


Tailoring the Lamellarity of Liposomes Prepared by Dual Centrifugation

 , ,
, , 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
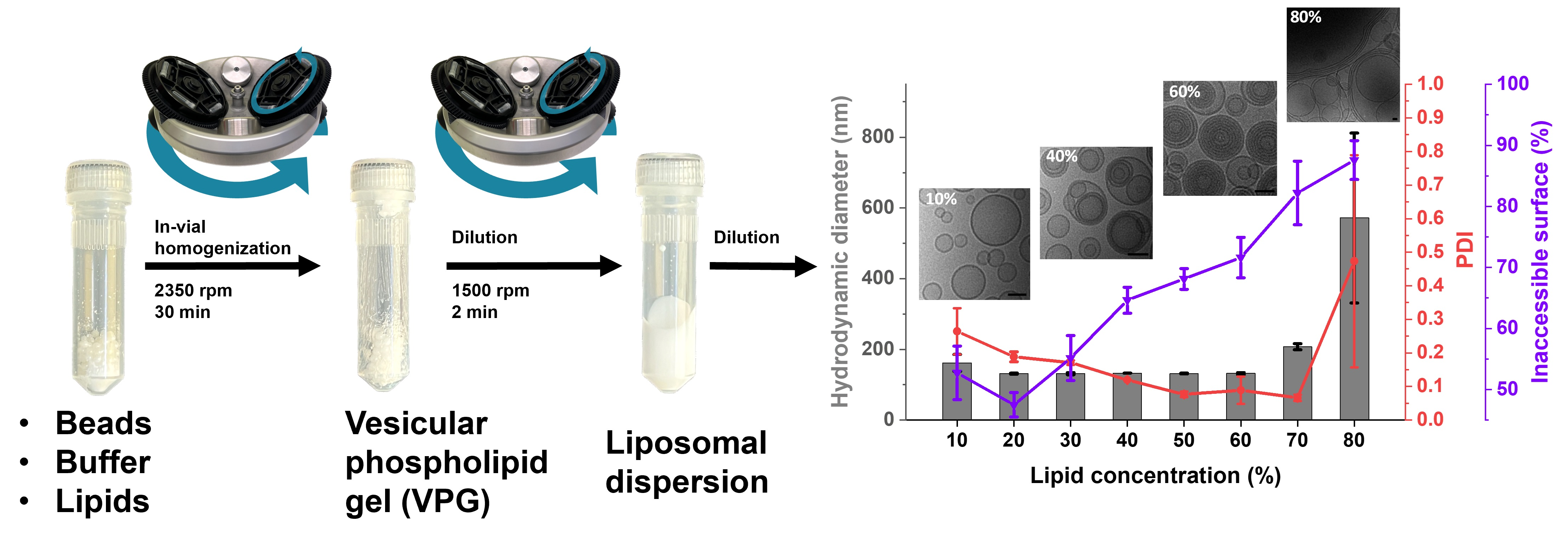
2.2.1. Preparation of Liposomes with Prior Lipid Film Preparation
2.2.2. Liposome Preparation without Lipid Film Preparation
2.2.3. Liposome Size and Size Distribution
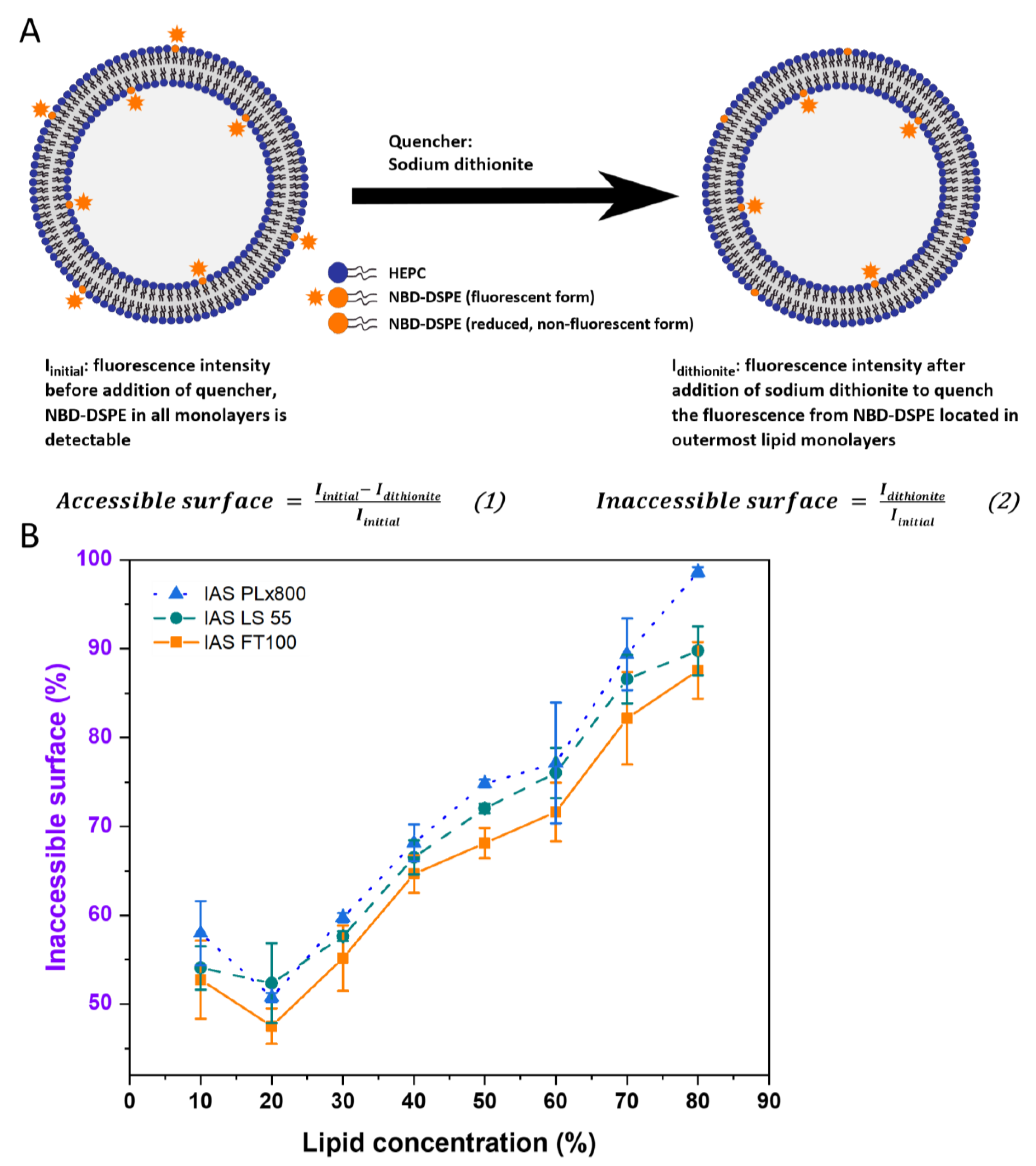
2.2.4. Inaccessible Surface Measurements
Time-Resolved Fluorescence (TRF) Measurements: FT100
Steady State Fluorescence Measurements: LS55
Steady State Fluorescence Measurements in a Plate Reader: PLx800
2.2.5. Uptake of Calcein into Open Lamellar Structures
2.2.6. Cryo-EM Specimen Preparation and Analysis
3. Results
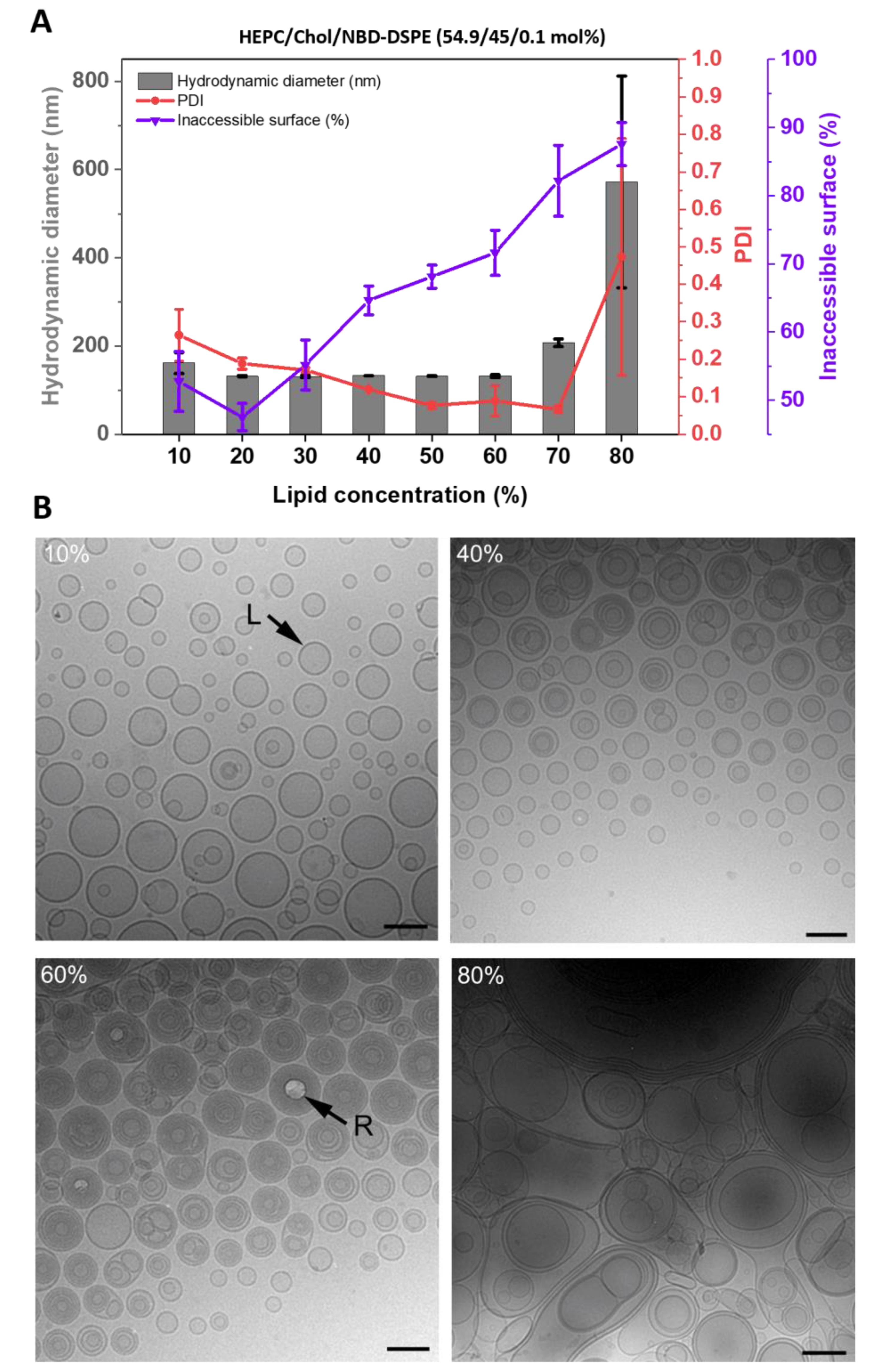
3.1. Morphology of Cholesterol-Containing Liposomes
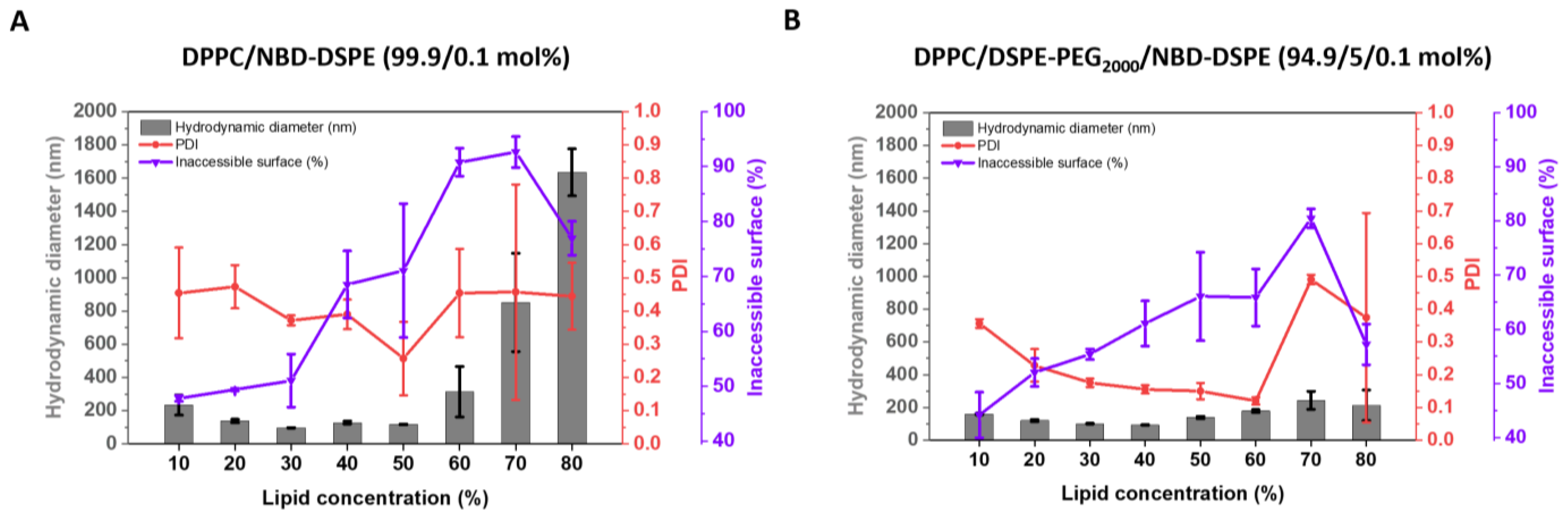
3.1.1. Fluorescence-Based Inaccessible Surface Assay
3.1.2. Cryo-EM
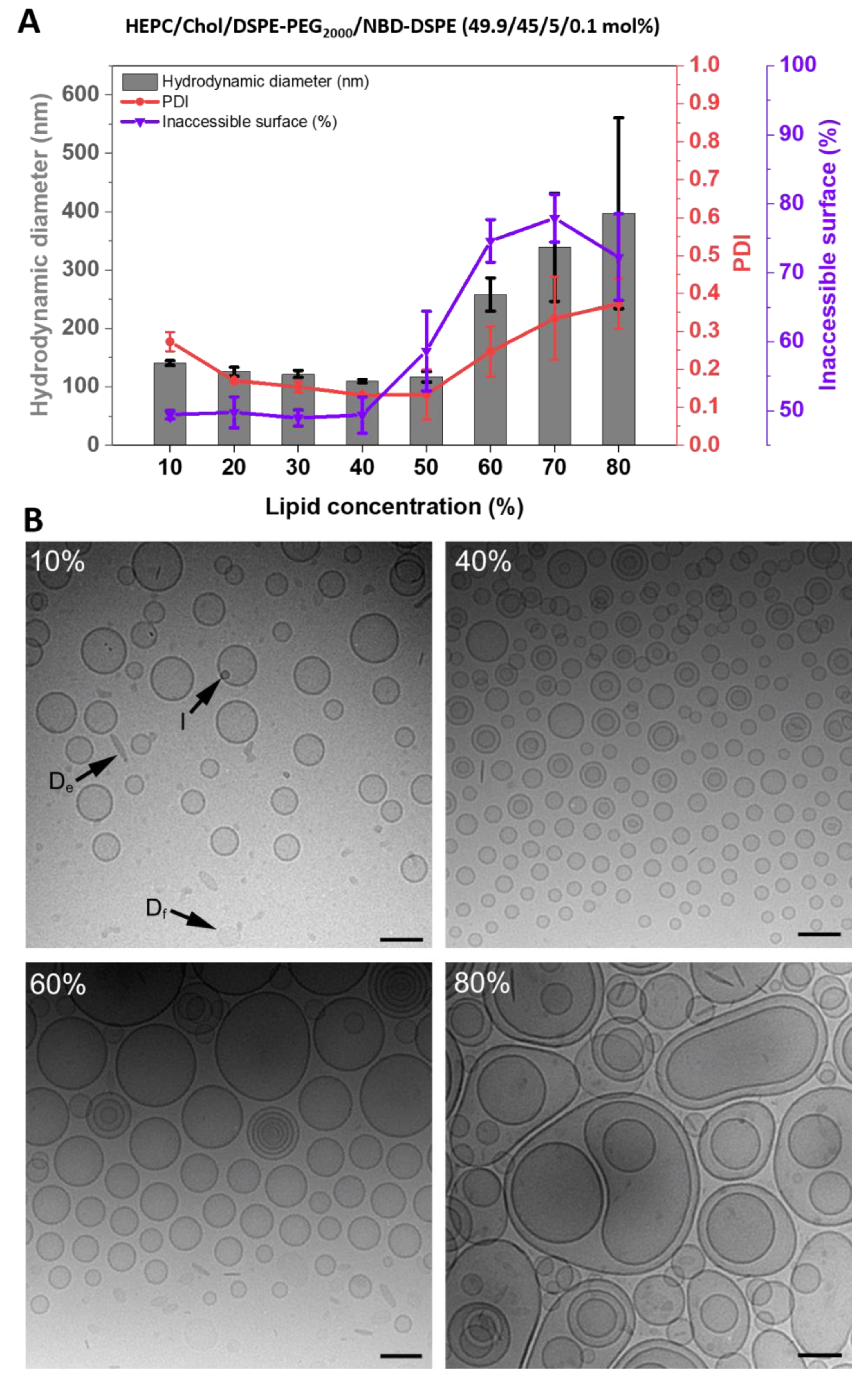
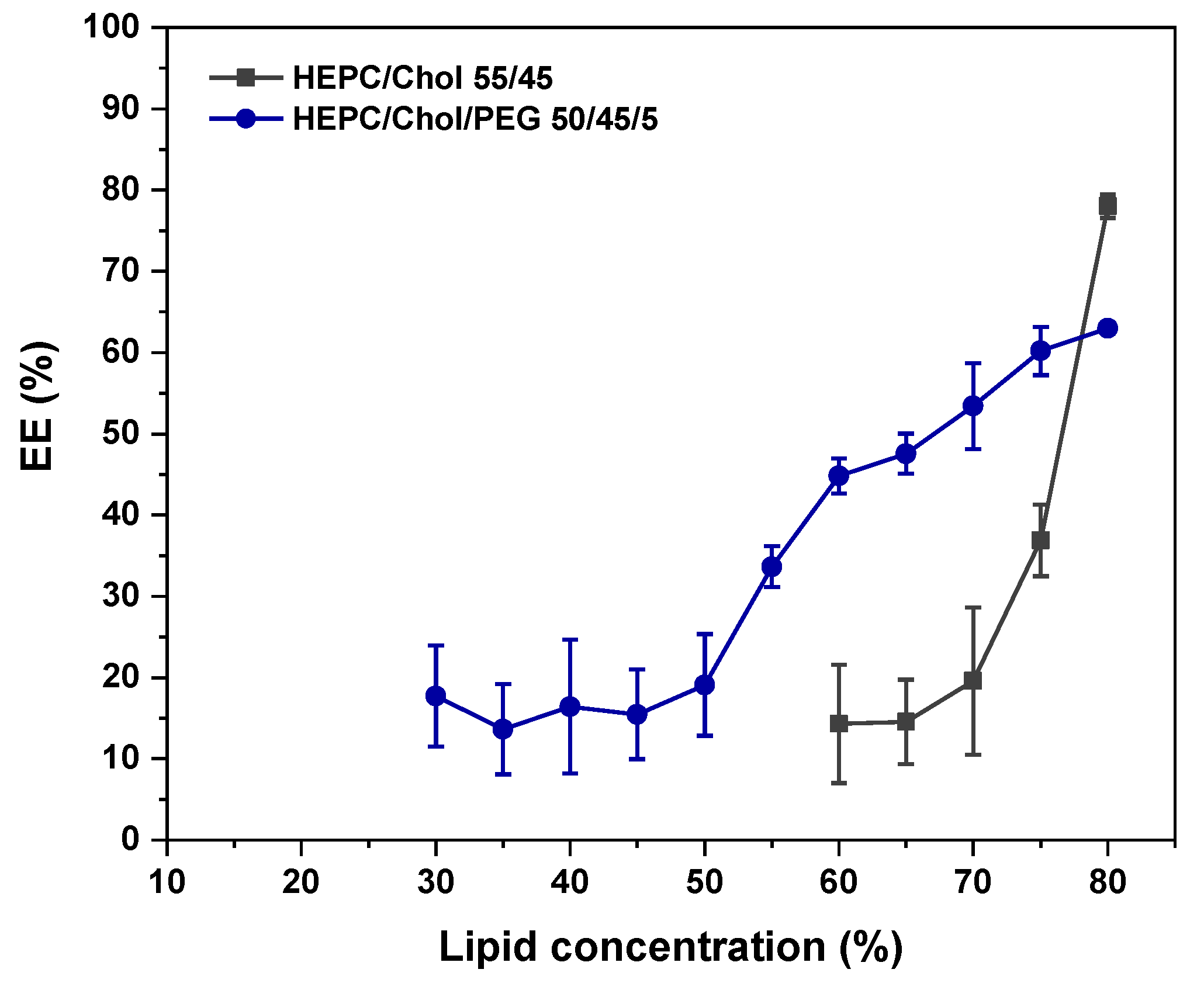
3.1.3. Effects of Adding 5 mol% DSPE-PEG2000 to the HEPC/Chol Lipid Mixtures
3.1.4. Open Membrane Stack Structures in VPGs
3.1.5. Liposome Formation without Lipid Film Preparation in Organic Solvents
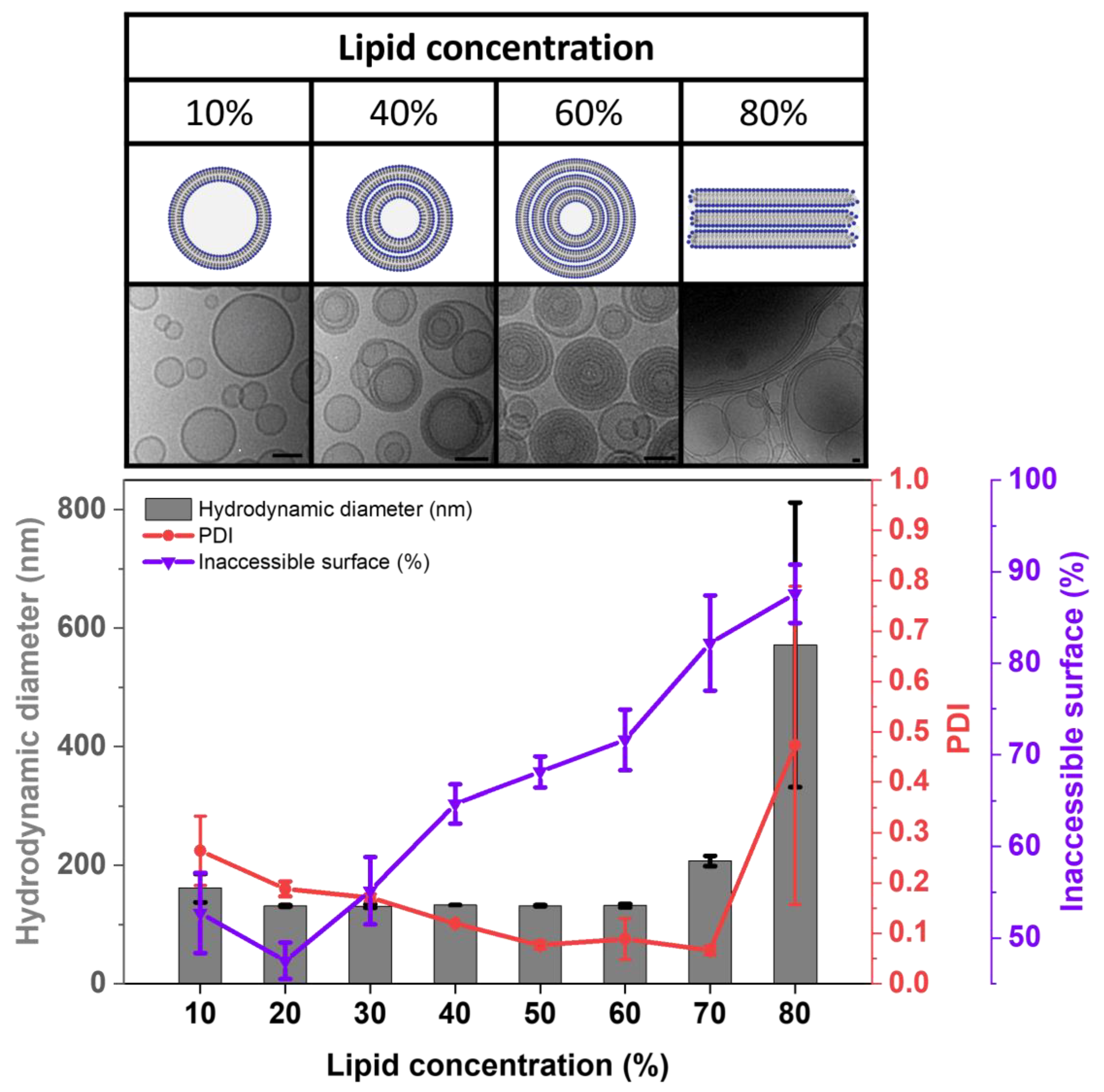
3.2. Morphology of Cholesterol-Free Liposomes
3.2.1. Investigations Based on IAS Measurements
3.2.2. Investigations Based on Cryo-EM Images
4. Discussion
4.1. Fluorescence-Based Lamellarity Assay—Opportunities and Limitations
