
HS 3D-SeboSkin Model Enables the Preclinical Exploration of Therapeutic Candidates for Hidradenitis Suppurativa/Acne Inversa

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. HS 3D-SeboSkin Model
2.2. Cells
2.3. Skin Specimens
2.4. Co-Culture Experiments
2.5. Tissue and Culture Supernatant Protein Extraction
2.6. Protein Quantification
2.7. Statistics
3. Results
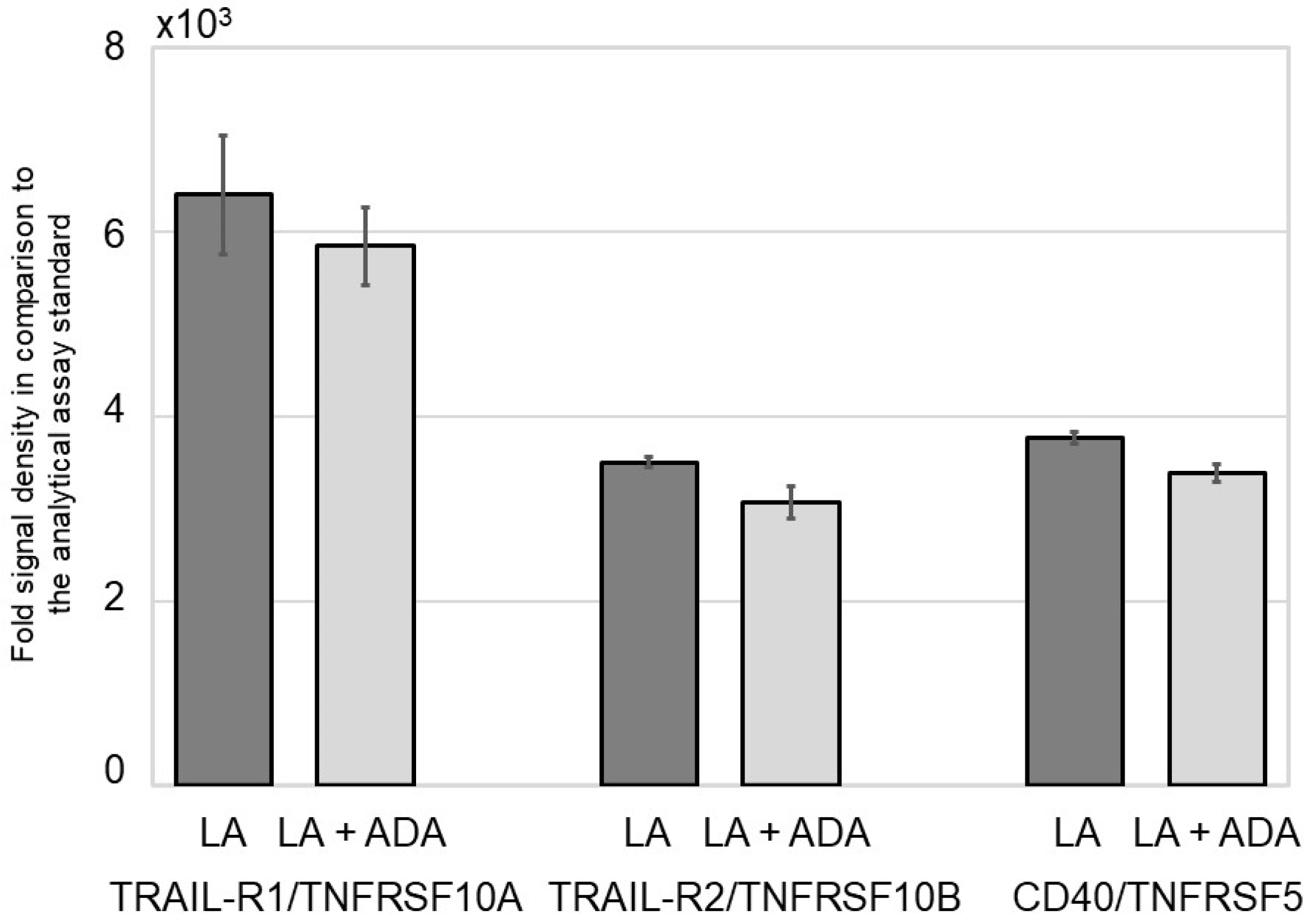
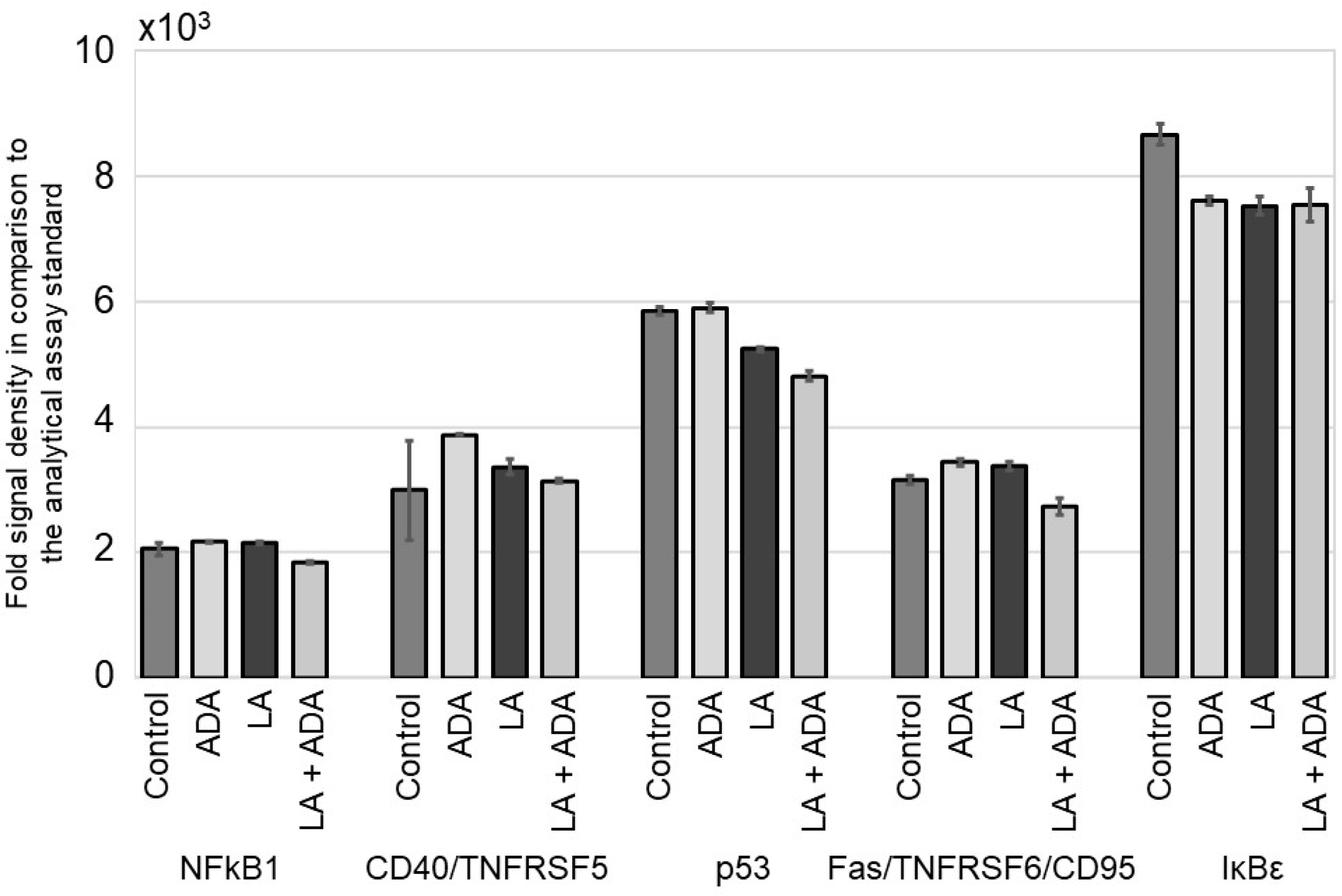
3.1. ADA Does Not Modify the NFκB Signal Pathway in Epithelial Cells
3.2. ADA Modifies Tissue Cytokine Expression in the HS 3D-SeboSkin Model
