
Transcriptional Regulation of Liver-Type OATP1B3 (Lt-OATP1B3) and Cancer-Type OATP1B3 (Ct-OATP1B3) Studied in Hepatocyte-Derived and Colon Cancer-Derived Cell Lines



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. RNA Isolation and Quantitative RT-Polymerase Chain Reaction
2.4. MatInspector Analysis
2.5. Generation of the Reporter Gene Constructs and Conduction of the Luciferase Assays
2.6. Site-Directed Mutagenesis
2.7. Nuclear Protein Isolation
2.8. Electrophoretic Mobility Shift Assay
2.9. Statistical Analysis
3. Results
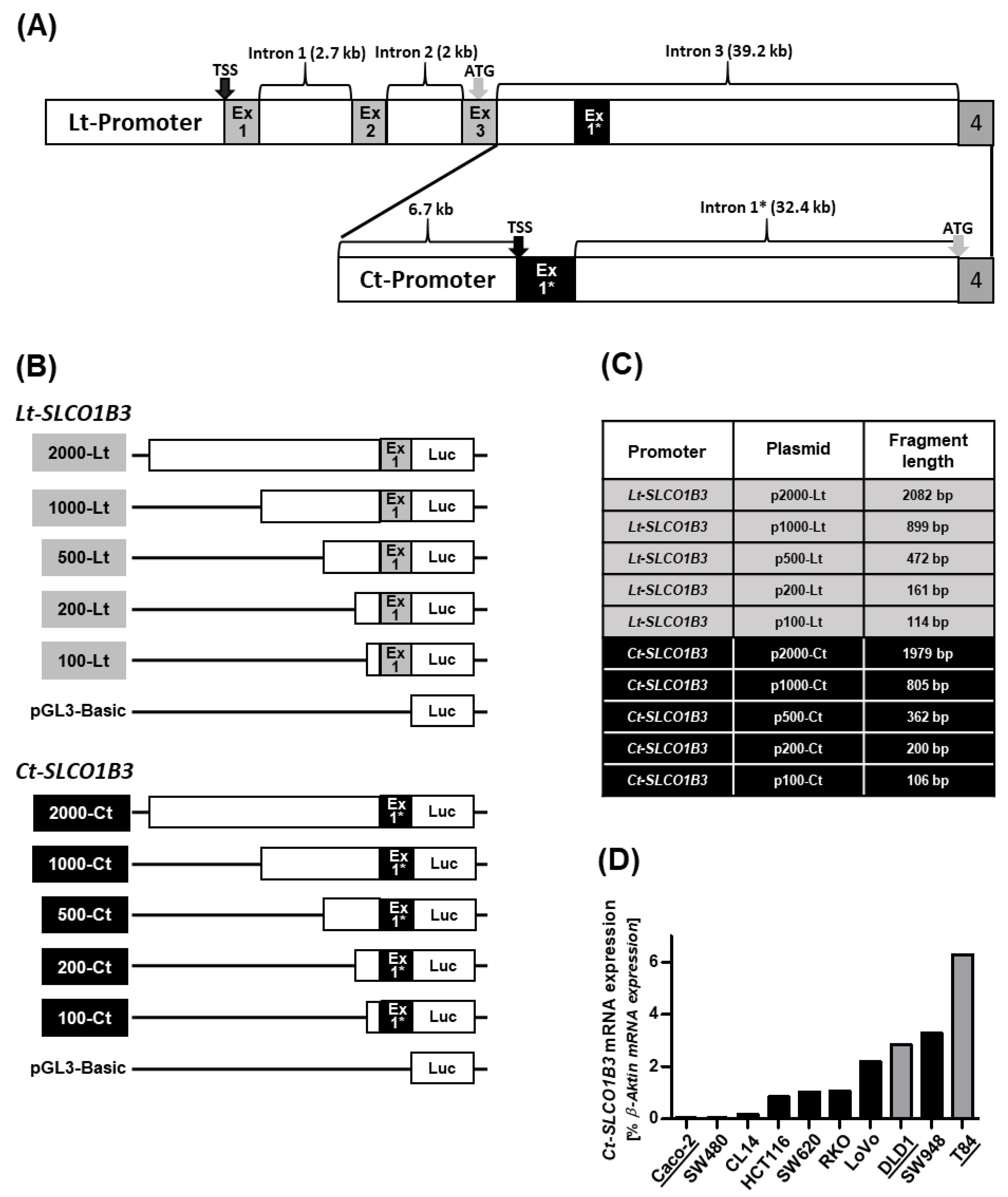