4.2. Tailoring the Lamellarity of Small Liposomes
4.3. The Shortage of Water Limits the Formation of Closed Vesicles
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tian, W.; Schulze, S.; Brandl, M.; Winter, G. Vesicular Phospholipid Gel-Based Depot Formulations for Pharmaceutical Proteins: Development and in Vitro Evaluation. J. Control. Release 2010, 142, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Koehler, J.K.; Schnur, J.; Heerklotz, H.; Massing, U. Screening for Optimal Liposome Preparation Conditions by Using Dual Centrifugation and Time-Resolved Fluorescence Measurements. Pharmaceutics 2021, 13, 2046. [Google Scholar] [CrossRef] [PubMed]
- Massing, U.; Cicko, S.; Ziroli, V. Dual Asymmetric Centrifugation (DAC)—A New Technique for Liposome Preparation. J. Control. Release Off. J. Control. Release Soc. 2008, 125, 16–24. [Google Scholar] [CrossRef]
- Massing, U.; Ingebrigtsen, S.G.; Škalko-Basnet, N.; Holsæter, A.M. Dual Centrifugation—A Novel “in-Vial” Liposome Processing Technique. In Liposomes; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Deuringer, B.; Härdtner, C.; Krebs, K.; Thomann, R.; Holzer, M.; Hilgendorf, I.; Süss, R. Everolimus-Loaded Reconstituted High-Density Lipoprotein Prepared by a Novel Dual Centrifugation Approach for Anti-Atherosclerotic Therapy. Int. J. Nanomed. 2022, 17, 5081–5097. [Google Scholar] [CrossRef]
