

The Potential Use of Exosomes in Anti-Cancer Effect Induced by Polarized Macrophages



Abstract
:1. Introduction
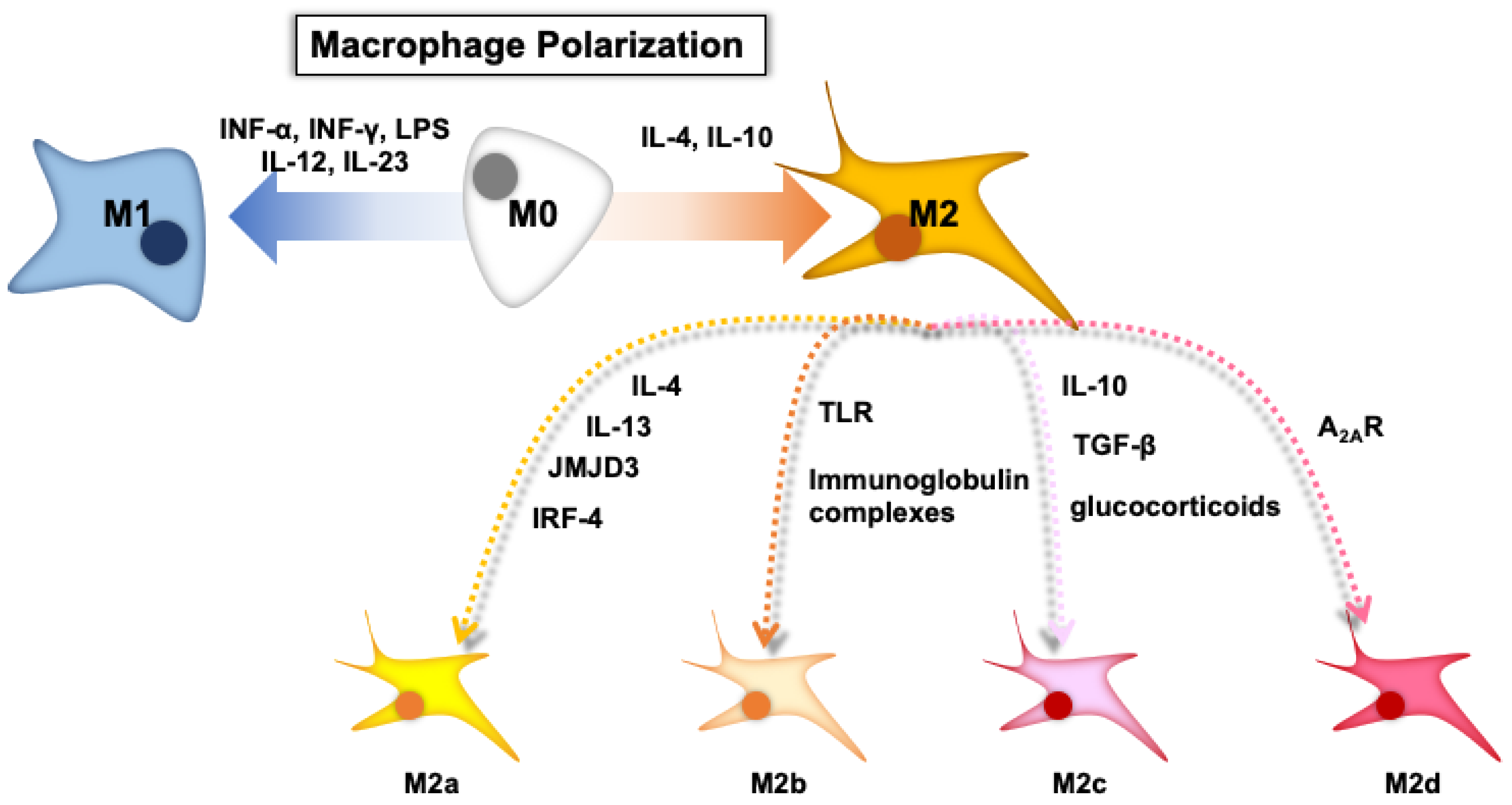
2. Macrophage Polarization
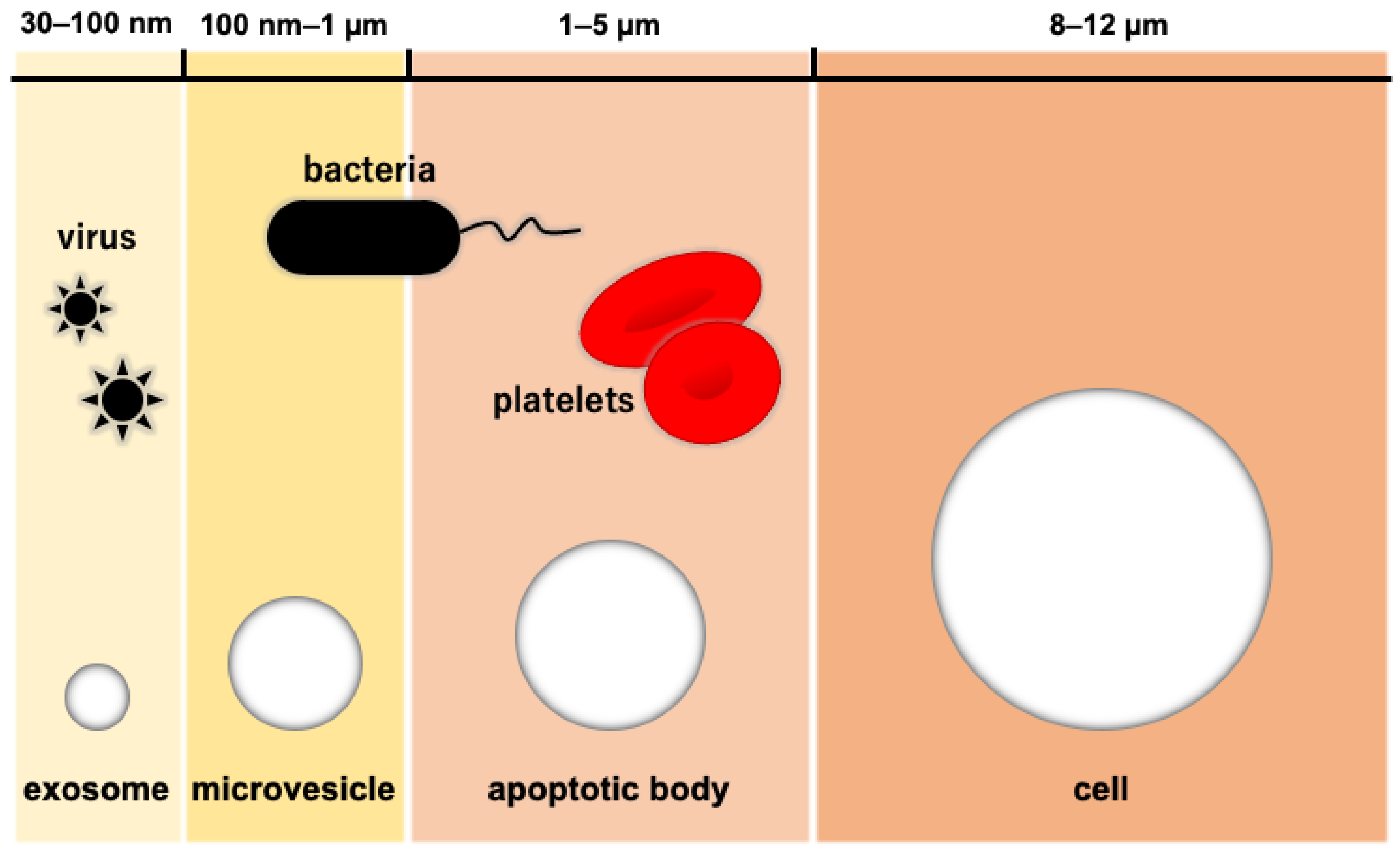
3. Exosome
3.1. The Potential Use of Exosomes for Therapy
3.2. The Potential Use of Exosomes for Diagnostics of Cancers
4. Exosome-Induced Macrophage Polarization in Cancers
4.1. M1/M2 Macrophages in Cancers
4.2. The Role of Tumor-Induced Exosomes in Macrophage Polarization
4.3. Anticancer Effects by Exosome-Induced Macrophage Polarization
5. Perspective and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMACR | an enzyme a-Methylacyl-CoA racemase |
| M2 | anti-inflammatory macrophage |
| AUC | area under the curve |
| CAFs | cancer-associated fibroblasts |
| circRNAs | circular RNAs |
| ccRCC | clear cell RCC |
| C3G | cyanidin-3-galactoside |
| DA | dopamine |
| EGCG | epigallocatechin-3-gallate |
| DSS | dextran sodium sulfate |
| ADSC-Exos | exosome from adipose-derived stem cells |
| EXPLORs | exosomes for protein loading via optically reversible protein–protein interactions |
| EVs | extracellular vesicles |
| GC | gastric cancer |
| HNSCC | head and neck squamous cell carcinoma |
| HCG18 | HLA complex group 18 |
| IRF | interferon regulatory factor |
| IRF1 | interferon-regulatory factor 1 |
| INF-γ | interferon-γ |
| IL | Interleukin |
| JMJD3 | jumonji domain-containing-3 |
| LPS | Lipopolysaccharides |
| Lnc RNA | long non-cording RNA |
| LUAD | lung adenocarcinoma |
| miRNAs | microRNAs |
| M0 | naïve macrophage |
| NSCLC | non-small cell lung cancer |
| PEDF | pigment epithelium-derived factor |
| M1 | pro-inflammatory macrophage |
| PCa | prostate cancer |
| PTPRO | protein tyrosine phosphatase receptor type O |
| ROS | reactive oxygen species |
| RCC | renal cell carcinoma |
| STK16 | serine/threonine kinase 16 |
| SERS | surface-enhanced Raman spectroscopy |
| TLR | toll-like receptor |
| TGF-β | transforming growth factor β |
| TIMS | trapped ion mobility spectrometry |
| TNF | tumor necrosis factor |
References
- Ferlay, J.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.; Bray, F. Global Cancer Observatory: Cancer Today; International Agency for Research on Cancer: Lyon, France, 2020; Available online: https://gco.iarc.fr/today/online-analysis-multi-bars (accessed on 20 March 2023).
- Yadav, A.; Singh, S.; Sohi, H.; Dang, S. Advances in Delivery of Chemotherapeutic Agents for Cancer Treatment. AAPS PharmSciTech 2021, 23, 25. [Google Scholar] [CrossRef] [PubMed]
- Rosenblum, D.; Joshi, N.; Tao, W.; Karp, J.M.; Peer, D. Progress and Challenges towards Targeted Delivery of Cancer Therapeutics. Nat. Commun. 2018, 9, 1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Q.; Wilhelm, S.; Ding, D.; Syed, A.M.; Sindhwani, S.; Zhang, Y.; Chen, Y.Y.; MacMillan, P.; Chan, W.C.W. Quantifying the Ligand-Coated Nanoparticle Delivery to Cancer Cells in Solid Tumors. ACS Nano 2018, 12, 8423–8435. [Google Scholar] [CrossRef]