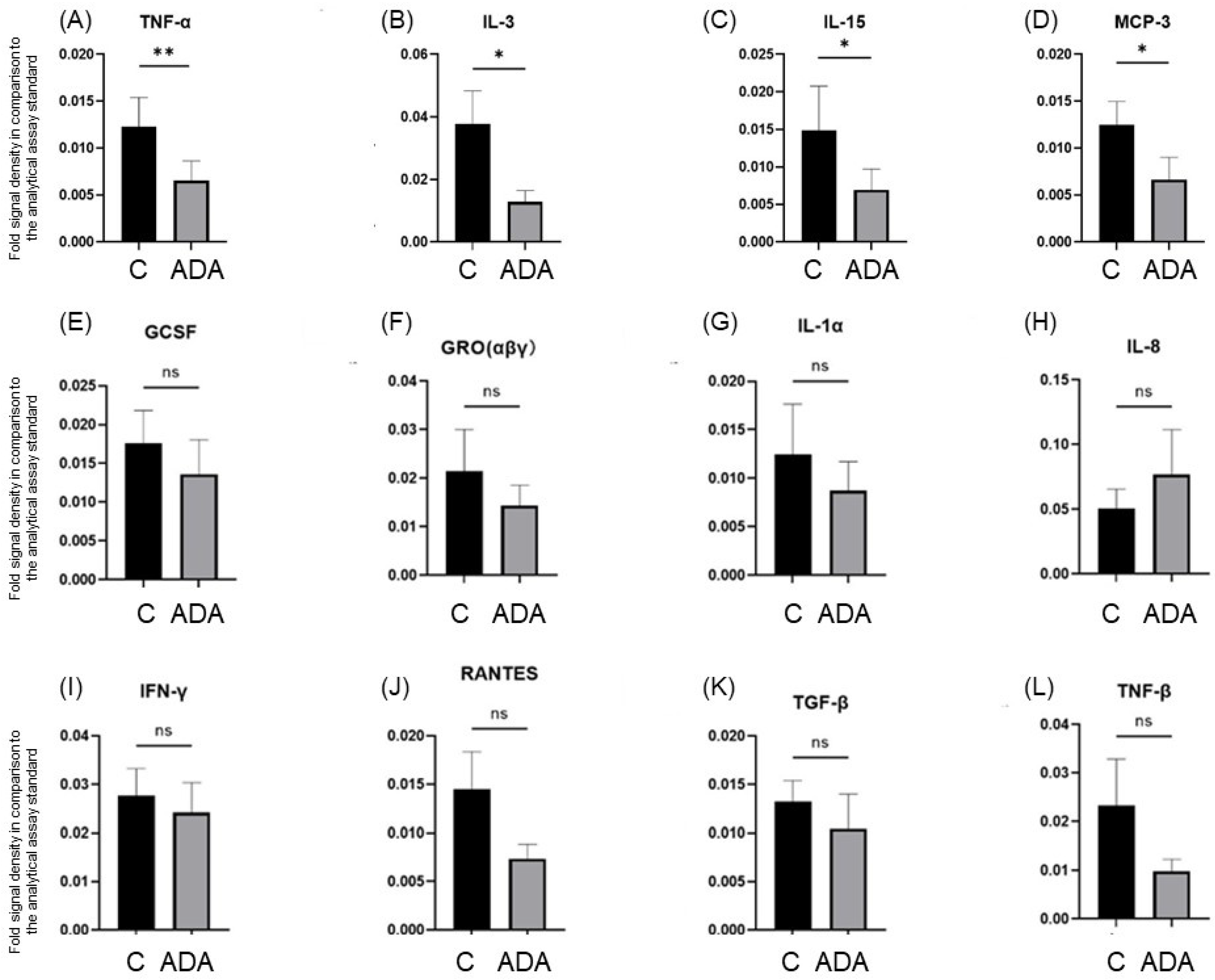
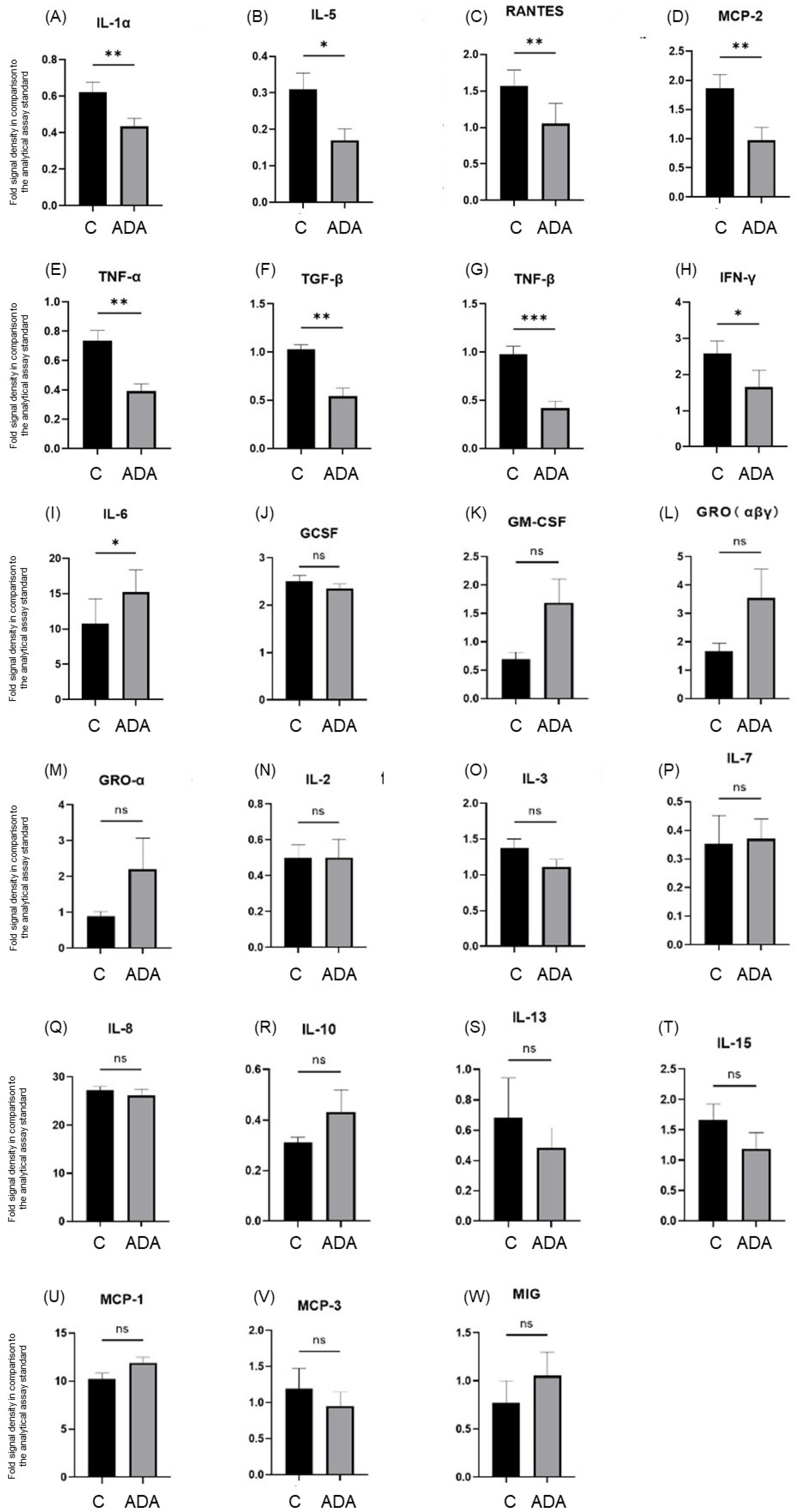
3.3. ADA Modifies Cytokine Secretion in the HS 3D-SeboSkin Model
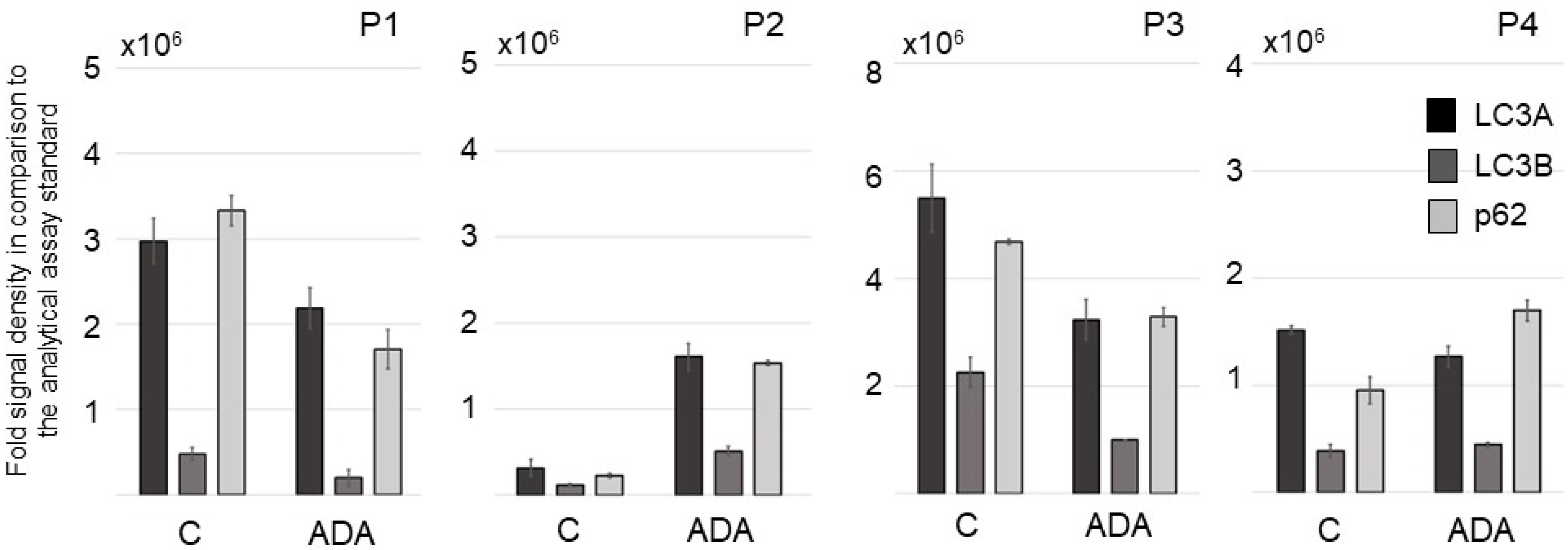
3.4. ADA Affects Autophagy Procedures in Lesional and Perilesional HS Skin Ex Vivo in the HS 3D-SeboSkin Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zouboulis, C.C.; del Marmol, V.; Mrowietz, U.; Prens, E.P.; Tzellos, T.; Jemec, G.B.E. Hidradenitis Suppurativa/Acne Inversa: Criteria for Diagnosis, Severity Assessment, Classification and Disease Evaluation. Dermatology 2015, 231, 184–190. [Google Scholar] [CrossRef]