- Langer, C.; Köll-Weber, M.; Holzer, M.; Hantel, C.; Süss, R. Mitotane Nanocarriers for the Treatment of Adrenocortical Carcinoma: Evaluation of Albumin-Stabilized Nanoparticles and Liposomes in a Preclinical In Vitro Study with 3D Spheroids. Pharmaceutics 2022, 14, 1891. [Google Scholar] [CrossRef] [PubMed]
- Hagedorn, M.; Bögershausen, A.; Rischer, M.; Schubert, R.; Massing, U. Dual Centrifugation—A New Technique for Nanomilling of Poorly Soluble Drugs and Formulation Screening by an DoE-Approach. Int. J. Pharm. 2017, 530, 79–88. [Google Scholar] [CrossRef]
- Willmann, A.-C.; Berkenfeld, K.; Faber, T.; Wachtel, H.; Boeck, G.; Wagner, K.G. Itraconazole Nanosuspensions via Dual Centrifugation Media Milling: Impact of Formulation and Process Parameters on Particle Size and Solid-State Conversion as Well as Storage Stability. Pharmaceutics 2022, 14, 1528. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Staufenbiel, S.; Bodmeier, R. Kinetic Solubility Improvement and Influence of Polymers on Controlled Supersaturation of Itraconazole-Succinic Acid Nano-Co-Crystals. Int. J. Pharm. 2022, 616, 121536. [Google Scholar] [CrossRef]
- Steiner, D.; Bunjes, H. Influence of Process and Formulation Parameters on the Preparation of Solid Lipid Nanoparticles by Dual Centrifugation. Int. J. Pharm. X 2021, 3, 100085. [Google Scholar] [CrossRef]
- Steiner, D.; Emmendörffer, J.F.; Bunjes, H. Orodispersible Films: A Delivery Platform for Solid Lipid Nanoparticles? Pharmaceutics 2021, 13, 2162. [Google Scholar] [CrossRef]