- Vargason, A.M.; Anselmo, A.C.; Mitragotri, S. The Evolution of Commercial Drug Delivery Technologies. Nat. Biomed. Eng. 2021, 5, 951–967. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, T.; Itaya, M.; Burdeos, G.C.; Nakagawa, K.; Miyazawa, T. A Critical Review of the Use of Surfactant-Coated Nanoparticles in Nanomedicine and Food Nanotechnology. Int. J. Nanomed. 2021, 16, 3937–3999. [Google Scholar] [CrossRef]
- Kibria, G.; Ramos, E.K.; Wan, Y.; Gius, D.R.; Liu, H. Exosomes as a Drug Delivery System in Cancer Therapy: Potential and Challenges. Mol. Pharm. 2018, 15, 3625–3633. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Z. The History and Advances in Cancer Immunotherapy: Understanding the Characteristics of Tumor-Infiltrating Immune Cells and Their Therapeutic Implications. Cell. Mol. Immunol. 2020, 17, 807–821. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Smith, W.; Hao, D.; He, B.; Kong, L. M1 and M2 Macrophage Polarization and Potentially Therapeutic Naturally Occurring Compounds. Int. Immunopharmacol. 2019, 70, 459–466. [Google Scholar] [CrossRef]
- Li, R.; Li, D.; Wang, H.; Chen, K.; Wang, S.; Xu, J.; Ji, P. Exosomes from Adipose-Derived Stem Cells Regulate M1/M2 Macrophage Phenotypic Polarization to Promote Bone Healing via MiR-451a/MIF. Stem Cell Res. Ther. 2022, 13, 149. [Google Scholar] [CrossRef]
- Zhang, B.; Yang, Y.; Yi, J.; Zhao, Z.; Ye, R. Hyperglycemia Modulates M1/M2 Macrophage Polarization via Reactive Oxygen Species Overproduction in Ligature-Induced Periodontitis. J. Periodontal Res. 2021, 56, 991–1005. [Google Scholar] [CrossRef]
- Liu, L.; Wu, Y.; Wang, B.; Jiang, Y.; Lin, L.; Li, X.; Yang, S. DA-DRD5 Signaling Controls Colitis by Regulating Colonic M1/M2 Macrophage Polarization. Cell Death Dis. 2021, 12, 500. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, W.; Wang, S.; Zhang, P.; Wang, Q.; Xiao, J.; Zhang, C.; Zheng, X.; Xu, X.; Xue, S.; et al. YAP Aggravates Inflammatory Bowel Disease by Regulating M1/M2 Macrophage Polarization and Gut Microbial Homeostasis. Cell Rep. 2019, 27, 1176–1189.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, Y.; Zhou, K.; Zhang, L.; Nan, S.; Fu, Z. Hydrolyzed Chicken Meat Extract Boosts the Immunoregulatory Effect by Regulating M1/M2 Macrophage Polarization. J. Funct. Foods 2022, 95, 105194. [Google Scholar] [CrossRef]
- Lu, H.; Wu, L.; Liu, L.; Ruan, Q.; Zhang, X.; Hong, W.; Wu, S.; Jin, G.; Bai, Y. Quercetin Ameliorates Kidney Injury and Fibrosis by Modulating M1/M2 Macrophage Polarization. Biochem. Pharmacol. 2018, 154, 203–212. [Google Scholar] [CrossRef]
- Cui, Y.; Lin, Y.; Meng, X.; Ma, J.; Deng, H.; Liu, X.; He, X.; Zhao, J. Cyanidin-3-Galactoside from Aronia Melanocarpa Ameliorates PM10 Induced Pulmonary Injury by Modulating M1/M2 Macrophage Polarization and NRF2/Sirt1 MAPK Signaling. J. Funct. Foods 2021, 78, 104363. [Google Scholar] [CrossRef]
- Zhuang, H.; Lv, Q.; Zhong, C.; Cui, Y.; He, L.; Zhang, C.; Yu, J. Tiliroside Ameliorates Ulcerative Colitis by Restoring the M1/M2 Macrophage Balance via the HIF-1α/Glycolysis Pathway. Front. Immunol. 2021, 12, 649463. [Google Scholar] [CrossRef]
- Bao, S.; Zou, Y.; Wang, B.; Li, Y.; Zhu, J.; Luo, Y.; Li, J. Ginsenoside Rg1 Improves Lipopolysaccharide-Induced Acute Lung Injury by Inhibiting Inflammatory Responses and Modulating Infiltration of M2 Macrophages. Int. Immunopharmacol. 2015, 28, 429–434. [Google Scholar] [CrossRef]
