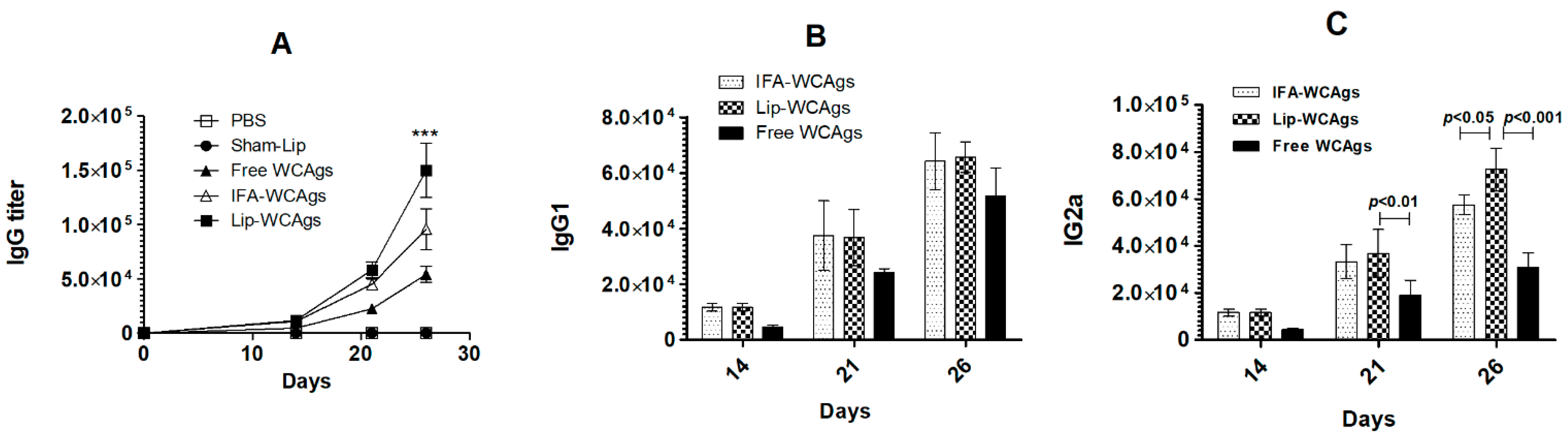
3.3. Immunization with Lip-WCAgs Induced the Greater the Secretion of Total IgG and IgG2a
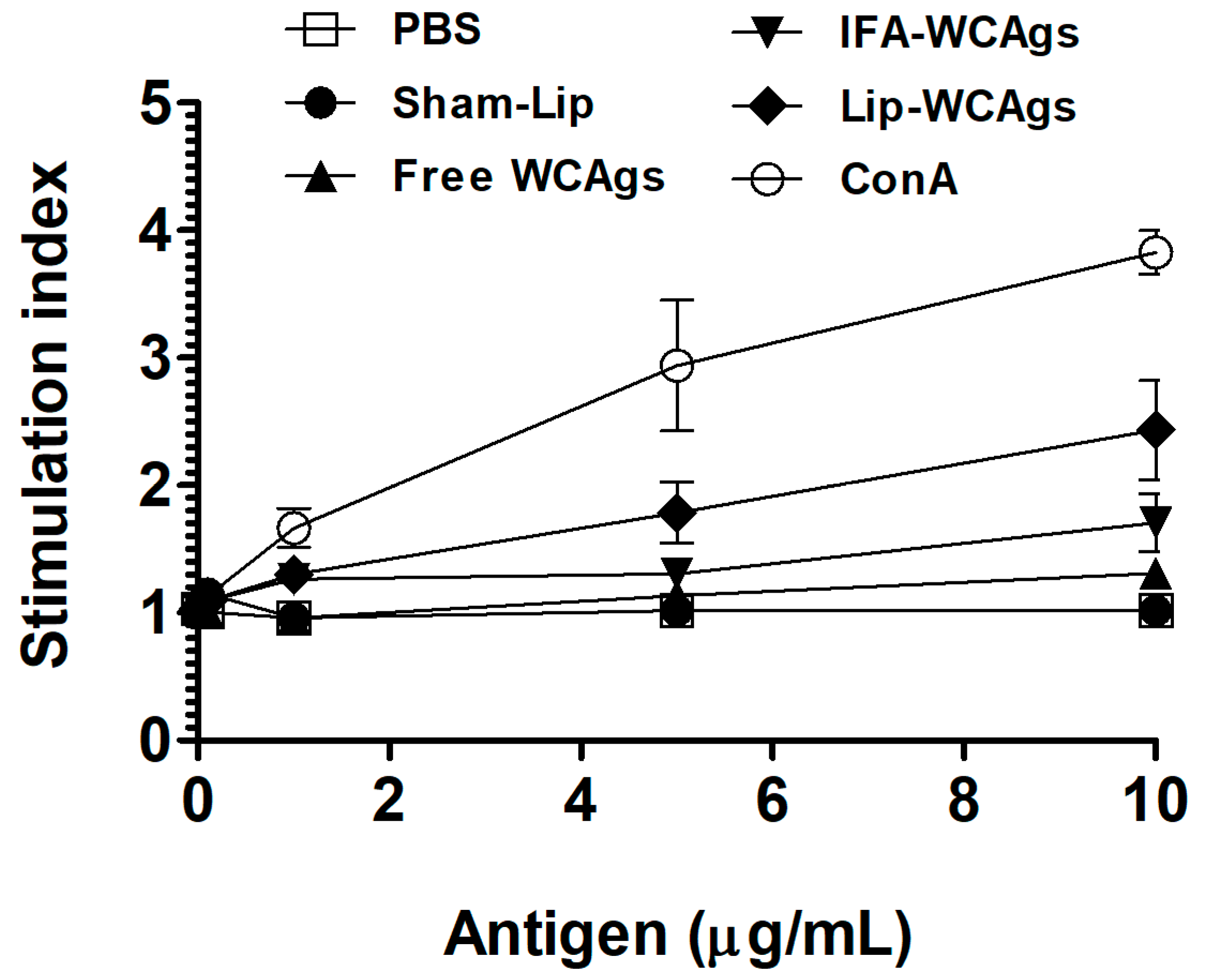
3.4. Splenocytes from Mice Immunized with Lip-WCAgs Showed Greater Proliferation
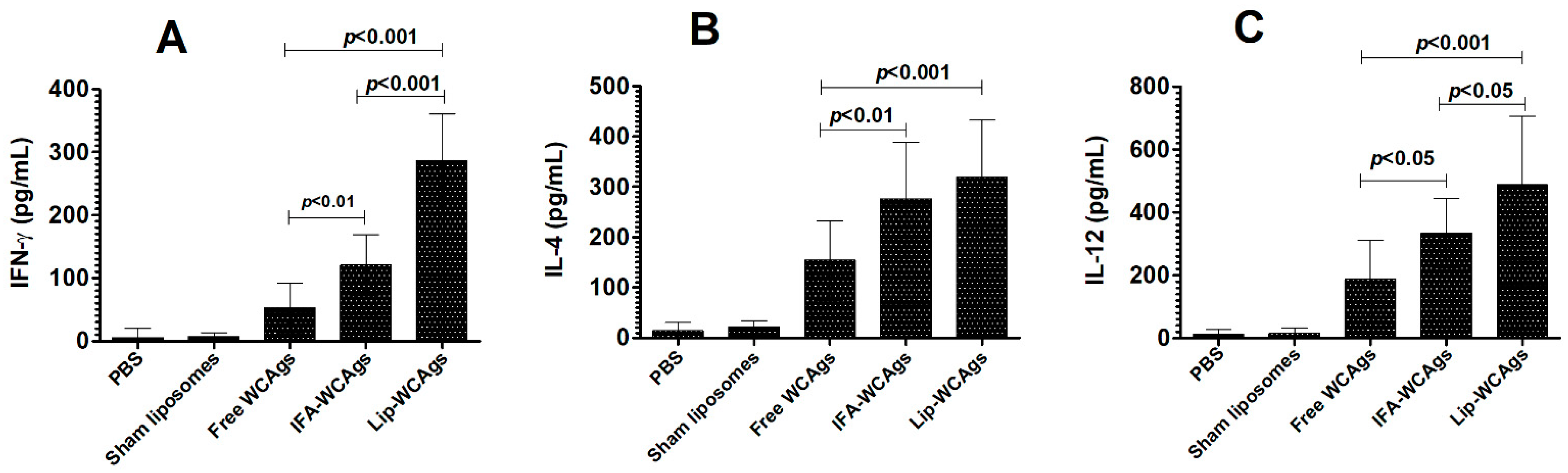
3.5. The Splenocytes from the Mice Immunized with Lip-WCAgs Secreted Higher Levels of Cytokines
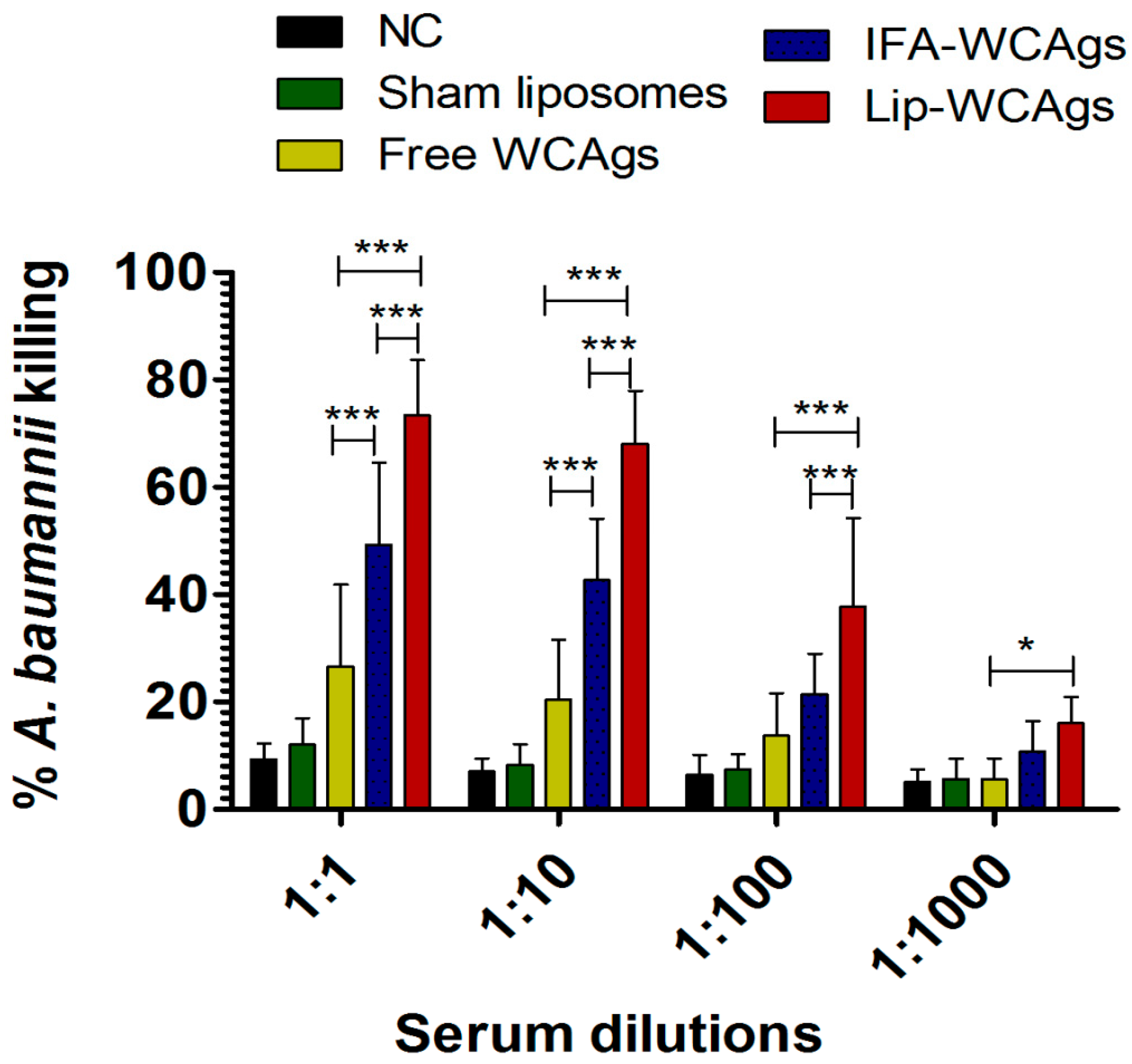
3.6. Antisera from Lip-WCAgs-Immunized Mice Showed Greater Opsonophagocytic Killing of A. baumannii
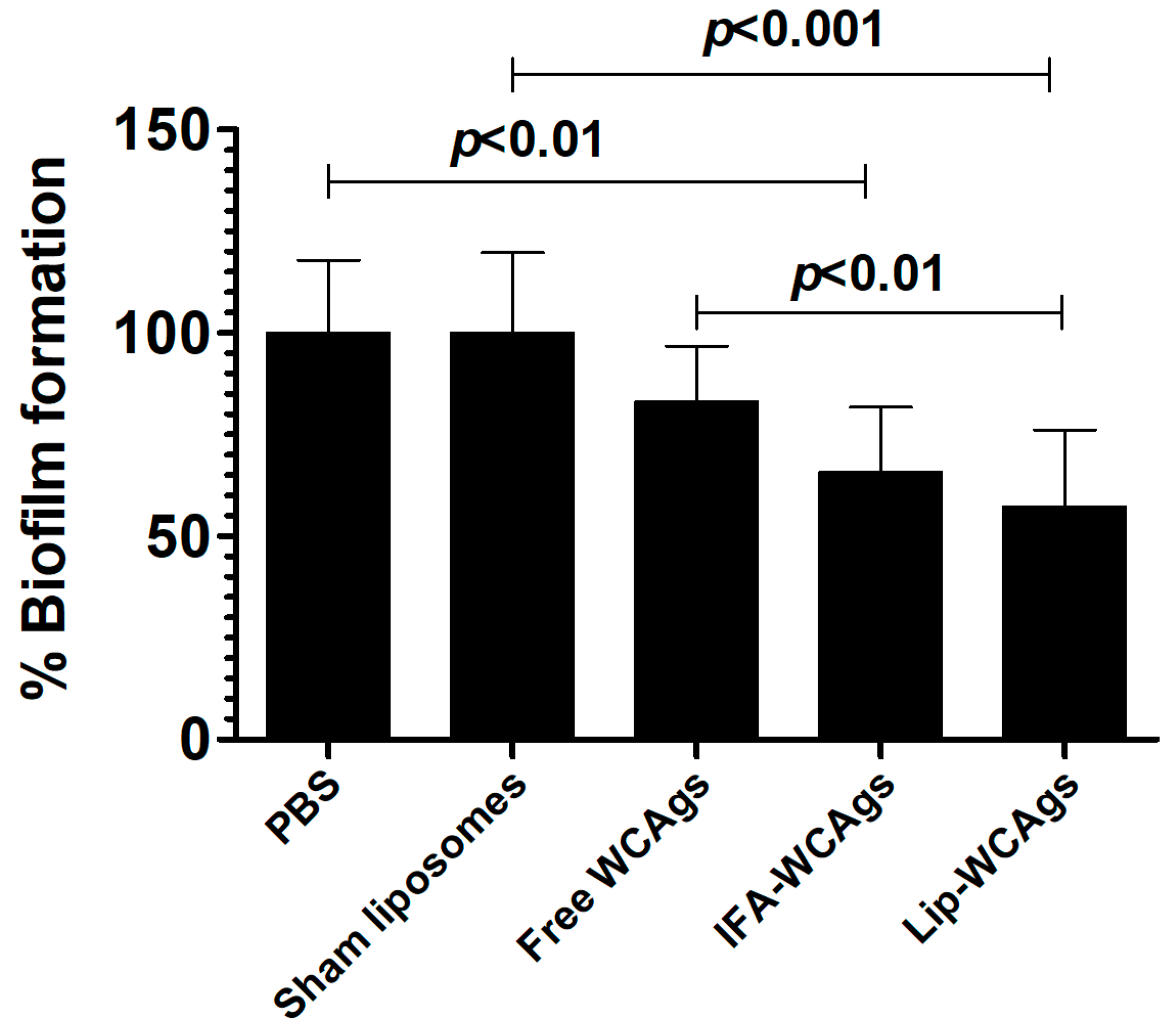
3.7. A. baumannii-Specific Antisera Inhibited the Biofilm Formation
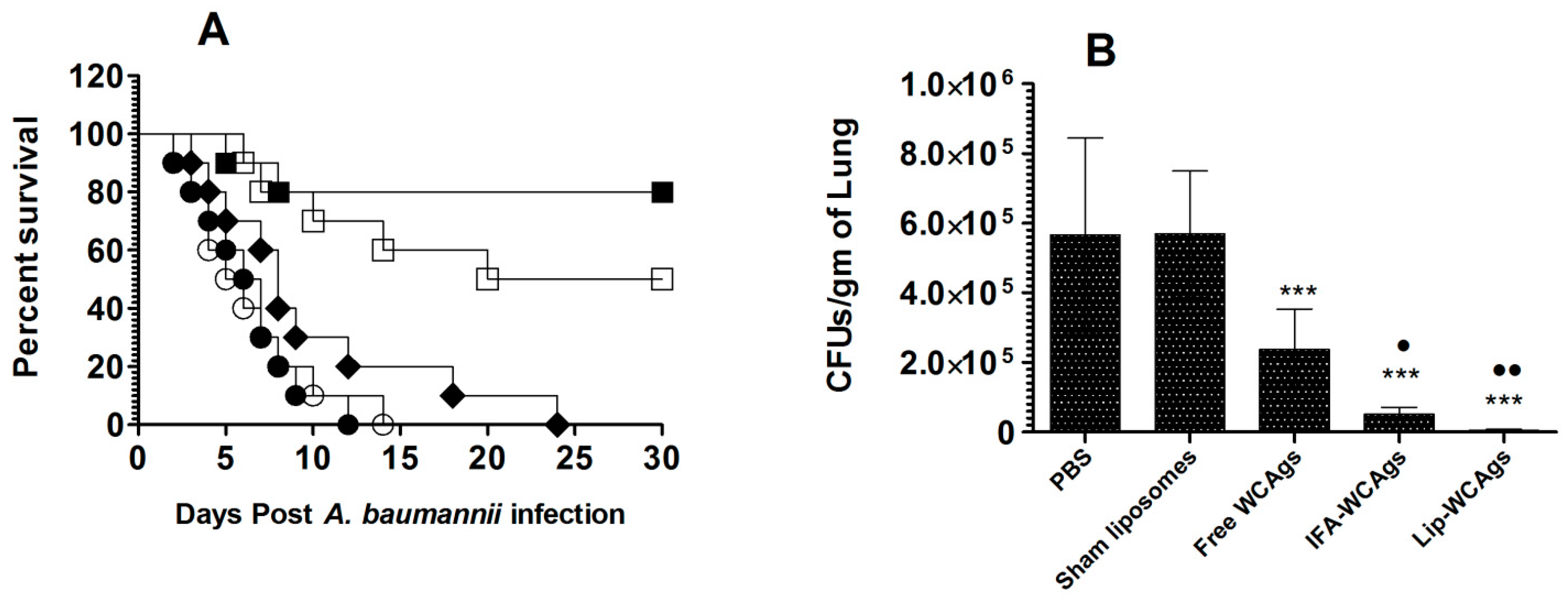
3.8. Immunization with Lip-WCAgs Effectively Protected the Mice against the Systemic Infection of A. baumannii
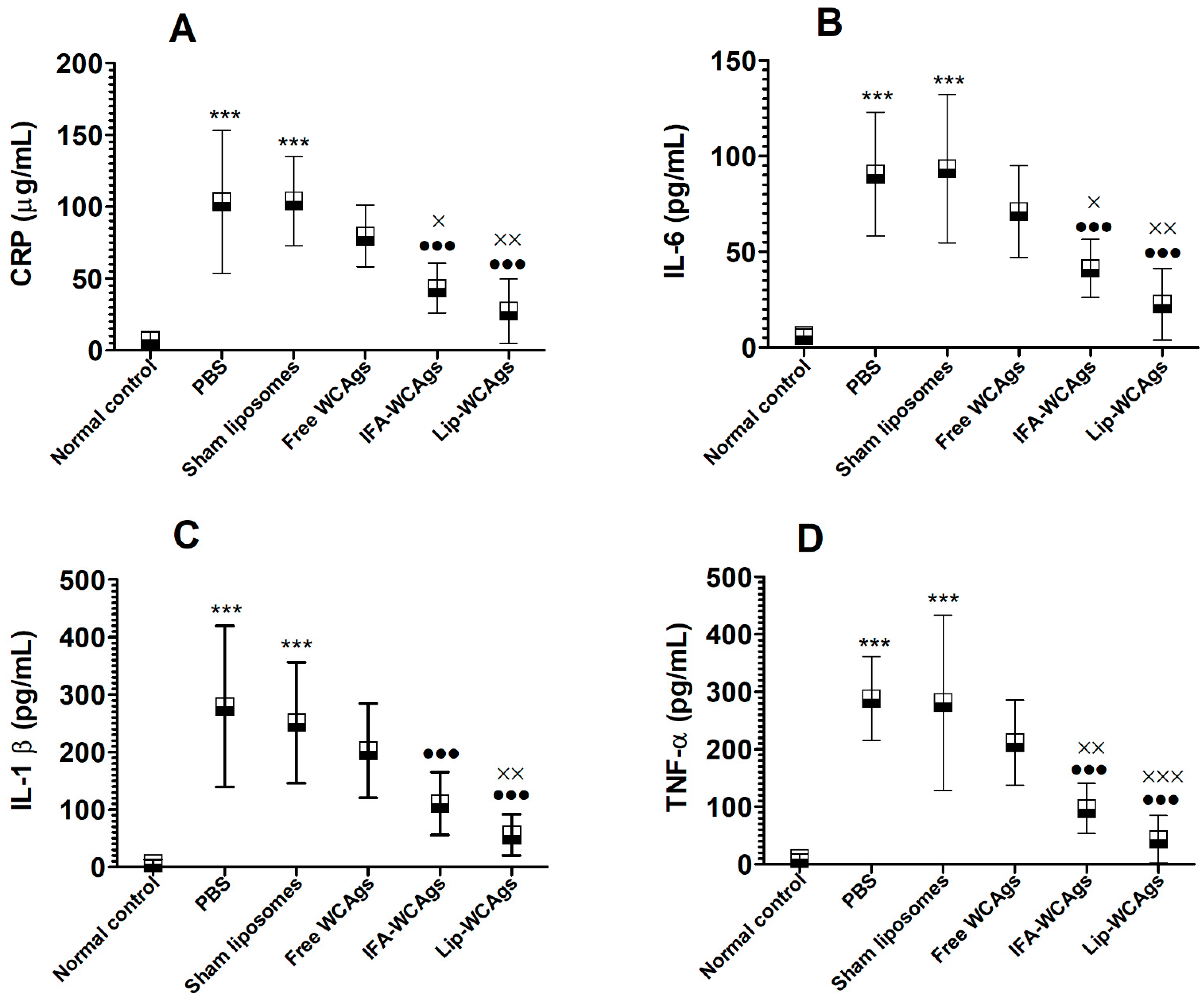
3.9. Lip-WCAgs-Immunized Mice Showed Lower Levels of Inflammation Markers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roca, I.; Espinal, P.; Vila-Farrés, X.; Vila, J. The Acinetobacter baumannii Oxymoron: Commensal Hospital Dweller Turned Pan-Drug-Resistant Menace. Front. Microbiol. 2012, 3, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarshar, M.; Behzadi, P.; Scribano, D.; Palamara, A.T.; Ambrosi, C. Acinetobacter baumannii: An Ancient Commensal with Weapons of a Pathogen. Pathogens 2021, 10, 387. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.E. Prevalence of Acinetobacter baumannii in Saudi Arabia: Risk factors, antimicrobial resistance patterns and mechanisms of carbapenem resistance. Ann. Clin. Microbiol. Antimicrob. 2019, 18, 1. [Google Scholar] [CrossRef] [PubMed]
