
Isosteviol Sodium (STVNA) Reduces Pro-Inflammatory Cytokine IL-6 and GM-CSF in an In Vitro Murine Stroke Model of the Blood–Brain Barrier (BBB)

 , , , ,
, , , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture
2.3. Oxygen-Glucose Deprivation (OGD) and Isosteviol Sodium (STVNA) Treatment
2.4. Western Blot
2.5. Nucleus/Cytoplasm Fractionation
2.6. Immunofluorescence Staining
2.7. Multiplex ELISA
2.8. Transfection and Luciferase Assay
2.9. Statistical Analysis
3. Results
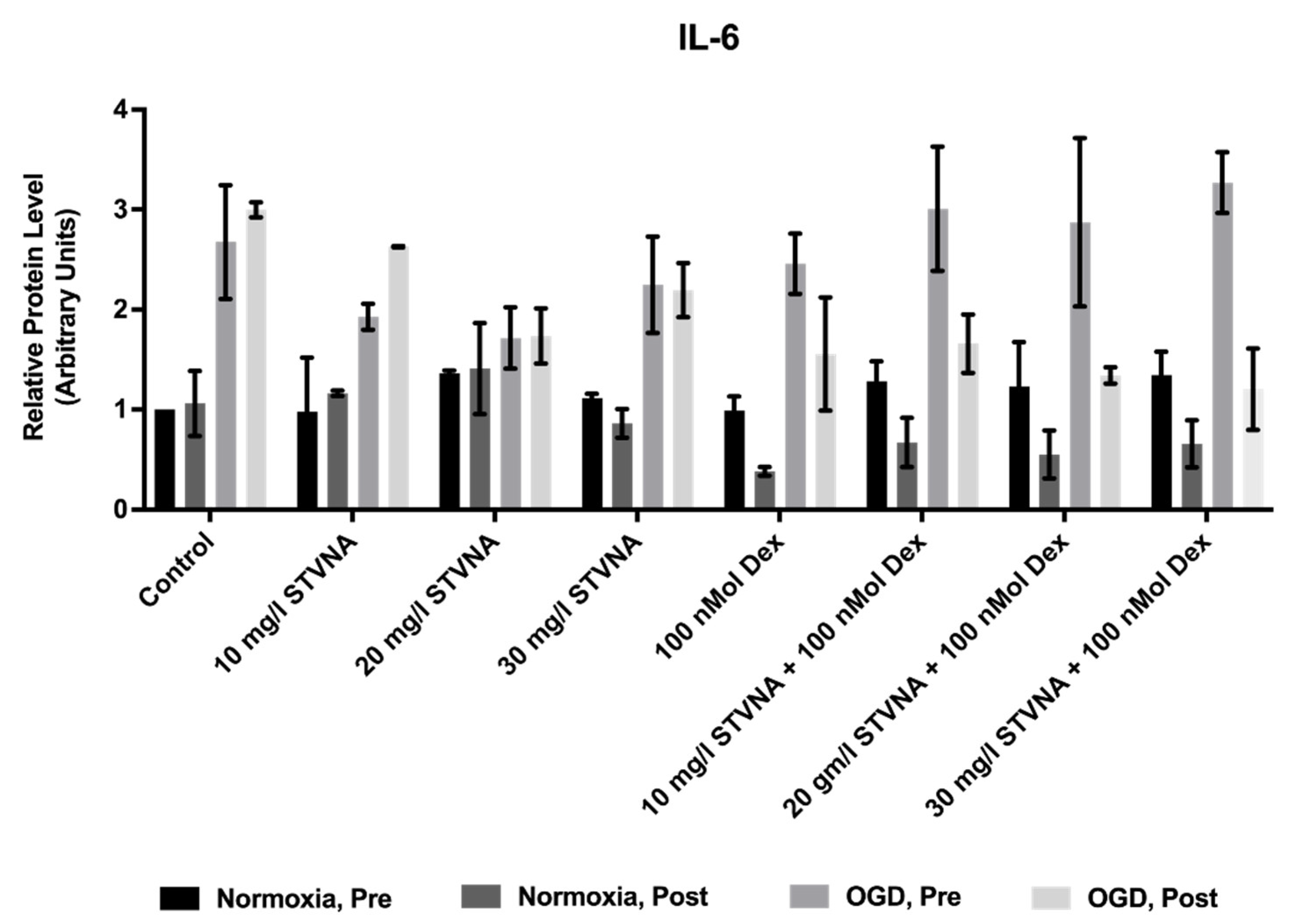
3.1. STVNA Administration Demonstrates a Trend towards Reduced OGD-Induced IL-6 Expression
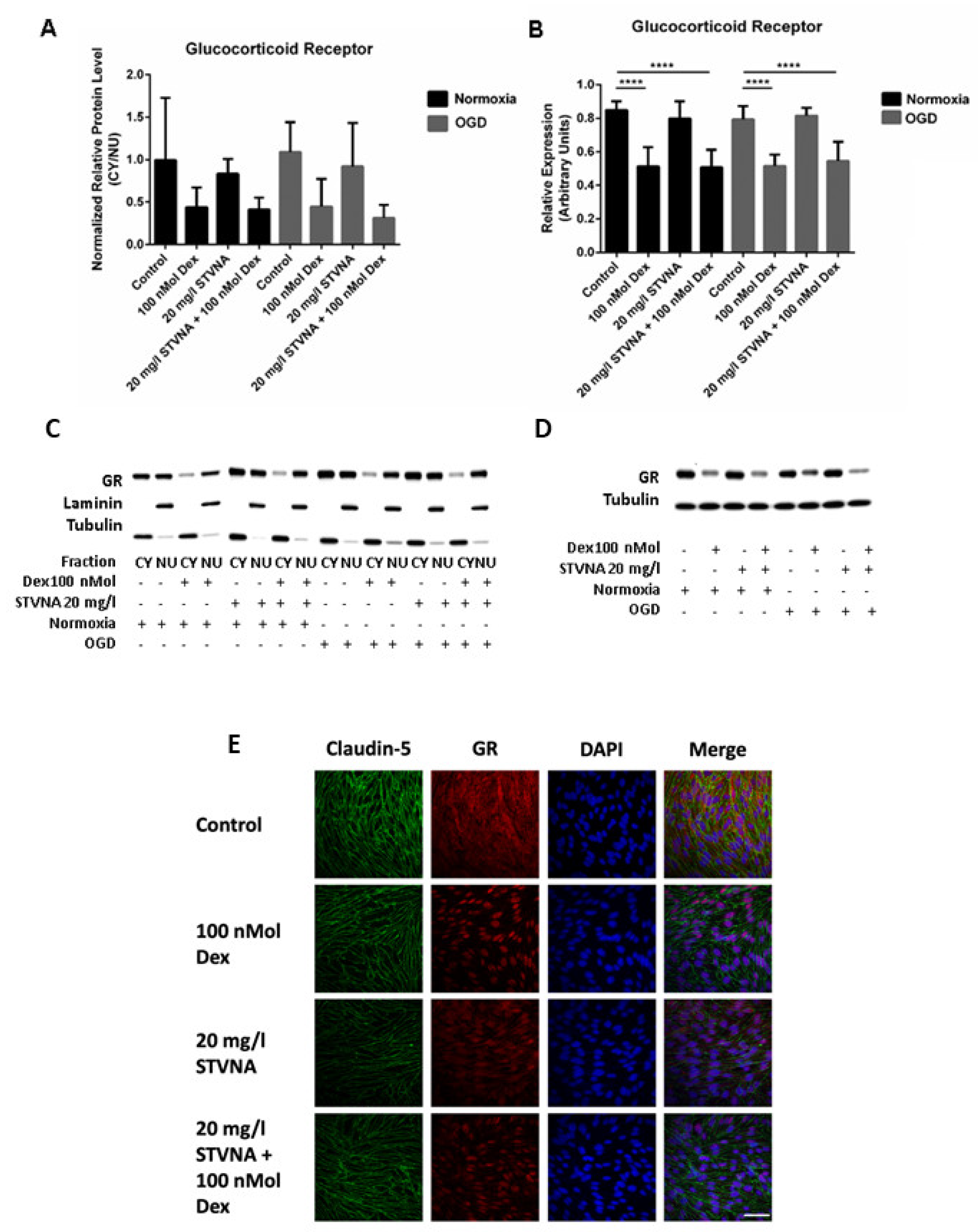
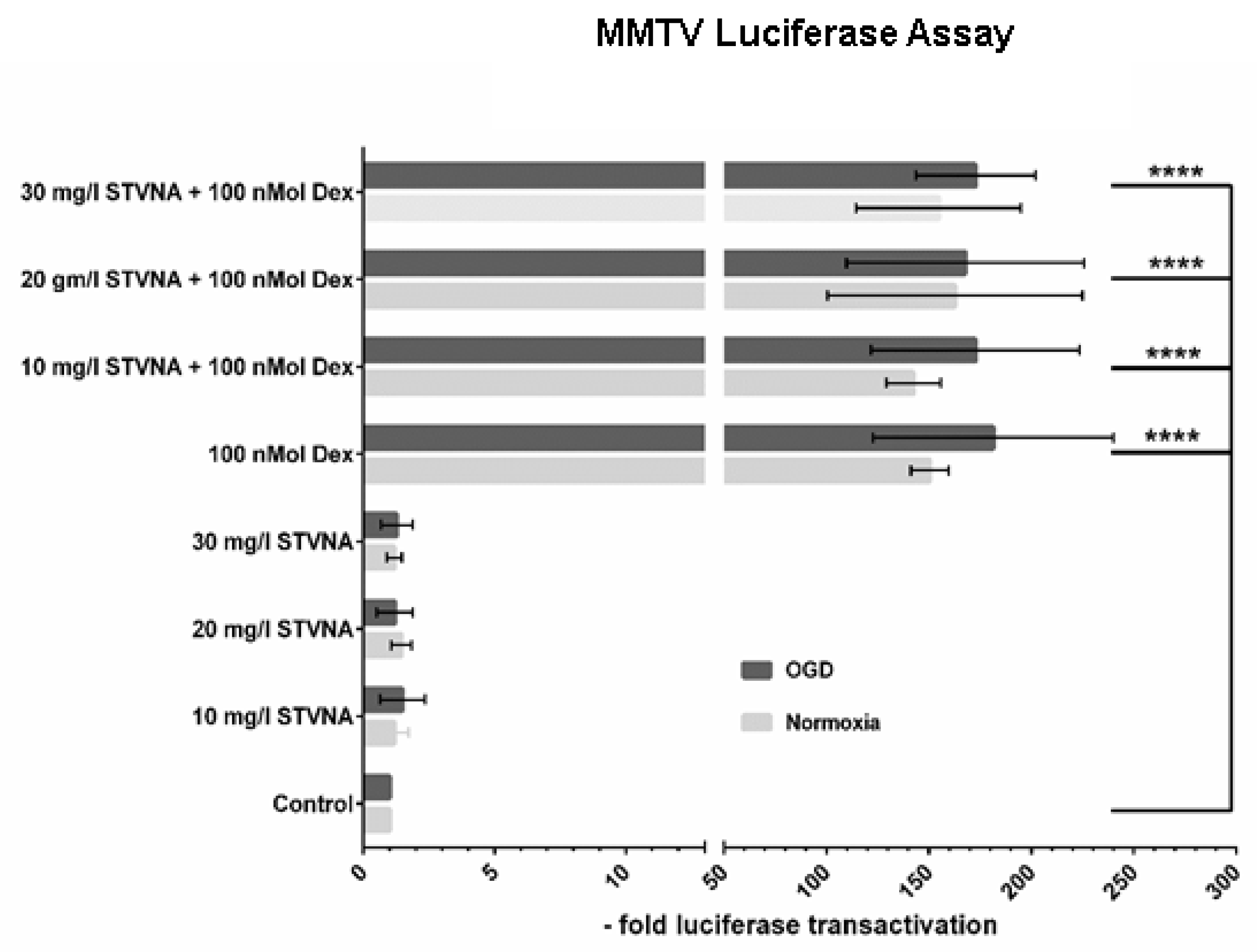
3.2. Glucocorticoid Receptor (GR) Activity Is Unaltered by STVNA
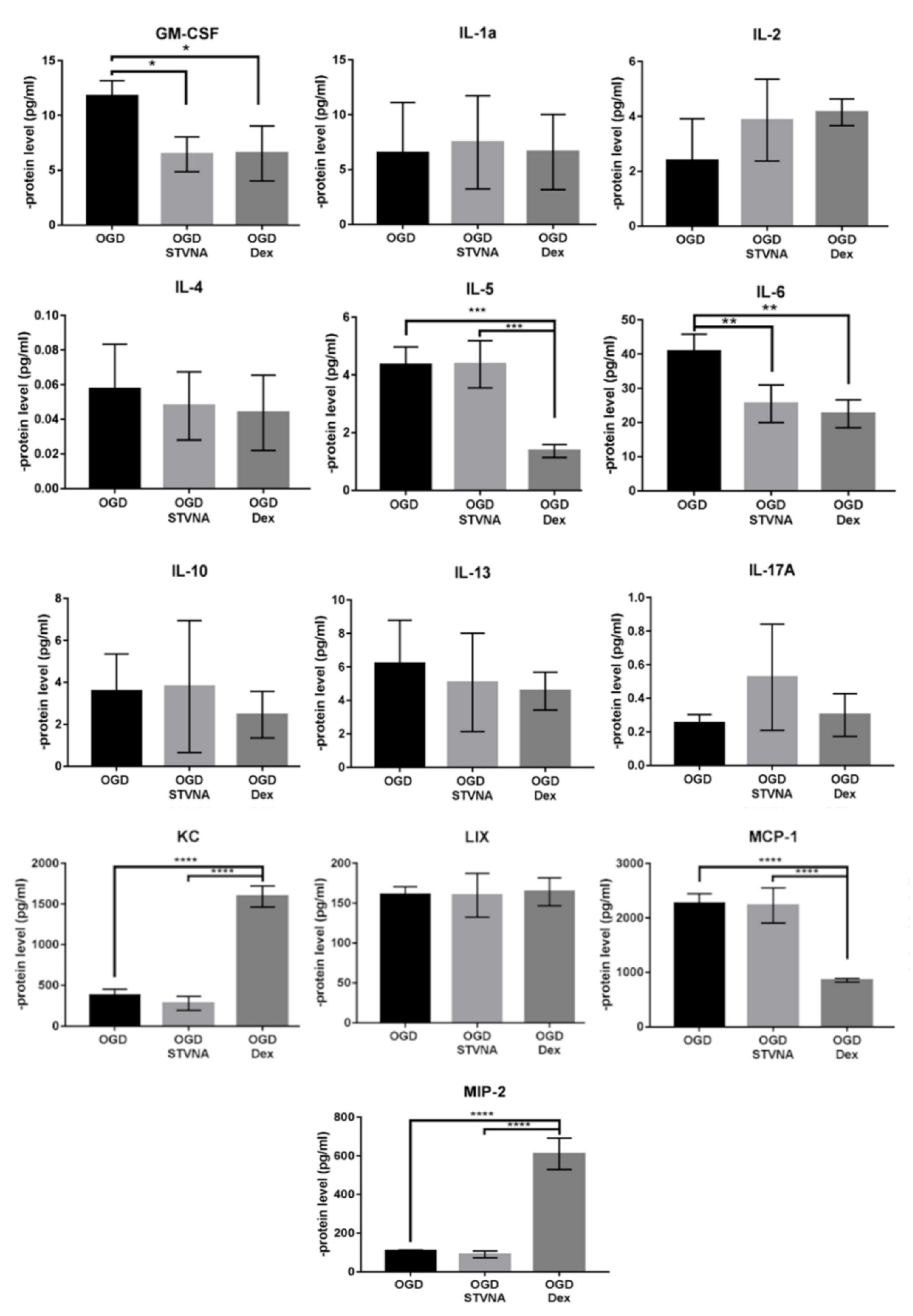
3.3. Decreased IL-6 and GSM-CSF Expression after STVNA Treatment
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feigin, V.L.; Krishnamurthi, R.V.; Parmar, P.; Norrving, B.; Mensah, G.A.; Bennett, D.A.; Barker-Collo, S.; Moran, A.E.; Sacco, R.L.; Truelsen, T.; et al. Update on the global burden of ischemic and hemorrhagic stroke in 1990–2013: The GBD 2013 study. Neuroepidemiology 2015, 45, 161–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacco, R.L.; Kasner, S.E.; Broderick, J.P.; Caplan, L.R.; Connors, J.J.; Culebras, A.; Elkind, M.S.V.; George, M.G.; Hamdan, A.D.; Higashida, R.T.; et al. An updated definition of stroke for the 21st century: A statement for healthcare professionals from the American Heart Association/American Stroke Association. Stroke 2013, 44, 2064–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maida, C.D.; Norrito, R.L.; Daidone, M.; Tuttolomondo, A.; Pinto, A. Neuroinflammatory Mechanisms in Ischemic Stroke: Focus on Cardioembolic Stroke, Background, and Therapeutic Approaches. Int. J. Mol. Sci. 2020, 21, 6454. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Andjelkovic, A.V.; Zhu, L.; Yang, T.; Bennett, M.V.L.; Chen, J.; Keep, R.F.; Shi, Y. Blood-brain barrier dysfunction and recovery after ischemic stroke. Prog. Neurobiol. 2018, 163–164, 144–171. [Google Scholar]