- Yang, P.; Ling, L.; Sun, W.; Yang, J.; Zhang, L.; Chang, G.; Guo, J.; Sun, J.; Sun, L.; Lu, D. Ginsenoside Rg1 Inhibits Apoptosis by Increasing Autophagy via the AMPK/MTOR Signaling in Serum Deprivation Macrophages. Acta Biochim. Biophys. Sin. 2018, 50, 144–155. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.J.; Jang, M.; Choi, J.; Chang, B.S.; Kim, D.Y.; Kim, S.-H.; Kwak, Y.-S.; Oh, S.; Lee, J.-H.; Chang, B.-J.; et al. Korean Red Ginseng and Ginsenoside-Rb1/-Rg1 Alleviate Experimental Autoimmune Encephalomyelitis by Suppressing Th1 and Th17 Cells and Upregulating Regulatory T Cells. Mol. Neurobiol. 2016, 53, 1977–2002. [Google Scholar] [CrossRef]
- Long, J.; Liu, X.-K.; Kang, Z.-P.; Wang, M.-X.; Zhao, H.-M.; Huang, J.-Q.; Xiao, Q.-P.; Liu, D.-Y.; Zhong, Y.-B. Ginsenoside Rg1 Ameliorated Experimental Colitis by Regulating the Balance of M1/M2 Macrophage Polarization and the Homeostasis of Intestinal Flora. Eur. J. Pharmacol. 2022, 917, 174742. [Google Scholar] [CrossRef]
- Wang, S.; Cao, M.; Xu, S.; Shi, J.; Mao, X.; Yao, X.; Liu, C. Luteolin Alters Macrophage Polarization to Inhibit Inflammation. Inflammation 2020, 43, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Abron, J.D.; Singh, N.P.; Price, R.L.; Nagarkatti, M.; Nagarkatti, P.S.; Singh, U.P. Genistein Induces Macrophage Polarization and Systemic Cytokine to Ameliorate Experimental Colitis. PLoS ONE 2018, 13, e0199631. [Google Scholar] [CrossRef] [PubMed]
- Wakade, C.; Giri, B.; Malik, A.; Khodadadi, H.; Morgan, J.C.; Chong, R.K.; Baban, B. Niacin Modulates Macrophage Polarization in Parkinson’s Disease. J. Neuroimmunol. 2018, 320, 76–79. [Google Scholar] [CrossRef]
- Abe, C.; Miyazawa, T.; Miyazawa, T. Current Use of Fenton Reaction in Drugs and Food. Molecules 2022, 27, 5451. [Google Scholar] [CrossRef] [PubMed]
- Behmoaras, J. The Versatile Biochemistry of Iron in Macrophage Effector Functions. FEBS J. 2021, 288, 6972–6989. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-Mediated Transfer of MRNAs and MicroRNAs Is a Novel Mechanism of Genetic Exchange between Cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, A.; Okada, R.; Nagao, K.; Kawamata, Y.; Hanyu, A.; Yoshimoto, S.; Takasugi, M.; Watanabe, S.; Kanemaki, M.T.; Obuse, C.; et al. Exosomes Maintain Cellular Homeostasis by Excreting Harmful DNA from Cells. Nat. Commun. 2017, 8, 15287. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Hagiwara, K.; Takeshita, F.; Ochiya, T. Competitive Interactions of Cancer Cells and Normal Cells via Secretory MicroRNAs*. J. Biol. Chem. 2012, 287, 1397–1405. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory Mechanisms and Intercellular Transfer of MicroRNAs in Living Cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [Green Version]
- Tan, A.; Rajadas, J.; Seifalian, A.M. Exosomes as Nano-Theranostic Delivery Platforms for Gene Therapy. Adv. Drug Deliv. Rev. 2013, 65, 357–367. [Google Scholar] [CrossRef]
- ExoCarta: Home—Exosome Database. Available online: http://exocarta.org/# (accessed on 20 March 2023).