- Al-Hamad, A.; Pal, T.; Leskafi, H.; Abbas, H.; Hejles, H.; Alsubikhy, F.; Darwish, D.; Ghazawi, A.; Sonnevend, A. Molecular characterization of clinical and environmental carbapenem resistant Acinetobacter baumannii isolates in a hospital of the Eastern Region of Saudi Arabia. J. Infect. Public Health 2020, 13, 632–636. [Google Scholar] [CrossRef]
- Lin, M.F.; Lan, C.Y. Antimicrobial resistance in Acinetobacter baumannii: From bench to bedside. World J. Clin. Cases 2014, 2, 787–814. [Google Scholar] [CrossRef]
- Aly, M.; Tayeb, H.T.; Al Johani, S.M.; Alyamani, E.J.; Aldughaishem, F.; Alabdulkarim, I.; Balkhy, H.H. Genetic diversity of OXA-51-like genes among multidrug-resistant Acinetobacter baumannii in Riyadh, Saudi Arabia. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1223–1228. [Google Scholar] [CrossRef]
- Saeed, N.K.; Kambal, A.M.; El-Khizzi, N.A. Antimicrobial-resistant bacteria in a general intensive care unit in Saudi Arabia. Saudi Med. J. 2010, 31, 1341–1349. [Google Scholar]
- Gedefie, A.; Demsis, W.; Ashagrie, M.; Kassa, Y.; Tesfaye, M.; Tilahun, M.; Bisetegn, H.; Sahle, Z. Acinetobacter baumannii Biofilm Formation and Its Role in Disease Pathogenesis: A Review. Infect. Drug Resist. 2021, 14, 3711–3719. [Google Scholar] [CrossRef]
- Nasr, P. Genetics, epidemiology, and clinical manifestations of multidrug-resistant Acinetobacter baumannii. J. Hosp. Infect. 2020, 104, 4–11. [Google Scholar] [CrossRef]
- Dijkshoorn, L.; Nemec, A.; Seifert, H. An increasing threat in hospitals: Multidrug-resistant Acinetobacter baumannii. Nat. Rev. Microbiol. 2007, 5, 939–951. [Google Scholar] [CrossRef]
- Gellings, P.S.; Wilkins, A.A.; Morici, L.A. Recent Advances in the Pursuit of an Effective Acinetobacter baumannii Vaccine. Pathogens 2020, 9, 1066. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, T.; Cao, J.; Sun, J.; Dai, W.; Zhang, L. Mucosal immunization with purified OmpA elicited protective immunity against infections caused by multidrug-resistant Acinetobacter baumannii. Microb. Pathog. 