- Abdullahi, W.; Tripathi, D.; Ronaldson, P.T. Blood-brain barrier dysfunction in ischemic stroke: Targeting tight junctions and transporters for vascular protection. Am. J. Physiol. Cell Physiol. 2018, 315, C343–C356. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Hawkins, K.E.; Doré, S.; Candelario-Jalil, E. Neuroinflammatory mechanisms of blood-brain barrier damage in ischemic stroke. Am. J. Physiol. Cell Physiol. 2019, 316, C135–C153. [Google Scholar] [CrossRef] [PubMed]
- Khatri, R.; McKinney, A.M.; Swenson, B.; Janardhan, V. Blood-brain barrier, reperfusion injury, and hemorrhagic transfor-mation in acute ischemic stroke. Neurology 2012, 79 (Suppl. S1), S52–S57. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, G.A.; Yang, Y. Vasogenic edema due to tight junction disruption by matrix metalloproteinases in cerebral is-chemia. Neurosurg. Focus 2007, 22, E4. [Google Scholar]
- Brouns, R.; Wauters, A.; De Surgeloose, D.; Mariën, P.; De Deyn, P.P. Biochemical markers for blood-brain barrier dysfunc-tion in acute ischemic stroke correlate with evolution and outcome. Eur. Neurol. 2011, 65, 23–31. [Google Scholar] [CrossRef]
- Salvador, E.; Shityakov, S.; Förster, C. Glucocorticoids and endothelial cell barrier function. Cell Tissue Res. 2014, 355, 597–605. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.H.; He, F.; Zhang, N.N.; Zhao, Z.A.; Chen, H.S. Time dependent neuroprotection of dexamethasone in experimental focal cerebral ischemia: The involvement of NF-κB pathways. Brain Res. 2018, 1701, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-T.; Lee, T.-H.; Lee, I.-N.; Chung, C.-Y.; Kuo, C.-H.; Weng, H.-H. Dexamethasone inhibits ICAM-1 and MMP-9 ex-pression and reduces brain edema inintracerebral hemorrhagic rats. Acta. Neurochir. 2011, 153, 2197–2203. [Google Scholar] [CrossRef] [PubMed]