- Bang, C.; Thum, T. Exosomes: New Players in Cell–Cell Communication. Int. J. Biochem. Cell Biol. 2012, 44, 2060–2064. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Q.; Zhang, X.; Huang, H.; Tang, S.; Chai, Y.; Xu, Z.; Li, M.; Chen, X.; Liu, J.; et al. Recent Advances in Exosome-Mediated Nucleic Acid Delivery for Cancer Therapy. J. Nanobiotechnol. 2022, 20, 279. [Google Scholar] [CrossRef] [PubMed]
- Yim, N.; Ryu, S.-W.; Choi, K.; Lee, K.R.; Lee, S.; Choi, H.; Kim, J.; Shaker, M.R.; Sun, W.; Park, J.-H.; et al. Exosome Engineering for Efficient Intracellular Delivery of Soluble Proteins Using Optically Reversible Protein–Protein Interaction Module. Nat. Commun. 2016, 7, 12277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.S.; Haney, M.J.; Zhao, Y.; Mahajan, V.; Deygen, I.; Klyachko, N.L.; Inskoe, E.; Piroyan, A.; Sokolsky, M.; Okolie, O.; et al. Development of Exosome-Encapsulated Paclitaxel to Overcome MDR in Cancer Cells. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 655–664. [Google Scholar] [CrossRef] [Green Version]
- Gomari, H.; Moghadam, M.F.; Soleimani, M. Targeted Cancer Therapy Using Engineered Exosome as a Natural Drug Delivery Vehicle. Onco. Targets. Ther. 2018, 11, 5753–5762. [Google Scholar] [CrossRef] [Green Version]
- Kooijmans, S.A.A.; Vader, P.; van Dommelen, S.M.; van Solinge, W.W.; Schiffelers, R.M. Exosome Mimetics: A Novel Class of Drug Delivery Systems. Int. J. Nanomed. 2012, 7, 1525–1541. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, D.; Moreira, J.N.; Rodrigues, L.R. New Advances in Exosome-Based Targeted Drug Delivery Systems. Crit. Rev. Oncol. Hematol. 2022, 172, 103628. [Google Scholar] [CrossRef]
- Wang, J.; Chen, D.; Ho, E.A. Challenges in the Development and Establishment of Exosome-Based Drug Delivery Systems. J. Control. Release 2021, 329, 894–906. [Google Scholar] [CrossRef]
- Sheridan, C. Exosome Cancer Diagnostic Reaches Market. Nat. Biotechnol. 2016, 34, 359–360. [Google Scholar] [CrossRef]
- Clinical Trials. Available online: https://clinicaltrials.gov/ct2/home (accessed on 20 March 2023).
- Chen, Y.; Ma, X.; Lou, C.; Zhou, C.; Zhao, X.; Li, N.; Tian, H.; Meng, X. PLA2G10 Incorporated in Exosomes Could Be Diagnostic and Prognostic Biomarker for Non-Small Cell Lung Cancer. Clin. Chim. Acta 2022, 530, 55–65. [Google Scholar] [CrossRef]
- Jin, X.; Ji, J.; Niu, D.; Yang, Y.; Tao, S.; Wan, L.; Xu, B.; Chen, S.; Wang, F.; Chen, M. Urine Exosomal AMACR Is a Novel Biomarker for Prostate Cancer Detection at Initial Biopsy. Front. Oncol. 2022, 12, 904315. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Xu, K.; Zheng, X.; Chen, T.; Wang, J.; Song, Y.; Shao, Y.; Zheng, S. Application of Exosomes as Liquid Biopsy in Clinical Diagnosis. Signal Transduct. Target. Ther. 2020, 5, 144. [Google Scholar] [CrossRef]
- Wang, X.; Huang, J.; Chen, W.; Li, G.; Li, Z.; Lei, J. The Updated Role of Exosomal Proteins in the Diagnosis, Prognosis, and Treatment of Cancer. Exp. Mol. Med. 2022, 54, 1390–1400. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Xu, X.; Li, B.; Situ, B.; Pan, W.; Hu, Y.; An, T.; Yao, S.; Zheng, L. Single-Exosome-Counting Immunoassays for Cancer Diagnostics. Nano Lett. 2018, 18, 4226–4232. [Google Scholar] [CrossRef]
- Buck, K.M.; Roberts, D.S.; Aballo, T.J.; Inman, D.R.; Jin, S.; Ponik, S.; Brown, K.A.; Ge, Y. One-Pot Exosome Proteomics Enabled by a Photocleavable Surfactant. Anal. Chem. 2022, 94, 7164–7168. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Y.; Li, P.; Zhang, Y.; Du, L.; Wang, Y.; Zhang, C.; Wang, C. Exosome Detection via Surface-Enhanced Raman Spectroscopy for Cancer Diagnosis. Acta Biomater. 2022, 144, 1–14. [Google Scholar] [CrossRef]
- Zhang, Y.; Mi, X.; Tan, X.; Xiang, R. Recent Progress on Liquid Biopsy Analysis Using Surface-Enhanced Raman Spectroscopy. Theranostics 2019, 9, 491–525. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Zhao, N.; Cui, K.; Chen, G.; Chen, Y.; Wu, W.; Li, Q.; Cui, Y.; Li, R.; Xiao, Z. Ultrasensitive Exosome Detection by Modularized SERS Labeling for Postoperative Recurrence Surveillance. ACS Sensors 2021, 6, 3234–3241. [Google Scholar] [CrossRef]
- Wang, P.; Liang, O.; Zhang, W.; Schroeder, T.; Xie, Y.-H. Ultra-Sensitive Graphene-Plasmonic Hybrid Platform for Label-Free Detection. Adv. Mater. 2013, 25, 4918–4924. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Dutta, S.; Liu, Z.; Yu, X.; Mesgarzadeh, N.; Ji, F.; Bitan, G.; Xie, Y.-H. A Label-Free Platform for Identification of Exosomes from Different Sources. ACS Sens. 2019, 4, 488–497. [Google Scholar] [CrossRef]