2016, 96, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Yao, Y.; Wang, S.; Xia, Y.; Yang, X.; Long, Q.; Sun, W.; Liu, C.; Li, Y.; Chu, X.; et al. Immunization with a 22-kDa outer membrane protein elicits protective immunity to multidrug-resistant Acinetobacter baumannii. Sci. Rep. 2016, 6, 20724. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Xue, J.; Jiang, M.; Lin, S.; Huang, Y.; Deng, K.; Shu, L.; Xu, H.; Li, Z.; Yao, J.; et al. A Multiepitope Peptide, rOmp22, Encapsulated in Chitosan-PLGA Nanoparticles as a Candidate Vaccine Against Acinetobacter baumannii Infection. Int. J. Nanomed. 2021, 16, 1819–1836. [Google Scholar] [CrossRef]
- Khan, A.A.; Allemailem, K.S.; Almatroodi, S.A.; Almatroudi, A.; Rahmani, A.H. Recent strategies towards the surface modification of liposomes: An innovative approach for different clinical applications. 3 Biotech. 2020, 10, 163. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A. Targeted Drug Delivery Using Tuftsin-bearing Liposomes: Implications in the Treatment of Infectious Diseases and Tumors. Curr. Drug Targets 2021, 22, 770–778. [Google Scholar] [CrossRef]
- Alameh, M.G.; Tombácz, I.; Bettini, E.; Lederer, K.; Sittplangkoon, C.; Wilmore, J.R.; Gaudette, B.T.; Soliman, O.Y.; Pine, M.; Hicks, P.; et al. Lipid nanoparticles enhance the efficacy of mRNA and protein subunit vaccines by inducing robust T follicular helper cell and humoral responses. Immunity 2021, 54, 2877–2892.e7. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, C.; Yang, S.; Xiao, W.; Zheng, Q.; Song, X. The investigation of mRNA vaccines formulated in liposomes administrated in multiple routes against SARS-CoV-2. J. Control. Release 2021, 335, 449–456. [Google Scholar] [CrossRef]