- Pantze, S.F.; Parmentier, J.; Hofhaus, G.; Fricker, G. Matrix Liposomes: A Solid Liposomal Formulation for Oral Administration. Eur. J. Lipid Sci. Technol. 2014, 116, 1145–1154. [Google Scholar] [CrossRef]
- Binnefeld, M.; Fritz, S.; Balzer, V.; Skalická, V.; Witzigmann, D.; Kauczor, H.-U.; Fricker, G.; Salomon, J.J. Physicochemical and Biopharmaceutical Characterization of Novel Matrix-Liposomes. Eur. J. Pharm. Biopharm. 2020, 153, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Krämer, W.; Schubert, R.; Massing, U. Small-Scale Preparation of Perfluorocarbon-Nanoemulsions Utilizing Dual Centrifugation. Int. J. Pharm. 2019, 572, 118753. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, J.C.; Sleight, R.G. Fluorescence Assay for Phospholipid Membrane Asymmetry. Biochemistry 1991, 30, 11819–11827. [Google Scholar] [CrossRef]
- Gruber, H.J.; Schindler, H. External Surface and Lamellarity of Lipid Vesicles: A Practice-Oriented Set of Assay Methods. Biochim. Biophys. Acta Biomembr. 1994, 1189, 212–224. [Google Scholar] [CrossRef]
- Almgren, M.; Edwards, K.; Karlsson, G. Cryo Transmission Electron Microscopy of Liposomes and Related Structures. Colloids Surf. A Physicochem. Eng. Asp. 2000, 174, 3–21. [Google Scholar] [CrossRef]