- Sandercock, P.A.; Soane, T. Corticosteroids for acute ischaemic stroke. Cochrane Database Syst. Rev. 2011, 2011, CD000064. [Google Scholar] [CrossRef]
- Alderson, P.; Roberts, I. Corticosteroids for acute traumatic brain injury. Cochrane Database Syst. Rev. 2005, 2005, CD000196. [Google Scholar] [CrossRef] [PubMed]
- Roberts, I.; Yates, D.; Sandercock, P.; Farrell, B.; Wasserberg, J.; Lomas, G.; Cottingham, R.; Svoboda, P.; Brayley, N.; Mazairac, G.; et al. Effect of intravenous corticosteroids on death within 14 days in 10008 adults with clinically significant head injury (MRC CRASH trial): Randomised placebocontrolled trial. Lancet 2004, 364, 1321–1328. [Google Scholar]
- Thal, S.C.; Schaible, E.V.; Neuhaus, W.; Scheffer, D.; Brandstetter, M.; Engelhard, K.; Wunder, C.; Förster, C.Y. Inhibition of proteasomal glucocorticoid receptor degradation restores dexamethasone-mediated stabilization of the blood-brain barrier after traumatic brain injury. Crit. Care Med. 2013, 41, 1305–1315. [Google Scholar] [CrossRef]
- Kleinschnitz, C.; Blecharz, K.; Kahles, T.; Schwarz, T.; Kraft, P.; Gobel, K.; Meuth, S.G.; Burek, M.; Thum, T.; Stoll, G.; et al. Glucocorticoid insensitivity at the hypoxic blood-brain barrier can be reversed by inhibition of the proteasome. Stroke 2011, 42, 1081–1089. [Google Scholar] [CrossRef] [Green Version]
- Rösing, N.; Salvador, E.; Güntzel, P.; Kempe, C.; Burek, M.; Holzgrabe, U.; Soukhoroukov, V.; Wunder, C.; Förster, C. Neu-roprotective Effects of Isosteviol Sodium in Murine Brain Capillary Cerebellar Endothelial Cells (cerebEND) After Hypox-ia. Front Cell Neurosci. 2020, 14, 573950. [Google Scholar] [CrossRef]