- Tian, M.; Zhou, Z.; Tan, S.; Fan, X.; Li, L.; Ullah, N. Formulation in DDA-MPLA-TDB Liposome Enhances the Immunogenicity and Protective Efficacy of a DNA Vaccine against Mycobacterium tuberculosis Infection. Front. Immunol. 2018, 9, 310. [Google Scholar] [CrossRef]
- Bhalla, M.; Nayerhoda, R.; Tchalla, E.Y.I.; Abamonte, A.; Park, D.; Simmons, S.R.; Pfeifer, B.A.; Bou Ghanem, E.N. Liposomal Encapsulation of Polysaccharides (LEPS) as an Effective Vaccine Strategy to Protect Aged Hosts against S. pneumoniae Infection. Front. Aging 2021, 2, 798868. [Google Scholar] [CrossRef]
- Ahmad, N.; Deeba, F.; Faisal, S.M.; Khan, A.; Agrewala, J.N.; Dwivedi, V.; Owais, M. Role of fusogenic non-PC liposomes in elicitation of protective immune response against experimental murine salmonellosis. Biochimie 2006, 88, 1391–1400. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Khan, A. Role of NKT Cells during Viral Infection and the Development of NKT Cell-Based Nanovaccines. Vaccines 2021, 9, 949. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Malik, A.; Alruwetei, A.M.; Alzohairy, M.A.; Alhatlani, B.Y.; Al Rugaie, O.; Alhumaydhi, F.A.; Khan, A. Delivery of MERS antigen encapsulated in α-GalCer-bearing liposomes elicits stronger antigen-specific immune responses. J. Drug Target 2022, 22, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.H.; Younus, H.; Allemailem, K.S.; Almatroudi, A.; Alrumaihi, F.; Alruwetei, A.M.; Alsahli, M.A.; Khan, A.; Khan, M.A. Potential of Methylglyoxal-Conjugated Chitosan Nanoparticles in Treatment of Fluconazole-Resistant Candida albicans Infection in a Murine Model. Int. J. Nanomed. 2020, 15, 3681–3693. [Google Scholar] [CrossRef]