- Kurzen, H.; Kurokawa, I.; Jemec, G.B.E.; Emtestam, L.; Sellheyer, K.; Giamarellos-Bourboulis, E.J.; Nagy, I.; Bechara, F.G.; Sartorius, K.; Lapins, J.; et al. What Causes Hidradenitis Suppurativa? Exp. Dermatol. 2008, 17, 455–472. [Google Scholar] [CrossRef]
- EHSF’s History. Available online: https://ehsf.eu/ehsf/ehsfs-history/ (accessed on 26 December 2022).
- Hidradenitis Suppurativa. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=hidradenitis+suppurativa&sort=date&size=200 (accessed on 26 December 2022).
- Jfri, A.; Nassim, D.; O’Brien, E.; Gulliver, W.; Nikolakis, G.; Zouboulis, C.C. Prevalence of Hidradenitis Suppurativa: A Systematic Review and Meta-Regression Analysis. JAMA Dermatol. 2021, 157, 924–931. [Google Scholar] [CrossRef]
- Kimball, A.B.; Okun, M.M.; Williams, D.A.; Gottlieb, A.B.; Papp, K.A.; Zouboulis, C.C.; Armstrong, A.W.; Kerdel, F.; Gold, M.H.; Forman, S.B.; et al. Two Phase 3 Trials of Adalimumab for Hidradenitis Suppurativa. N. Engl. J. Med. 2016, 375, 422–434. [Google Scholar] [CrossRef]
- Zouboulis, C.C. Adalimumab for the Treatment of Hidradenitis Suppurativa/Acne Inversa. Expert Rev. Clin. Immunol. 2016, 12, 1015–1026. [Google Scholar] [CrossRef]
- Hidradenitis Suppurativa. Available online: https://clinicaltrials.gov/ct2/results?cond=hidradenitis+suppurativa&term=&cntry=&state=&city=&dist= (accessed on 26 December 2022).
- InflaRx Provides Update on Development Plans for Vilobelimab in Hidradenitis Suppurativa. Available online: https://www.inflarx.de/Home/Investors/Press-Releases/02-2022-InflaRx-Provides-Update-on-Development-Plans-for-Vilobelimab-in-Hidradenitis-Suppurativa.html (accessed on 26 December 2022).
- TREMFYA® Treatment of Hidradenitis Suppurativa. Available online: https://www.janssenmd.com/Tremfya/clinical-use/Tremfya-treatment-of-hidradenitis-suppurativa (accessed on 26 December 2022).