- Zetterberg, M.M.; Ahlgren, S.; Agmo Hernández, V.; Parveen, N.; Edwards, K. Optimization of Lipodisk Properties by Modification of the Extent and Density of the PEG Corona. J. Colloid Interface Sci. 2016, 484, 86–96. [Google Scholar] [CrossRef]
- Ruppel, D.; Sackmann, E. On Defects in Different Phases of Two-Dimensional Lipid Bilayers. J. Phys. France 1983, 44, 1025–1034. [Google Scholar] [CrossRef]
- Ickenstein, L.M.; Arfvidsson, M.C.; Needham, D.; Mayer, L.D.; Edwards, K. Disc Formation in Cholesterol-Free Liposomes during Phase Transition. Biochim. Biophys. Acta Biomembr. 2003, 1614, 135–138. [Google Scholar] [CrossRef] [Green Version]
- Needham, D.; Dewhirst, M.W. The Development and Testing of a New Temperature-Sensitive Drug Delivery System for the Treatment of Solid Tumors. Adv. Drug Deliv. Rev. 2001, 53, 285–305. [Google Scholar] [CrossRef]
- Fröhlich, M.; Brecht, V.; Peschka-Süss, R. Parameters Influencing the Determination of Liposome Lamellarity by 31P-NMR. Chem. Phys. Lipids 2001, 109, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, N.; Kimpfler, A.; Massing, U.; Burger, A.M.; Fiebig, H.H.; Brandl, M.; Schubert, R. 5-Fluorouracil in Vesicular Phospholipid Gels for Anticancer Treatment: Entrapment and Release Properties. Int. J. Pharm. 2003, 256, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Tardi, C.; Drechsler, M.; Bauer, K.H.; Brandl, M. Steam Sterilisation of Vesicular Phospholipid Gels. Int. J. Pharm. 