- Khan, M.A.; Khan, A.; Khan, S.H.; Azam, M.; Khan, M.M.U.; Khalilullah, H.; Younus, H. Coadministration of liposomal methylglyoxal increases the activity of amphotericin B against Candida albicans in leukopoenic mice. J. Drug Target 2021, 29, 78–87. [Google Scholar] [CrossRef]
- Lyu, Z.; Shang, Y.; Wang, X.; Wu, Y.; Zheng, J.; Liu, H.; Gong, T.; Ye, L.; Qu, D. Monoclonal Antibodies Specific to the Extracellular Domain of Histidine Kinase YycG of Staphylococcus epidermidis Inhibit Biofilm Formation. Front. Microbiol. 2020, 11, 18390. [Google Scholar] [CrossRef] [PubMed]
- Allemailem, K.S.; Alnuqaydan, A.M.; Almatroudi, A.; Alrumaihi, F.; Aljaghwani, A.; Khalilullah, H.; Younus, H.; Khan, A.; Khan, M.A. Safety and Therapeutic Efficacy of Thymoquinone-Loaded Liposomes against Drug-Sensitive and Drug-Resistant Acinetobacter baumannii. Pharmaceutics 2021, 13, 677. [Google Scholar] [CrossRef] [PubMed]
- Katip, W.; Oberdorfer, P. Clinical Efficacy and Nephrotoxicity of Colistin Alone versus Colistin Plus Vancomycin in Critically Ill Patients Infected with Carbapenem-Resistant Acinetobacter baumannii: A Propensity Score-Matched Analysis. Pharmaceutics 2021, 13, 162. [Google Scholar] [CrossRef] [PubMed]
- Katip, W.; Uitrakul, S.; Oberdorfer, P.A. Comparison of Colistin versus Colistin Plus Meropenem for the Treatment of Carbapenem-Resistant Acinetobacter baumannii in Critically Ill Patients: A Propensity Score-Matched Analysis. Antibiotiotics 2020, 9, 647. [Google Scholar] [CrossRef]
- Syed, F.M.; Khan, M.A.; Nasti, T.H.; Ahmad, N.; Mohammad, O. Antigen entrapped in the escheriosomes leads to the generation of CD4(+) helper and CD8(+) cytotoxic T cell response. Vaccine 2003, 21, 2383–2393. [Google Scholar] [CrossRef]