- Zouboulis, C.C.; Tzellos, T.; Kyrgidis, A.; Jemec, G.B.E.; Bechara, F.G.; Giamarellos-Bourboulis, E.J.; Ingram, J.R.; Kanni, T.; Karagiannidis, I.; Martorell, A.; et al. Development and Validation of the International Hidradenitis Suppurativa Severity Score System (IHS4), a Novel Dynamic Scoring System to Assess HS Severity. Br. J. Dermatol. 2017, 177, 1401–1409. [Google Scholar] [CrossRef] [PubMed]
- Tzellos, T.; van Straalen, K.R.; Kyrgidis, A.; Alavi, A.; Goldfarb, N.; Gulliver, W.; Jemec, G.B.E.; Lowes, M.A.; Marzano, A.V.; Prens, E.P.; et al. Development and Validation of IHS4-55, an IHS4 Dichotomous Outcome to Assess Treatment Effect for Hidradenitis Suppurativa. J. Eur. Acad. Dermatol. Venereol. 2022, 37, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C. Ex Vivo Human Models of Hidradenitis Suppurativa/Acne Inversa for Laboratory Research and Drug Screening. Br. J. Dermatol. 2019, 181, 244–246. [Google Scholar] [CrossRef] [PubMed]
- Frew, J.W.; Piguet, V. Ex Vivo Models and Interpretation of Mechanistic Studies in Hidradenitis Suppurativa. J. Investig. Dermatol. 2020, 140, 1323–1326. [Google Scholar] [CrossRef]
- Hou, X.; Hossini, A.M.; Nikolakis, G.; Balthasar, O.; Kurtz, A.; Zouboulis, C.C. 3D-SeboSkin Model for Human Ex Vivo Studies of Hidradenitis Suppurativa/Acne Inversa. Dermatology 2022, 238, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Vossen, A.R.J.V.; Ardon, C.B.; van der Zee, H.H.; Lubberts, E.; Prens, E.P. The Anti-Inflammatory Potency of Biologics Targeting Tumour Necrosis Factor-α, Interleukin (IL)-17A, IL-12/23 and CD20 in Hidradenitis Suppurativa: An Ex Vivo Study. Br. J. Dermatol. 2019, 181, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J.; Le Jan, S.; Muller, C.; François, C.; Renard, Y.; Durlach, A.; Bernard, P.; Reguiai, Z.; Antonicelli, F. Matrix Remodelling and MMP Expression/Activation Are Associated with Hidradenitis Suppurativa Skin Inflammation. Exp. Dermatol. 2019, 28, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Goliwas, K.F.; Kashyap, M.P.; Khan, J.; Sinha, R.; Weng, Z.; Oak, A.S.W.; Jin, L.; Atigadda, V.; Lee, M.B.; Elmets, C.A.; et al. Ex Vivo Culture Models of Hidradenitis Suppurativa for Defining Molecular Pathogenesis and Treatment Efficacy of Novel Drugs. Inflammation 2022, 45, 1388–1401. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C.; Seltmann, H.; Neitzel, H.; Orfanos, C.E. Establishment and Characterization of an Immortalized Human Sebaceous Gland Cell Line (SZ95). J. Investig. Dermatol. 1999, 113, 1011–1020. [Google Scholar] [CrossRef]
- Nikolakis, G.; Seltmann, H.; Hossini, A.M.; Makrantonaki, E.; Knolle, J.; Zouboulis, C.C. Ex Vivo Human Skin and SZ95 Sebocytes Exhibit a Homoeostatic Interaction in a Novel Coculture Contact Model. Exp. Dermatol. 2015, 24, 497–502. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Nogueira da Costa, A.; Makrantonaki, E.; Hou, X.X.; Almansouri, D.; Dudley, J.T.; Edwards, H.; Readhead, B.; Balthasar, O.; Jemec, G.B.E.; et al. Alterations in Innate Immunity and Epithelial Cell Differentiation Are the Molecular Pillars of Hidradenitis Suppurativa. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 846–861. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Nogueira da Costa, A.; Fimmel, S.; Zouboulis, K.C. Apocrine Glands Are Bystanders in Hidradenitis Suppurativa and Their Involvement Is Gender Specific. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 1555–1563. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, V.A.; Zouboulis, K.C.; Zouboulis, C.C. Hidradenitis Suppurativa and Comorbid Disorder Biomarkers, Druggable Genes, New Drugs and Drug Repurposing-A Molecular Meta-Analysis. Pharmaceutics 2021, 14, 44. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Angres, S.; Seltmann, H. Regulation of Stearoyl-Coenzyme A Desaturase and Fatty Acid Delta-6 Desaturase-2 Expression by Linoleic Acid and Arachidonic Acid in Human Sebocytes Leads to Enhancement of Proinflammatory Activity but Does Not Affect Lipogenesis. Br. J. Dermatol. 2011, 165, 269–276. [Google Scholar] [CrossRef]