- Liu, Z.; Li, T.; Wang, Z.; Liu, J.; Huang, S.; Min, B.H.; An, J.Y.; Kim, K.M.; Kim, S.; Chen, Y.; et al. Gold Nanopyramid Arrays for Non-Invasive Surface-Enhanced Raman Spectroscopy-Based Gastric Cancer Detection via SEVs. ACS Appl. Nano Mater. 2022, 5, 12506–12517. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Oh, S.; Hong, S.; Kang, M.; Kang, D.; Ji, Y.; Choi, B.H.; Kang, K.-W.; Jeong, H.; Park, Y.; et al. Early-Stage Lung Cancer Diagnosis by Deep Learning-Based Spectroscopic Analysis of Circulating Exosomes. ACS Nano 2020, 14, 5435–5444. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Su, X.; Wen, Y.; Zheng, C.; Li, M. Artificial Intelligent Label-Free SERS Profiling of Serum Exosomes for Breast Cancer Diagnosis and Postoperative Assessment. Nano Lett. 2022, 22, 7910–7918. [Google Scholar] [CrossRef]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage Polarization: Tumor-Associated Macrophages as a Paradigm for Polarized M2 Mononuclear Phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Tu, D.; Dou, J.; Wang, M.; Zhuang, H.; Zhang, X. M2 Macrophages Contribute to Cell Proliferation and Migration of Breast Cancer. Cell Biol. Int. 2021, 45, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Ye, J.; Huang, C.; Yan, Y.; Li, J. M2 Macrophage-Derived IL6 Mediates Resistance of Breast Cancer Cells to Hedgehog Inhibition. Toxicol. Appl. Pharmacol. 2019, 364, 77–82. [Google Scholar] [CrossRef]
- Yang, X.; Cai, S.; Shu, Y.; Deng, X.; Zhang, Y.; He, N.; Wan, L.; Chen, X.; Qu, Y.; Yu, S. Exosomal MiR-487a Derived from M2 Macrophage Promotes the Progression of Gastric Cancer. Cell Cycle 2021, 20, 434–444. [Google Scholar] [CrossRef]
- Ma, Y.-S.; Wu, T.-M.; Ling, C.-C.; Yu, F.; Zhang, J.; Cao, P.-S.; Gu, L.-P.; Wang, H.-M.; Xu, H.; Li, L.; et al. M2 Macrophage-Derived Exosomal MicroRNA-155-5p Promotes the Immune Escape of Colon Cancer by Downregulating ZC3H12B. Mol. Ther. Oncolytics 2021, 20, 484–498. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Sun, L.; Xu, F.; Liu, L.; Hu, F.; Song, D.; Hou, Z.; Wu, W.; Luo, X.; Wang, J.; et al. M2 Macrophage-Derived Exosomes Promote Cell Migration and Invasion in Colon Cancer. Cancer Res. 2019, 79, 146–158. [Google Scholar] [CrossRef] [Green Version]
- Wei, K.; Ma, Z.; Yang, F.; Zhao, X.; Jiang, W.; Pan, C.; Li, Z.; Pan, X.; He, Z.; Xu, J.; et al. M2 Macrophage-Derived Exosomes Promote Lung Adenocarcinoma Progression by Delivering MiR-942. Cancer Lett. 2022, 526, 205–216. [Google Scholar] [CrossRef]
- Goswami, K.K.; Ghosh, T.; Ghosh, S.; Sarkar, M.; Bose, A.; Baral, R. Tumor Promoting Role of Anti-Tumor Macrophages in Tumor Microenvironment. Cell. Immunol. 2017, 316, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhou, R.; Wang, H.; Li, W.; Pan, M.; Yao, X.; Zhan, W.; Yang, S.; Xu, L.; Ding, Y.; et al. Gut Microbiota-Stimulated Cathepsin K Secretion Mediates TLR4-Dependent M2 Macrophage Polarization and Promotes Tumor Metastasis in Colorectal Cancer. Cell Death Differ. 2019, 26, 2447–2463. [Google Scholar] [CrossRef]
- Xiao, M.; Bian, Q.; Lao, Y.; Yi, J.; Sun, X.; Sun, X.; Yang, J. SENP3 Loss Promotes M2 Macrophage Polarization and Breast Cancer Progression. Mol. Oncol. 2022, 16, 1026–1044. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Murakami, R.; Hamanishi, J.; Tanigaki, K.; Hosoe, Y.; Mise, N.; Takamatsu, S.; Mise, Y.; Ukita, M.; Taki, M.; et al. B7-H3 Suppresses Antitumor Immunity via the CCL2–CCR2–M2 Macrophage Axis and Contributes to Ovarian Cancer Progression. Cancer Immunol. Res. 2022, 10, 56–69. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Young, M.-J.; Chang, H.-P.; Liu, C.-Y.; Lee, C.-C.; Tseng, Y.-L.; Wang, Y.-C.; Chang, W.-C.; Hung, J.-J. Estradiol-Mediated Inhibition of DNMT1 Decreases P53 Expression to Induce M2-Macrophage Polarization in Lung Cancer Progression. Oncogenesis 2022, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Kobatake, K.; Ikeda, K.; Nakata, Y.; Yamasaki, N.; Ueda, T.; Kanai, A.; Sentani, K.; Sera, Y.; Hayashi, T.; Koizumi, M.; et al. Kdm6a Deficiency Activates Inflammatory Pathways, Promotes M2 Macrophage Polarization, and Causes Bladder Cancer in Cooperation with P53 Dysfunction. Clin. Cancer Res. 2020, 26, 2065–2079. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Li, J.P.; Zhang, Y.; Nie, M.J.; Zhang, Y.H.; Liu, S.L.; Zou, X. Fstl3 Is a Prognostic Biomarker in Gastric Cancer and Is Correlated with M2 Macrophage Infiltration. Onco Targets Ther. 2021, 14, 4099–4117. [Google Scholar] [CrossRef]
- Zhao, L.; Zhou, Y.; Zhao, Y.; Li, Q.; Zhou, J.; Mao, Y. Long Non-Coding RNA TUSC8 Inhibits Breast Cancer Growth and Metastasis via MiR-190b-5p/MYLIP Axis. Aging 2020, 12, 2974–2991. [Google Scholar] [CrossRef]
- Zhou, L.; Li, J.; Liao, M.; Zhang, Q.; Yang, M. LncRNA MIR155HG Induces M2 Macrophage Polarization and Drug Resistance of Colorectal Cancer Cells by Regulating ANXA2. Cancer Immunol. Immunother. 2022, 71, 1075–1091. [Google Scholar] [CrossRef]
- Liu, S.-Q.; Zhou, Z.-Y.; Dong, X.; Guo, L.; Zhang, K.-J. LncRNA GNAS-AS1 Facilitates ER+ Breast Cancer Cells Progression by Promoting M2 Macrophage Polarization via Regulating MiR-433-3p/GATA3 Axis. Biosci. Rep. 2020, 40, BSR20200626. [Google Scholar] [CrossRef]