- Mosettig, E.; Beglinger, U.; Dolder, F.; Lichti, H.; Quitt, P.; Walter, J. The Absolute Configuration of Steviol and Isosteviol. J. Am. Chem. Soc. 1963, 85, 2305–2309. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, X.; Xie, Y.; Zan, J.; Tan, W. Isosteviol Sodium Protects Against Permanent Cerebral Ischemia Injury in Mice via Inhibition of NF-κB-Mediated Inflammatory and Apoptotic Responses. J. Stroke Cerebrovasc. Dis. 2017, 26, 2603–2614. [Google Scholar] [CrossRef]
- Zan, J.; Zhang, H.; Lu, M.Y.; Beng, H.M.; Zhong, K.L.; Sun, X.O.; Tan, W. Isosteviol sodium injection improves outcomes by modulating TLRs/NF-κB-dependent inflammatory responses following experimental traumatic brain injury in rats. Neu-Roreport 2018, 29, 794–803. [Google Scholar] [CrossRef] [PubMed]
- Panagiotou, C.; Mihailidou, C.; Brauhli, G.; Katsarou, O.; Moutsatsou, P. Effect of steviol, steviol glycosides and stevia ex-tract on glucocorticoid receptor signaling in normal and cancer blood cells. Mol. Cell. Endocrinol. 2018, 460, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Silwedel, C.; Förster, C. Differential susceptibility of cerebral and cerebellar murine brain microvascular endothelial cells to loss of barrier properties in response to inflammatory stimuli. J. Neuroimmunol. 2006, 179, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Dilling, C.; Roewer, N.; Förster, C.Y.; Burek, M. Multiple protocadherins are expressed in brain microvascular endothelial cells and might play a role in tight junction protein regulation. J. Cereb. Blood Flow Metab. 2017, 37, 3391–3400. [Google Scholar] [CrossRef] [Green Version]
- Helms, H.C.; Abbott, N.J.; Burek, M.; Cecchelli, R.; Couraud, P.O.; Deli, M.A.; Förster, C.; Galla, H.J.; Romero, I.A.; Shusta, E.V.; et al. In vitro models of the blood-brain barrier: An overview of commonly used brain endothelial cell culture models and guidelines for their use. J. Cereb. Blood Flow Metab. 2016, 36, 862–890. [Google Scholar] [CrossRef]
- Burek, M.; Burmester, S.; Salvador, E.; Möller-Ehrlich, K.; Schneider, R.; Roewer, N.; Nagai, M.; Förster, C.Y. Kidney Ische-mia/Reperfusion Injury Induces Changes in the Drug Transporter Expression at the Blood-Brain Barrier in vivo and in vitro. Front Physiol. 2020, 11, 569881. [Google Scholar] [CrossRef]