- Kelly, G.; Hughes, R.; McGarry, T.; van den Born, M.; Adamzik, K.; Fitzgerald, R.; Lawlor, C.; Tobin, A.M.; Sweeney, C.M.; Kirby, B. Dysregulated Cytokine Expression in Lesional and Nonlesional Skin in Hidradenitis Suppurativa. Br. J. Dermatol. 2015, 173, 1431–1439. [Google Scholar] [CrossRef]
- Kaleta, K.P.; Nikolakis, G.; Hossini, A.M.; Balthasar, O.; Almansouri, D.; Vaiopoulos, A.; Knolle, J.; Boguslawska, A.; Wojas-Pelc, A.; Zouboulis, C.C. Metabolic Disorders/Obesity Is a Primary Risk Factor in Hidradenitis Suppurativa: An Immunohistochemical Real-World Approach. Dermatology 2022, 238, 251–259. [Google Scholar] [CrossRef]
- Kabeya, Y.; Mizushima, N.; Ueno, T.; Yamamoto, A.; Kirisako, T.; Noda, T.; Kominami, E.; Ohsumi, Y.; Yoshimori, T. LC3, a Mammalian Homologue of Yeast Apg8p, Is Localized in Autophagosome Membranes after Processing. EMBO J. 2000, 19, 5720–5728. [Google Scholar] [CrossRef] [PubMed]
- Vossen, A.R.J.V.; van der Zee, H.H.; Davelaar, N.; Mus, A.M.C.; van Doorn, M.B.A.; Prens, E.P. Apremilast for Moderate Hidradenitis Suppurativa: No Significant Change in Lesional Skin Inflammatory Biomarkers. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Degner, J.; Georgantas, R.W.; Nader, A.; Mostafa, N.M.; Teixeira, H.D.; Williams, D.A.; Kirsner, R.S.; Nichols, A.J.; Davis, J.W.; et al. A Genetic Variant in the BCL2 Gene Associates with Adalimumab Response in Hidradenitis Suppurativa Clinical Trials and Regulates Expression of BCL2. J. Investig. Dermatol. 2020, 140, 574–582.e2. [Google Scholar] [CrossRef]
- Lowe, M.M.; Naik, H.B.; Clancy, S.; Pauli, M.; Smith, K.M.; Bi, Y.; Dunstan, R.; Gudjonsson, J.E.; Paul, M.; Harris, H.; et al. Immunopathogenesis of Hidradenitis Suppurativa and Response to Anti-TNF-α Therapy. JCI Insight 2020, 5, e139932. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Harvey, B.P.; Hong, F.; Ruzek, M.; Wang, J.; Murphy, E.R.; Kaymakcalan, Z. Adalimumab Induces a Wound Healing Profile in Patients with Hidradenitis Suppurativa by Regulating Macrophage Differentiation and Matrix Metalloproteinase Expression. J. Investig. Dermatol. 2021, 141, 2730–2740.e9. [Google Scholar] [CrossRef] [PubMed]
- Navrazhina, K.; Frew, J.W.; Grand, D.; Williams, S.C.; Hur, H.; Gonzalez, J.; Garcet, S.; Krueger, J.G. Interleukin-17RA Blockade by Brodalumab Decreases Inflammatory Pathways in Hidradenitis Suppurativa Skin and Serum. Br. J. Dermatol. 2022, 187, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-ΚB Pathway for the Therapy of Diseases: Mechanism and Clinical Study. Signal Transduct. Target. Ther. 2020, 5, 209. [Google Scholar] [CrossRef]
- Gulliver, W.; Alavi, A.; Wiseman, M.C.; Gooderham, M.J.; Rao, J.; Alam, M.S.; Papp, K.A.; Desjardins, O.; Jean, C. Real-World Effectiveness of Adalimumab in Patients with Moderate-to-Severe Hidradenitis Suppurativa: The 1-Year SOLACE Study. J. Eur. Acad. Dermatol. Venereol. 2021, 35, 2431–2439. [Google Scholar] [CrossRef]
- Frew, J.W.; Jiang, C.S.; Singh, N.; Grand, D.; Navrazhina, K.; Vaughan, R.; Krueger, J.G. Dermal Tunnels Influence Time to Clinical Response and Family History Influences Time to Loss of Clinical Response in Patients with Hidradenitis Suppurativa Treated with Adalimumab. Clin. Exp. Dermatol. 2021, 46, 306–313. [Google Scholar] [CrossRef]
- Caposiena Caro, R.D.; Solivetti, F.M.; Candi, E.; Bianchi, L. Clinical and Power-Doppler Ultrasound Features Related with Persistence of Fistulous Tracts under Treatment with Adalimumab in Hidradenitis Suppurativa: 4 Years of Follow-Up. Dermatol. Ther. 2021, 34, e14804. [Google Scholar] [CrossRef]