- Jiang, H.; Deng, W.; Zhu, K.; Zeng, Z.; Hu, B.; Zhou, Z.; Xie, A.; Zhang, C.; Fu, B.; Zhou, X.; et al. LINC00467 Promotes Prostate Cancer Progression via M2 Macrophage Polarization and the MiR-494-3p/STAT3 Axis. Front. Oncol. 2021, 11, 661431. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Shi, W.; Xu, Y.; Xu, C.; Zhao, T.; Geng, B.; Yang, J.; Pan, J.; Hu, S.; Zhang, C.; et al. Tumor-Derived Lactate Induces M2 Macrophage Polarization via the Activation of the ERK/STAT3 Signaling Pathway in Breast Cancer. Cell Cycle 2018, 17, 428–438. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, S. Lactic Acid Promotes Macrophage Polarization through MCT-HIF1α Signaling in Gastric Cancer. Exp. Cell Res. 2020, 388, 111846. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, L.; Shi, Y. Down-Regulated Paxillin Suppresses Cell Proliferation and Invasion by Inhibiting M2 Macrophage Polarization in Colon Cancer. Biol. Chem. 2018, 399, 1285–1295. [Google Scholar] [CrossRef] [PubMed]
- Na, Y.-R.; Yoon, Y.-N.; Son, D.-I.; Seok, S.-H. Cyclooxygenase-2 Inhibition Blocks M2 Macrophage Differentiation and Suppresses Metastasis in Murine Breast Cancer Model. PLoS ONE 2013, 8, e63451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Qin, T.; Zhao, Z.; Yang, G.; Sanches, J.G.P.; Zhang, Q.; Fan, S.; Cao, L.; Hu, X. Pentraxin-3 Inhibits Milky Spots Metastasis of Gastric Cancer by Inhibiting M2 Macrophage Polarization. J. Cancer 2021, 12, 4686–4697. [Google Scholar] [CrossRef]
- Ma, M.; Wang, X.; Liu, N.; Shan, F.; Feng, Y. Low-Dose Naltrexone Inhibits Colorectal Cancer Progression and Promotes Apoptosis by Increasing M1-Type Macrophages and Activating the Bax/Bcl-2/Caspase-3/PARP Pathway. Int. Immunopharmacol. 2020, 83, 106388. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Sun, J.; Yang, Y.; Liu, Y.; Lou, J.; Pan, H.; Yao, J.; Han, W. TMP195 Exerts Antitumor Effects on Colorectal Cancer by Promoting M1 Macrophages Polarization. Int. J. Biol. Sci. 2022, 18, 5653–5666. [Google Scholar] [CrossRef]
- Veerman, R.E.; Güçlüler Akpinar, G.; Eldh, M.; Gabrielsson, S. Immune Cell-Derived Extracellular Vesicles–Functions and Therapeutic Applications. Trends Mol. Med. 2019, 25, 382–394. [Google Scholar] [CrossRef]
- Li, Z.; Suo, B.; Long, G.; Gao, Y.; Song, J.; Zhang, M.; Feng, B.; Shang, C.; Wang, D. Exosomal MiRNA-16-5p Derived from M1 Macrophages Enhances T Cell-Dependent Immune Response by Regulating PD-L1 in Gastric Cancer. Front. Cell Dev. Biol. 2020, 8, 572689. [Google Scholar] [CrossRef]
- Wang, X.; Huang, R.; Lu, Z.; Wang, Z.; Chen, X.; Huang, D. Exosomes from M1-Polarized Macrophages Promote Apoptosis in Lung Adenocarcinoma via the MiR-181a-5p/ETS1/STK16 Axis. Cancer Sci. 2022, 113, 986–1001. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhou, L.; Shen, N.; Ning, X.; Wu, D.; Jiang, K.; Huang, X. M1 Macrophage-Derived Exosomes and Their Key Molecule LncRNA HOTTIP Suppress Head and Neck Squamous Cell Carcinoma Progression by Upregulating the TLR5/NF-ΚB Pathway. Cell Death Dis. 2022, 13, 183. [Google Scholar] [CrossRef] [PubMed]
- Choo, Y.W.; Kang, M.; Kim, H.Y.; Han, J.; Kang, S.; Lee, J.-R.; Jeong, G.-J.; Kwon, S.P.; Song, S.Y.; Go, S.; et al. M1 Macrophage-Derived Nanovesicles Potentiate the Anticancer Efficacy of Immune Checkpoint Inhibitors. ACS Nano 2018, 12, 8977–8993. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.; Jeon, M.; Jung, H.N.; Lee, W.; Hwang, J.-E.; Lee, J.S.; Choi, Y.; Im, H.-J. M1 Macrophage-Derived Exosome-Mimetic Nanovesicles with an Enhanced Cancer Targeting Ability. ACS Appl. Bio. Mater. 2022, 5, 2862–2869. [Google Scholar] [CrossRef]
- Tang, Z.; Tang, C.; Sun, C.; Ying, X.; Shen, R. M1 Macrophage-Derived Exosomes Synergistically Enhance the Anti- Bladder Cancer Effect of Gemcitabine. Aging 2022, 14, 7364–7377. [Google Scholar] [CrossRef]
- Zhao, Y.; Zheng, Y.; Zhu, Y.; Zhang, Y.; Zhu, H.; Liu, T. M1 Macrophage-Derived Exosomes Loaded with Gemcitabine and Deferasirox against Chemoresistant Pancreatic Cancer. Pharmaceutics 2021, 13, 1493. [Google Scholar] [CrossRef]
- Zhao, Y.; Zheng, Y.; Zhu, Y.; Li, H.; Zhu, H.; Liu, T. Docetaxel-Loaded M1 Macrophage-Derived Exosomes for a Safe and Efficient Chemoimmunotherapy of Breast Cancer. J. Nanobiotechnol. 2022, 20, 359. [Google Scholar] [CrossRef]
- Wang, P.; Wang, H.; Huang, Q.; Peng, C.; Yao, L.; Chen, H.; Qiu, Z.; Wu, Y.; Wang, L.; Chen, W. Exosomes from M1-Polarized Macrophages Enhance Paclitaxel Antitumor Activity by Activating Macrophages-Mediated Inflammation. Theranostics 2019, 9, 1714–1727. [Google Scholar] [CrossRef]
- Wang, F.; Li, B.; Wei, Y.; Zhao, Y.; Wang, L.; Zhang, P.; Yang, J.; He, W.; Chen, H.; Jiao, Z.; et al. Tumor-Derived Exosomes Induce PD1+ Macrophage Population in Human Gastric Cancer that Promotes Disease Progression. Oncogenesis 2018, 7, 41. [Google Scholar] [CrossRef] [Green Version]