- Schwendener, R.A. Liposomes as vaccine delivery systems: A review of the recent advances. Adv. Vaccines 2014, 2, 159–182. [Google Scholar] [CrossRef] [PubMed]
- McConnell, M.J.; Domínguez-Herrera, J.; Smani, Y.; López-Rojas, R.; Docobo-Pérez, F.; Pachón, J. Vaccination with outer membrane complexes elicits rapid protective immunity to multidrug-resistant Acinetobacter baumannii. Infect. Immun. 2011, 79, 518–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McConnell, M.J.; Pachón, J. Active and passive immunization against Acinetobacter baumannii using an inactivated whole cell vaccine. Vaccine 2010, 29, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bentancor, L.V.; O’Malley, J.M.; Bozkurt-Guzel, C.; Pier, G.B.; Maira-Litrán, T. Poly-N-acetyl-β-(1-6)-glucosamine is a target for protective immunity against Acinetobacter baumannii infections. Infect. Immun. 2012, 80, 651–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McConnell, M.J.; Rumbo, C.; Bou, G.; Pachón, J. Outer membrane vesicles as an acellular vaccine against Acinetobacter baumannii. Vaccine 2011, 29, 5705–5710. [Google Scholar] [CrossRef] [PubMed]
- Bentancor, L.V.; Routray, A.; Bozkurt-Guzel, C.; Camacho-Peiro, A.; Pier, G.B.; Maira-Litrán, T. Evaluation of the trimeric autotransporter Ata as a vaccine candidate against Acinetobacter baumannii infections. Infect. Immun. 2012, 80, 3381–3388. [Google Scholar] [CrossRef] [Green Version]
- Luo, G.; Lin, L.; Ibrahim, A.S.; Baquir, B.; Pantapalangkoor, P.; Bonomo, R.A.; Doi, Y.; Adams, M.D.; Russo, T.A.; Spellberg, B. Active and passive immunization protects against lethal, extreme drug resistant-Acinetobacter baumannii infection. PLoS ONE 2012, 7, e29446. [Google Scholar] [CrossRef] [Green Version]