- Fujishima, S.; Watanabe, H.; Kawaguchi, M.; Suzuki, T.; Matsukura, S.; Homma, T.; Howell, B.G.; Hizawa, N.; Mitsuya, T.; Huang, S.-K.; et al. Involvement of IL-17F via the Induction of IL-6 in Psoriasis. Arch. Dermatol. Res. 2010, 302, 499–505. [Google Scholar] [CrossRef]
- Pietrzak, A.T.; Zalewska, A.; Chodorowska, G.; Krasowska, D.; Michalak-Stoma, A.; Nockowski, P.; Osemlak, P.; Paszkowski, T.; Roliński, J.M. Cytokines and Anticytokines in Psoriasis. Clin. Chim. Acta Int. J. Clin. Chem. 2008, 394, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Shuwa, H.A.; Dallatu, M.K.; Yeldu, M.H.; Ahmed, H.M.; Nasir, I.A. Effects of Adalimumab, an Anti-Tumour Necrosis Factor-Alpha (TNF-α) Antibody, on Obese Diabetic Rats. Malays. J. Med. Sci. 2018, 25, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Gallo, D.; de la Varga-Martínez, R.; Ossorio-García, L.; Collantes-Rodríguez, C.; Rodríguez, C.; Linares-Barrios, M. Effects of Adalimumab on T-Helper-17 Lymphocyte- and Neutrophil-Related Inflammatory Serum Markers in Patients with Moderate-to-Severe Hidradenitis Suppurativa. Cytokine 2018, 103, 20–24. [Google Scholar] [CrossRef]
- Olejniczak-Staruch, I.; Narbutt, J.; Bednarski, I.; Woźniacka, A.; Sieniawska, J.; Kraska-Gacka, M.; Śmigielski, J.; Lesiak, A. Interleukin 22 and 6 Serum Concentrations Decrease under Long-Term Biologic Therapy in Psoriasis. Postepy Dermatol. Alergol. 2020, 37, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.H.; Jung, J.Y.; Park, K.; Hossini, A.M.; Zouboulis, C.C.; Lee, S.E. Autophagy Regulates Lipid Production and Contributes to the Sebosuppressive Effect of Retinoic Acid in Human SZ95 Sebocytes. J. Dermatol. Sci. 2020, 98, 128–136. [Google Scholar] [CrossRef]
- Lee, Y.; Shin, K.; Shin, K.-O.; Yoon, S.; Jung, J.; Hwang, E.; Chung, H.-J.; Hossini, A.M.; Zouboulis, C.C.; Baek, M.J.; et al. Topical Application of Autophagy-Activating Peptide Improved Skin Barrier Function and Reduced Acne Symptoms in Acne-Prone Skin. J. Cosmet. Dermatol. 2021, 20, 1009–1016. [Google Scholar] [CrossRef]
- Hossini, A.M.; Hou, X.; Exner, T.; Fauler, B.; Eberle, J.; Rabien, A.; Makrantonaki, E.; Zouboulis, C.C. Free Fatty Acids Induce Lipid Accumulation, Autophagy and Apoptosis in Human Sebocytes. Skin Pharmacol. Physiol. 2022, 1–5. [Google Scholar] [CrossRef]
- Bechara, F.G.; Podda, M.; Prens, E.P.; Horváth, B.; Giamarellos-Bourboulis, E.J.; Alavi, A.; Szepietowski, J.C.; Kirby, J.; Geng, Z.; Jean, C.; et al. Efficacy and Safety of Adalimumab in Conjunction With Surgery in Moderate to Severe Hidradenitis Suppurativa: The SHARPS Randomized Clinical Trial. JAMA Surg. 2021, 156, 1001–1009. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zouboulis, C.C.; Hou, X.; von Waldthausen, H.; Zouboulis, K.C.; Hossini, A.M. HS 3D-SeboSkin Model Enables the Preclinical Exploration of Therapeutic Candidates for Hidradenitis Suppurativa/Acne Inversa. Pharmaceutics 2023, 15, 619. https://doi.org/10.3390/pharmaceutics15020619
Zouboulis CC, Hou X, von Waldthausen H, Zouboulis KC, Hossini AM. HS 3D-SeboSkin Model Enables the Preclinical Exploration of Therapeutic Candidates for Hidradenitis Suppurativa/Acne Inversa. Pharmaceutics. 2023; 15(2):619. https://doi.org/10.3390/pharmaceutics15020619
Chicago/Turabian StyleZouboulis, Christos C., Xiaoxiao Hou, Henriette von Waldthausen, Konstantin C. Zouboulis, and Amir M. Hossini. 2023. "HS 3D-SeboSkin Model Enables the Preclinical Exploration of Therapeutic Candidates for Hidradenitis Suppurativa/Acne Inversa" Pharmaceutics 15, no. 2: 619. https://doi.org/10.3390/pharmaceutics15020619
APA StyleZouboulis, C. C., Hou, X., von Waldthausen, H., Zouboulis, K. C., & Hossini, A. M. (2023). HS 3D-SeboSkin Model Enables the Preclinical Exploration of Therapeutic Candidates for Hidradenitis Suppurativa/Acne Inversa. Pharmaceutics, 15(2), 619. https://doi.org/10.3390/pharmaceutics15020619