- Ittner, C.; Burek, M.; Störk, S.; Nagai, M.; Förster, C.Y. Increased Catecholamine Levels and Inflammatory Mediators Alter Barrier Properties of Brain Microvascular Endothelial Cells in vitro. Front Cardiovasc Med. 2020, 7, 73. [Google Scholar] [CrossRef]
- Burek, M.; König, A.; Lang, M.; Fiedler, J.; Oerter, S.; Roewer, N.; Bohnert, M.; Thal, S.C.; Blecharz-Lang, K.G.; Woitzik, J.; et al. Hypoxia-Induced MicroRNA-212/132 Alter Blood-Brain Barrier Integrity Through Inhibition of Tight Junc-tion-Associated Proteins in Human and Mouse Brain Microvascular Endothelial Cells. Transl. Stroke Res. 2019, 10, 672–683. [Google Scholar] [CrossRef] [Green Version]
- Salvador, E.; Burek, M.; Förster, C.Y. Stretch and/or oxygen glucose deprivation (OGD) in an in vitro traumatic brain injury (TBI) model induces calcium alteration and inflammatory cascade. Front Cell Neurosci. 2015, 9, 323. [Google Scholar] [CrossRef]
- Salvador, E.; Burek, M.; Löhr, M.; Nagai, M.; Hagemann, C.; Förster, C.Y. Senescence and associated blood-brain barrier alterations in vitro. Histochem. Cell Biol. 2021, 156, 283–292. [Google Scholar] [CrossRef]
- Burek, M.; Steinberg, K.; Förster, C.Y. Mechanisms of transcriptional activation of the mouse claudin-5 promoter by estro-gen receptor alpha and beta. Mol. Cell Endocrinol. 2014, 392, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Blecharz, K.G.; Burek, M.; Bauersachs, J.; Thum, T.; Tsikas, D.; Widder, J.; Roewer, N.; Förster, C.Y. Inhibition of pro-teasome-mediated glucocorticoid receptor degradation restores nitric oxide bioavailability in myocardial endothelial cells in vitro. Biol. Cell 2014, 106, 219–235. [Google Scholar] [CrossRef]