- Russo, T.A.; Beanan, J.M.; Olson, R.; MacDonald, U.; Cox, A.D.; St Michael, F.; Vinogradov, E.V.; Spellberg, B.; Luke-Marshall, N.R.; Campagnari, A.A. The K1 capsular polysaccharide from Acinetobacter baumannii is a potential therapeutic target via passive immunization. Infect. Immun. 2013, 81, 915–922. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Tan, B.; Pantapalangkoor, P.; Ho, T.; Hujer, A.M.; Taracila, M.A.; Bonomo, R.A.; Spellberg, B. Acinetobacter baumannii rOmpA vaccine dose alters immune polarization and immunodominant epitopes. Vaccine 2013, 31, 313–318. [Google Scholar] [CrossRef] [Green Version]
- Goel, V.K.; Kapil, A. Monoclonal antibodies against the iron regulated outer membrane Proteins of Acinetobacter baumannii are bactericidal. BMC Microbiol. 2001, 1, 16. [Google Scholar] [CrossRef]
- Getahun, A.; Dahlström, J.; Wernersson, S.; Heyman, B. IgG2a-mediated enhancement of antibody and T cell responses and its relation to inhibitory and activating Fc gamma receptors. J. Immunol. 2004, 172, 5269–5276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Freitas, S.B.; Wozeak, D.R.; Neto, A.S.; Cardoso, T.L.; Hartwig, D.D. A hypothetical adhesin protein induces anti-biofilm antibodies against multi-drug resistant Acinetobacter baumannii. Microb. Pathog. 2021, 159, 105112. [Google Scholar] [PubMed]
- Naghipour Erami, A.; Rasooli, I.; Jahangiri, A.; Darvish Alipour Astaneh, S. Anti-Omp34 antibodies protect against Acinetobacter baumannii in a murine sepsis model. Microb. Pathog. 2021, 161, 105291. [Google Scholar] [CrossRef]
- Singh, R.; Capalash, N.; Sharma, P. Immunoprotective potential of BamA, the outer membrane protein assembly factor, against MDR Acinetobacter baumannii. Sci. Rep. 2017, 7, 12411. [Google Scholar] [CrossRef]