2001, 217, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Barba, A.A.; Bochicchio, S.; Dalmoro, A.; Lamberti, G. Lipid Delivery Systems for Nucleic-Acid-Based-Drugs: From Production to Clinical Applications. Pharmaceutics 2019, 11, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirsch, M.; Ziroli, V.; Helm, M.; Massing, U. Preparation of Small Amounts of Sterile SiRNA-Liposomes with High Entrapping Efficiency by Dual Asymmetric Centrifugation (DAC). J. Control. Release 2009, 135, 80–88. [Google Scholar] [CrossRef]
- Adrian, J.E.; Wolf, A.; Steinbach, A.; Rössler, J.; Süss, R. Targeted Delivery to Neuroblastoma of Novel SiRNA-Anti-GD2-Liposomes Prepared by Dual Asymmetric Centrifugation and Sterol-Based Post-Insertion Method. Pharm. Res. 2011, 28, 2261. [Google Scholar] [CrossRef]
- Maeda, H.; Wu, J.; Sawa, T.; Matsumura, Y.; Hori, K. Tumor Vascular Permeability and the EPR Effect in Macromolecular Therapeutics: A Review. J. Control. Release 2000, 65, 271–284. [Google Scholar] [CrossRef]
- Gerlowski, L.E.; Jain, R.K. Microvascular Permeability of Normal and Neoplastic Tissues. Microvasc. Res. 1986, 31, 288–305. [Google Scholar] [CrossRef]
- Vorselen, D.; van Dommelen, S.M.; Sorkin, R.; Piontek, M.C.; Schiller, J.; Döpp, S.T.; Kooijmans, S.A.A.; van Oirschot, B.A.; Versluijs, B.A.; Bierings, M.B.; et al. The Fluid Membrane Determines Mechanics of Erythrocyte Extracellular Vesicles and Is Softened in Hereditary Spherocytosis. Nat. Commun. 2018, 9, 4960. [Google Scholar] [CrossRef] [Green Version]