- Huang, P.; Chandra, V.; Rastinejad, F. Structural overview of the nuclear receptor superfamily: Insights into physiology and therapeutics. Ann. Rev. Physiol. 2010, 72, 247–272. [Google Scholar] [CrossRef] [Green Version]
- Förster, C.; Waschke, J.; Burek, M.; Leers, J.; Drenckhahn, D. Glucocorticoid effects on mouse microvascular endothelial barrier permeability are brain specific. J. Physiol. 2006, 573, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Deroo, B.J.; Archer, T.K. Glucocorticoid receptor-mediated chromatin remodeling in vivo. Oncogene 2001, 20, 3039–3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harke, N.; Leers, J.; Kietz, S.; Drenckham, D.; Förster, C. Glucocorticoids regulate the human occludin gene through a single imperfect palindromic glucocorticoid response element. Mol. Cell. Endocrinol. 2008, 295, 39–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieplinger, B.; Bocksrucker, C.; Egger, M.; Eggers, C.; Haltmayer, M.; Mueller, T. Prognostic Value of Inflammatory and Cardiovascular Biomarkers for Prediction of 90-Day All-Cause Mortality after Acute Ischemic Stroke-Results from the Linz Stroke Unit Study. Clin. Chem. 2017, 63, 1101–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, K.L.; Lu, M.Y.; Liu, F.; Mei, Y.; Zhang, X.J.; Zhang, H.; Zan, J.; Sun, X.O.; Tan, W. Isosteviol Sodium Protects Neural Cells Against Hypoxia-Induced Apoptosis Through Inhibiting MAPK and NF-κB Pathways. J. Stroke Cerebrovasc. Dis. 2019, 28, 175–184. [Google Scholar] [CrossRef]
- Herrmann, O.; Tarabin, V.; Suzuki, S.; Attigah, N.; Coserea, I.; Schneider, A.; Vogel, J.; Prinz, S.; Schwab, S.; Monyer, H.; et al. Regulation of body temperature and neuroprotection by endogenous interleukin-6 in cerebral ischemia. J. Cereb. Blood Flow Metab. 2003, 23, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Su, J.H.; Luo, M.Y.; Liang, N.; Gong, S.X.; Chen, W.; Huang, W.Q.; Tian, Y.; Wang, A.P. Interleukin-6: A Novel Target for Cardio-Cerebrovascular Diseases. Front. Pharmacol. 2021, 12, 745061. [Google Scholar] [CrossRef]