- Xin, L.; Wu, Y.; Liu, C.; Zeng, F.; Wang, J.-L.; Wu, D.-Z.; Wu, J.; Yue, Z.-Q.; Gan, J.-H.; Lu, H.; et al. Exosome-Mediated Transfer of LncRNA HCG18 Promotes M2 Macrophage Polarization in Gastric Cancer. Mol. Immunol. 2021, 140, 196–205. [Google Scholar] [CrossRef]
- Qiu, S.; Xie, L.; Lu, C.; Gu, C.; Xia, Y.; Lv, J.; Xuan, Z.; Fang, L.; Yang, J.; Zhang, L.; et al. Gastric Cancer-Derived Exosomal MiR-519a-3p Promotes Liver Metastasis by Inducing Intrahepatic M2-like Macrophage-Mediated Angiogenesis. J. Exp. Clin. Cancer Res. 2022, 41, 296. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, K.; Zhi, Y.; Wu, Y.; Chen, B.; Bai, J.; Wang, X. Tumor-Derived Exosomal MiR-19b-3p Facilitates M2 Macrophage Polarization and Exosomal LINC00273 Secretion to Promote Lung Adenocarcinoma Metastasis via Hippo Pathway. Clin. Transl. Med. 2021, 11, e478. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Yu, G. Hypoxic Lung Cancer Cell-Derived Exosomal MiR-21 Mediates Macrophage M2 Polarization and Promotes Cancer Cell Proliferation through Targeting IRF1. World J. Surg. Oncol. 2022, 20, 241. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Liu, Y.; Li, C.; Xu, C.; Ding, C.; Chen, J.; Zhao, J. Tumor-Derived Exosomal CircFARSA Mediates M2 Macrophage Polarization via the PTEN/PI3K/AKT Pathway to Promote Non-Small Cell Lung Cancer Metastasis. Cancer Treat. Res. Commun. 2021, 28, 100412. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Zhang, L.; Chen, Y.; Yu, Z.; Zhao, Z. Cancer Cell-Derived Exosomal LINC00313 Induces M2 Macrophage Differentiation in Non-Small Cell Lung Cancer. Clin. Transl. Oncol. 2022, 24, 2395–2408. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zhou, W.; Rong, Y.; Kuang, T.; Xu, X.; Wu, W.; Wang, D.; Lou, W. Exosomal MiRNA-106b from Cancer-Associated Fibroblast Promotes Gemcitabine Resistance in Pancreatic Cancer. Exp. Cell Res. 2019, 383, 111543. [Google Scholar] [CrossRef]
- Zhao, M.; Zhuang, A.; Fang, Y. Cancer-Associated Fibroblast-Derived Exosomal MiRNA-320a Promotes Macrophage M2 Polarization In Vitro by Regulating PTEN/PI3Kγ Signaling in Pancreatic Cancer. J. Oncol. 2022, 2022, 9514697. [Google Scholar] [CrossRef]
- Shen, T.; Miao, S.; Zhou, Y.; Yi, X.; Xue, S.; Du, B.; Tang, C.; Qu, L.; Fu, D.; Jia, R.; et al. Exosomal AP000439.2 from Clear Cell Renal Cell Carcinoma Induces M2 Macrophage Polarization to Promote Tumor Progression through Activation of STAT3. Cell Commun. Signal 2022, 20, 152. [Google Scholar] [CrossRef]
- Baig, M.S.; Roy, A.; Rajpoot, S.; Liu, D.; Savai, R.; Banerjee, S.; Kawada, M.; Faisal, S.M.; Saluja, R.; Saqib, U.; et al. Tumor-Derived Exosomes in the Regulation of Macrophage Polarization. Inflamm. Res. 2020, 69, 435–451. [Google Scholar] [CrossRef]
- Xu, Z.; Chen, Y.; Ma, L.; Chen, Y.; Liu, J.; Guo, Y.; Yu, T.; Zhang, L.; Zhu, L.; Shu, Y. Role of Exosomal Non-Coding RNAs from Tumor Cells and Tumor-Associated Macrophages in the Tumor Microenvironment. Mol. Ther. 2022, 30, 3133–3154. [Google Scholar] [CrossRef]
- Gharavi, A.T.; Hanjani, N.A.; Movahed, E.; Doroudian, M. The Role of Macrophage Subtypes and Exosomes in Immunomodulation. Cell. Mol. Biol. Lett. 2022, 27, 83. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhang, S.; Jia, J.; Yang, J.; Song, Y.; Duan, H. Exosomal MiR-423–3p Inhibits Macrophage M2 Polarization to Suppress the Malignant Progression of Cervical Cancer. Pathol. Res. Pract. 2022, 235, 153882. [Google Scholar] [CrossRef] [PubMed]
- Tong, F.; Mao, X.; Zhang, S.; Xie, H.; Yan, B.; Wang, B.; Sun, J.; Wei, L. HPV + HNSCC-Derived Exosomal MiR-9 Induces Macrophage M1 Polarization and Increases Tumor Radiosensitivity. Cancer Lett. 2020, 478, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Moradi-Chaleshtori, M.; Bandehpour, M.; Heidari, N.; Mohammadi-Yeganeh, S.; Mahmoud Hashemi, S. Exosome-Mediated MiR-33 Transfer Induces M1 Polarization in Mouse Macrophages and Exerts Antitumor Effect in 4T1 Breast Cancer Cell Line. Int. Immunopharmacol. 2021, 90, 107198. [Google Scholar] [CrossRef]
- Moradi-Chaleshtori, M.; Shojaei, S.; Mohammadi-Yeganeh, S.; Hashemi, S.M. Transfer of MiRNA in Tumor-Derived Exosomes Suppresses Breast Tumor Cell Invasion and Migration by Inducing M1 Polarization in Macrophages. Life Sci. 2021, 282, 119800. [Google Scholar] [CrossRef]
- Moradi-Chaleshtori, M.; Bandehpour, M.; Soudi, S.; Mohammadi-Yeganeh, S.; Hashemi, S.M. In Vitro and in Vivo Evaluation of Anti-Tumoral Effect of M1 Phenotype Induction in Macrophages by MiR-130 and MiR-33 Containing Exosomes. Cancer Immunol. Immunother. 2021, 70, 1323–1339. [Google Scholar] [CrossRef]
- Moradi-Chaleshtori, M.; Koochaki, A.; Shojaei, S.; Paryan, M.; Safarzadeh, M.; Hashemi, S.M.; Mohammadi-Yeganeh, S. Overexpression of Pigment Epithelium-Derived Factor in Breast Cancer Cell-Derived Exosomes Induces M1 Polarization in Macrophages. Immunol. Lett. 2022, 248, 31–36. [Google Scholar] [CrossRef]