- Breitsamer, M.; Winter, G. Vesicular Phospholipid Gels as Drug Delivery Systems for Small Molecular Weight Drugs, Peptides and Proteins: State of the Art Review. Int. J. Pharm. 2019, 557, 1–8. [Google Scholar] [CrossRef]
- Tardi, C.; Brandl, M.; Schubert, R. Erosion and Controlled Release Properties of Semisolid Vesicular Phospholipid Dispersions. J. Control. Release 1998, 55, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Tu, K.; Tobias, D.J.; Blasie, J.K.; Klein, M.L. Molecular Dynamics Investigation of the Structure of a Fully Hydrated Gel-Phase Dipalmitoylphosphatidylcholine Bilayer. Biophys. J. 1996, 70, 595–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, H.; Kohlstrunk, B.; Heerklotz, H.H. A Humidity Titration Calorimetry Technique to Study the Thermodynamics of Hydration. Chem. Phys. Lett. 1999, 304, 329–335. [Google Scholar] [CrossRef]
- Jendrasiak, G.L.; Smith, R.L.; Shaw, W. The Water Adsorption Characteristics of Charged Phospholipids. Biochim. Biophys. Acta Biomembr. 1996, 1279, 63–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grad, P.; Gedda, L.; Edwards, K. Effect of Gangliosides on Structure and Integrity of Polyethylene Glycol (PEG)-Stabilized Liposomes. J. Colloid Interface Sci. 2020, 578, 281–289. [Google Scholar] [CrossRef]