- Lambertsen, K.L.; Biber, K.; Finsen, B. Inflammatory cytokines in experimental and human stroke. J. Cereb. Blood Flow Metab. 2012, 32, 1677–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Upadhyay, U.M.; Tamargo, R.J. Inflammation in stroke and focal cerebral ischemia. Surg. Neurol. 2006, 66, 232–245. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lin, S.; Chang, C.; Wu, C.; Chen, W.; Liao, S.; Chen, Y.; Wang, W.; Chen, C. Jak2 inhibitor AG490 improved post-stroke central and peripheral inflammation and metabolic abnormalities in a rat model of ischemic stroke. Antioxidants 2021, 10, 1958. [Google Scholar] [CrossRef] [PubMed]
- Betz, A.; Coester, H. Effect of steroids on edema and sodium uptake of the brain during focal ischemia in rats. Stroke 1990, 21, 1199–1204. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, J.; Rao, K.; Pastorino, S.; Kesari, S. Corticosteroids in brain cancer patients: Benefits and pitfalls. Expert Rev. Clin. Pharmacol. 2011, 4, 233–242. [Google Scholar] [CrossRef] [Green Version]
- Ray, A.; Prefontaine, K.E. Physical association and functional antagonism between the p65 subunit of transcription factor NF-kappa B and the glucocorticoid receptor. Proc. Natl. Acad. Sci. USA 1994, 91, 752–756. [Google Scholar] [CrossRef] [Green Version]
- Lopatkiewicz, A.M.; Gradek-Kwinta, E.; Czyzycki, M.; Pera, J.; Slowik, A.; Dziedzic, T. Glucocorticoid Resistance is Associated with Poor Functional Outcome after Stroke. Cell. Mol. Neurobiol. 2020, 40, 1321–1326. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Sobrino, M.; Rosell, A.; Penalba, A.; Ribó, M.; Alvarez-Sabín, J.; Fernández-Cadenas, I.; Montaner, J. Role of en-dogenous granulocyte-macrophage colony stimulating factor following stroke and relationship to neurological outcome. Curr. Neurovasc. Res. 2009, 6, 246–251. [Google Scholar] [CrossRef]