- Jang, J.-Y.; Lee, J.-K.; Jeon, Y.-K.; Kim, C.-W. Exosome Derived from Epigallocatechin Gallate Treated Breast Cancer Cells Suppresses Tumor Growth by Inhibiting Tumor-Associated Macrophage Infiltration and M2 Polarization. BMC Cancer 2013, 13, 421. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.; Xie, C.; Jiang, Y.; Li, K.; Lin, Y.; Pang, X.; Xiong, X.; Zheng, J.; Ke, X.; Chen, Y.; et al. Tumor-Derived Exosomal Protein Tyrosine Phosphatase Receptor Type O Polarizes Macrophage to Suppress Breast Tumor Cell Invasion and Migration. Front. Cell Dev. Biol. 2021, 9, 703537. [Google Scholar] [CrossRef]
- Lin, Y.; Xu, J.; Lan, H. Tumor-Associated Macrophages in Tumor Metastasis: Biological Roles and Clinical Therapeutic Applications. J. Hematol. Oncol. 2019, 12, 76. [Google Scholar] [CrossRef]
- Rezaie, J.; Feghhi, M.; Etemadi, T. A review on exosomes application in clinical trials: Perspective, questions, and challenges. Cell Commun. Signal. 2022, 20, 145. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yao, X.; Yue, J.; Fang, Y.; Cao, G.; Midgley, A.C.; Nishinari, K.; Yang, Y. Advances in Bioactivity of MicroRNAs of Plant-Derived Exosome-Like Nanoparticles and Milk-Derived Extracellular Vesicles. J. Agric. Food Chem. 2022, 70, 6285–6299. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yan, X.; Zhang, Y.; Yang, M.; Ma, Y.; Zhang, Y.; Xu, Q.; Tu, K.; Zhang, M. Oral Administration of Turmeric-Derived Exosome-like Nanovesicles with Anti-Inflammatory and pro-Resolving Bioactions for Murine Colitis Therapy. J. Nanobiotechnol. 2022, 20, 206. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, Q.; Yu, Q.; Xiao, J.; Zhao, H. Cigarette Smoke Extract-Treated Airway Epithelial Cells-Derived Exosomes Promote M1 Macrophage Polarization in Chronic Obstructive Pulmonary Disease. Int. Immunopharmacol. 2021, 96, 107700. [Google Scholar] [CrossRef] [PubMed]
- Sica, A.; Erreni, M.; Allavena, P.; Chiara, P. Macrophage polarization in pathology. Cell. Mol. Life Sci. 2015, 72, 4111–4126. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Molecules | Tested Cancer | Polarization Skewing | Feature | Ref. |
|---|---|---|---|---|
| Cathepsin K | colorectal cancer | M2 | promoted tumor metastasis | [65] |
| SENP3 | breast cancer | M2 | promoted cancer progression | [66] |
| B7-H3 | ovarian cancer | M2 | promoted cancer progression | [67] |
| DNMT1 | lung cancer | M2 | promoted cancer progression | [68] |
| Kdm6a | bladder cancer | M2 | caused bladder cancer | [69] |
| Fstl3 | gastric cancer | M2 | macrophage infiltration | [70] |
| LncRNA MIR155HG | colorectal cancer | M2 | drug resistance of cancer cells | [72] |
| LncRNA GNAS-AS1 | breast cancer | M2 | promoted cancer progression | [73] |
| LINC00467 | prostate cancer | M2 | promoted cancer progression | [74] |
| Lactic acid | breast cancer | M2 | – | [75] |
| gastric cancer | M2 | – | [76] | |
| Low-dose naltrexone | malignant tumors | M1 | anti-tumor effect | [80] |
| TMP195 | colorectal cancer | M1 | anti-tumor effect | [81] |
| miR-16-5p (M1 macrophage derivative) | gastric cancer | – | regulated T cells | [83] |
| miR-181a-5p (M1 macrophage derivative) | lung adenocarcinoma | – | regulated apoptosis | [84] |
| LncRNA HOTTIP (M1 macrophage derivative) | head and neck squamous cell carcinoma | – | anti-cancer effect | [85] |
| Types of Cancer | Cargo | Exosomes Derived from | Feature | Ref. |
|---|---|---|---|---|
| Cervical cancer | miR-423-3p | HeLa cell | attenuated cancer cell progression | [105] |
| HNSCC oncogenesis | miR-9 | HPV + HNSCC cell | enhanced radiosensitivity | [106] |
| Breast cancer | miR-33 | 4T1 breast cancer cells | inhibited the invasion and migration | [107] |
| miR-130 | 4T1 breast cancer cells | reduced proliferation, migration, and invasion | [108] | |
| miR-130 and miR-33 | MDA-MB-231 cells | lowered tumor volumes | [109] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abe, C.; Bhaswant, M.; Miyazawa, T.; Miyazawa, T. The Potential Use of Exosomes in Anti-Cancer Effect Induced by Polarized Macrophages. Pharmaceutics 2023, 15, 1024. https://doi.org/10.3390/pharmaceutics15031024
Abe C, Bhaswant M, Miyazawa T, Miyazawa T. The Potential Use of Exosomes in Anti-Cancer Effect Induced by Polarized Macrophages. Pharmaceutics. 2023; 15(3):1024. https://doi.org/10.3390/pharmaceutics15031024
Chicago/Turabian StyleAbe, Chizumi, Maharshi Bhaswant, Teruo Miyazawa, and Taiki Miyazawa. 2023. "The Potential Use of Exosomes in Anti-Cancer Effect Induced by Polarized Macrophages" Pharmaceutics 15, no. 3: 1024. https://doi.org/10.3390/pharmaceutics15031024
APA StyleAbe, C., Bhaswant, M., Miyazawa, T., & Miyazawa, T. (2023). The Potential Use of Exosomes in Anti-Cancer Effect Induced by Polarized Macrophages. Pharmaceutics, 15(3), 1024. https://doi.org/10.3390/pharmaceutics15031024