- Johansson, E.; Engvall, C.; Arfvidsson, M.; Lundahl, P.; Edwards, K. Development and Initial Evaluation of PEG-Stabilized Bilayer Disks as Novel Model Membranes. Biophys. Chem. 2005, 113, 183–192. [Google Scholar] [CrossRef]
- Edwards, K.; Johnsson, M.; Karlsson, G.; Silvander, M. Effect of Polyethyleneglycol-Phospholipids on Aggregate Structure in Preparations of Small Unilamellar Liposomes. Biophys. J. 1997, 73, 258–266. [Google Scholar] [CrossRef] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koehler, J.K.; Gedda, L.; Wurster, L.; Schnur, J.; Edwards, K.; Heerklotz, H.; Massing, U. Tailoring the Lamellarity of Liposomes Prepared by Dual Centrifugation. Pharmaceutics 2023, 15, 706. https://doi.org/10.3390/pharmaceutics15020706
Koehler JK, Gedda L, Wurster L, Schnur J, Edwards K, Heerklotz H, Massing U. Tailoring the Lamellarity of Liposomes Prepared by Dual Centrifugation. Pharmaceutics. 2023; 15(2):706. https://doi.org/10.3390/pharmaceutics15020706
Chicago/Turabian StyleKoehler, Jonas K., Lars Gedda, Leonie Wurster, Johannes Schnur, Katarina Edwards, Heiko Heerklotz, and Ulrich Massing. 2023. "Tailoring the Lamellarity of Liposomes Prepared by Dual Centrifugation" Pharmaceutics 15, no. 2: 706. https://doi.org/10.3390/pharmaceutics15020706
APA StyleKoehler, J. K., Gedda, L., Wurster, L., Schnur, J., Edwards, K., Heerklotz, H., & Massing, U. (2023). Tailoring the Lamellarity of Liposomes Prepared by Dual Centrifugation. Pharmaceutics, 15(2), 706. https://doi.org/10.3390/pharmaceutics15020706