- Hamilton, J.A. Colony-stimulating factors in inflammation and autoimmunity. Nat. Rev. Immunol. 2008, 8, 533–544. [Google Scholar] [CrossRef]
- Sugiyama, Y.; Yagita, Y.; Oyama, N.; Terasaki, Y.; Omura-Matsuoka, E.; Sasaki, T.; Kitagawa, K. Granulocyte colo-ny-stimulating factor enhances arteriogenesis and ameliorates cerebral damage in a mouse model of ischemic stroke. Stroke 2011, 42, 770–775. [Google Scholar] [CrossRef] [Green Version]
- Faura, J.; Bustamante, A.; Miró-Mur, F.; Montaner, J. Stroke-induced immunosuppression: Implications for the prevention and prediction of post-stroke infections. J. Neuroinflamm. 2021, 18, 127. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reschke, M.; Salvador, E.; Schlegel, N.; Burek, M.; Karnati, S.; Wunder, C.; Förster, C.Y. Isosteviol Sodium (STVNA) Reduces Pro-Inflammatory Cytokine IL-6 and GM-CSF in an In Vitro Murine Stroke Model of the Blood–Brain Barrier (BBB). Pharmaceutics 2022, 14, 1753. https://doi.org/10.3390/pharmaceutics14091753
Reschke M, Salvador E, Schlegel N, Burek M, Karnati S, Wunder C, Förster CY. Isosteviol Sodium (STVNA) Reduces Pro-Inflammatory Cytokine IL-6 and GM-CSF in an In Vitro Murine Stroke Model of the Blood–Brain Barrier (BBB). Pharmaceutics. 2022; 14(9):1753. https://doi.org/10.3390/pharmaceutics14091753
Chicago/Turabian StyleReschke, Moritz, Ellaine Salvador, Nicolas Schlegel, Malgorzata Burek, Srikanth Karnati, Christian Wunder, and Carola Y. Förster. 2022. "Isosteviol Sodium (STVNA) Reduces Pro-Inflammatory Cytokine IL-6 and GM-CSF in an In Vitro Murine Stroke Model of the Blood–Brain Barrier (BBB)" Pharmaceutics 14, no. 9: 1753. https://doi.org/10.3390/pharmaceutics14091753
APA StyleReschke, M., Salvador, E., Schlegel, N., Burek, M., Karnati, S., Wunder, C., & Förster, C. Y. (2022). Isosteviol Sodium (STVNA) Reduces Pro-Inflammatory Cytokine IL-6 and GM-CSF in an In Vitro Murine Stroke Model of the Blood–Brain Barrier (BBB). Pharmaceutics, 14(9), 1753. https://doi.org/10.3390/pharmaceutics14091753