- Huang, W.; Zhang, Q.; Li, W.; Chen, Y.; Shu, C.; Li, Q.; Zhou, J.; Ye, C.; Bai, H.; Sun, W.; et al. Anti-outer Membrane Vesicle Antibodies Increase Antibiotic Sensitivity of Pan-Drug-Resistant Acinetobacter baumannii. Front. Microbiol. 2019, 10, 1379. [Google Scholar] [CrossRef]
- Smani, Y.; Docobo-Pérez, F.; McConnell, M.J.; Pachón, J. Acinetobacter baumannii-induced lung cell death: Role of inflammation, oxidative stress and cytosolic calcium. Microb. Pathog. 2011, 50, 224–232. [Google Scholar] [CrossRef]
- Huang, W.; Yao, Y.; Long, Q.; Yang, X.; Sun, W.; Liu, C.; Jin, X.; Li, Y.; Chu, X.; Chen, B.; et al. Immunization against multidrug-resistant Acinetobacter baumannii effectively protects mice in both pneumonia and sepsis models. PLoS ONE 2014, 9, e100727. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.A.; Allemailem, K.S.; Maswadeh, H.; Younus, H. Safety and Prophylactic Efficacy of Liposome-Based Vaccine against the Drug-Resistant Acinetobacter baumannii in Mice. Pharmaceutics 2022, 14, 1357. https://doi.org/10.3390/pharmaceutics14071357
Khan MA, Allemailem KS, Maswadeh H, Younus H. Safety and Prophylactic Efficacy of Liposome-Based Vaccine against the Drug-Resistant Acinetobacter baumannii in Mice. Pharmaceutics. 2022; 14(7):1357. https://doi.org/10.3390/pharmaceutics14071357
Chicago/Turabian StyleKhan, Masood Alam, Khaled S. Allemailem, Hamzah Maswadeh, and Hina Younus. 2022. "Safety and Prophylactic Efficacy of Liposome-Based Vaccine against the Drug-Resistant Acinetobacter baumannii in Mice" Pharmaceutics 14, no. 7: 1357. https://doi.org/10.3390/pharmaceutics14071357
APA StyleKhan, M. A., Allemailem, K. S., Maswadeh, H., & Younus, H. (2022). Safety and Prophylactic Efficacy of Liposome-Based Vaccine against the Drug-Resistant Acinetobacter baumannii in Mice. Pharmaceutics, 14(7), 1357. https://doi.org/10.3390/pharmaceutics14071357