3.1. Cloning of the CT- and Lt-SLCO1B3 Reporter Gene Constructs
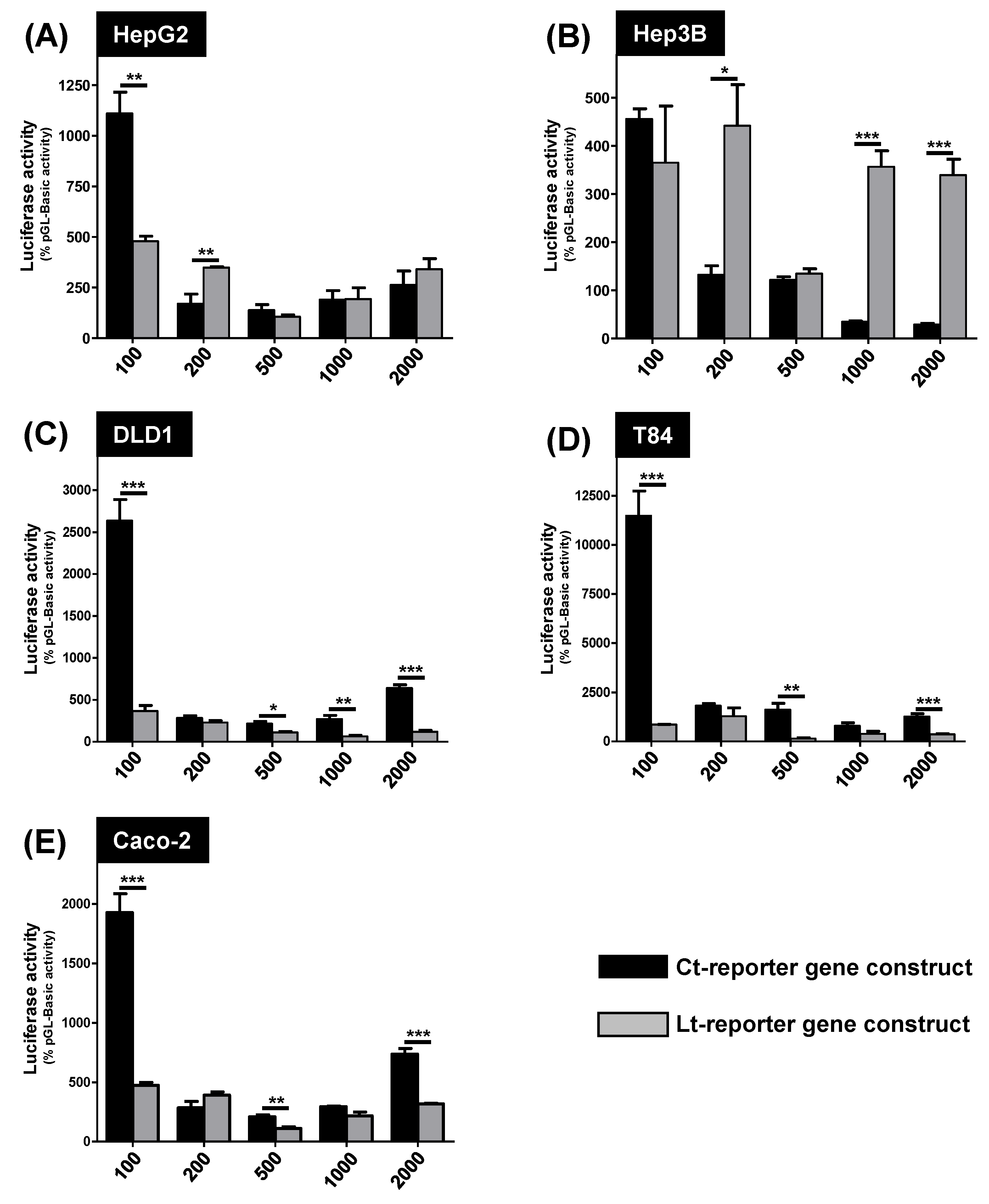
3.2. Measurement of the Luciferase Activity in the Different Cell Lines
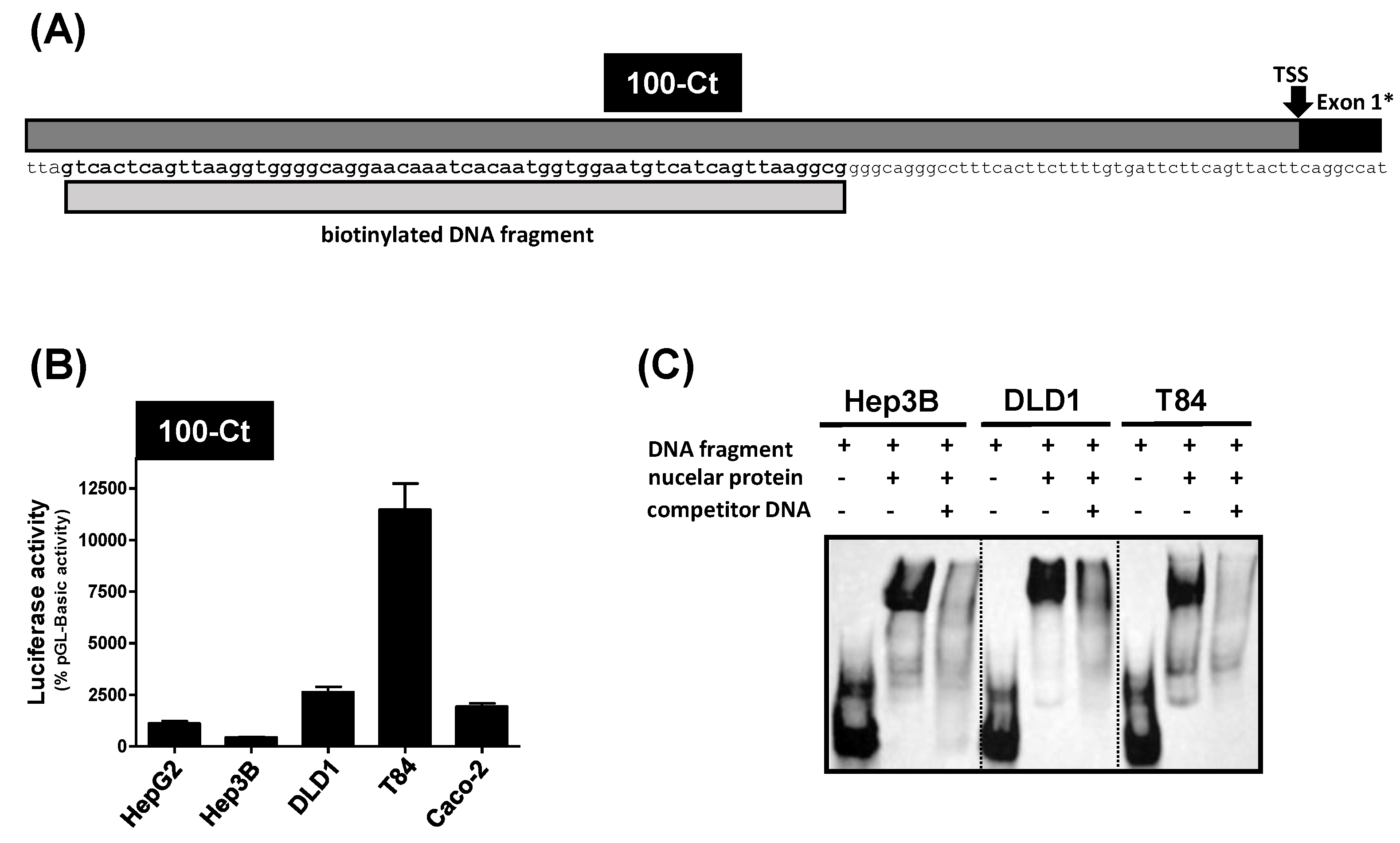
3.3. Analysis of the 100-Ct Promoter Construct
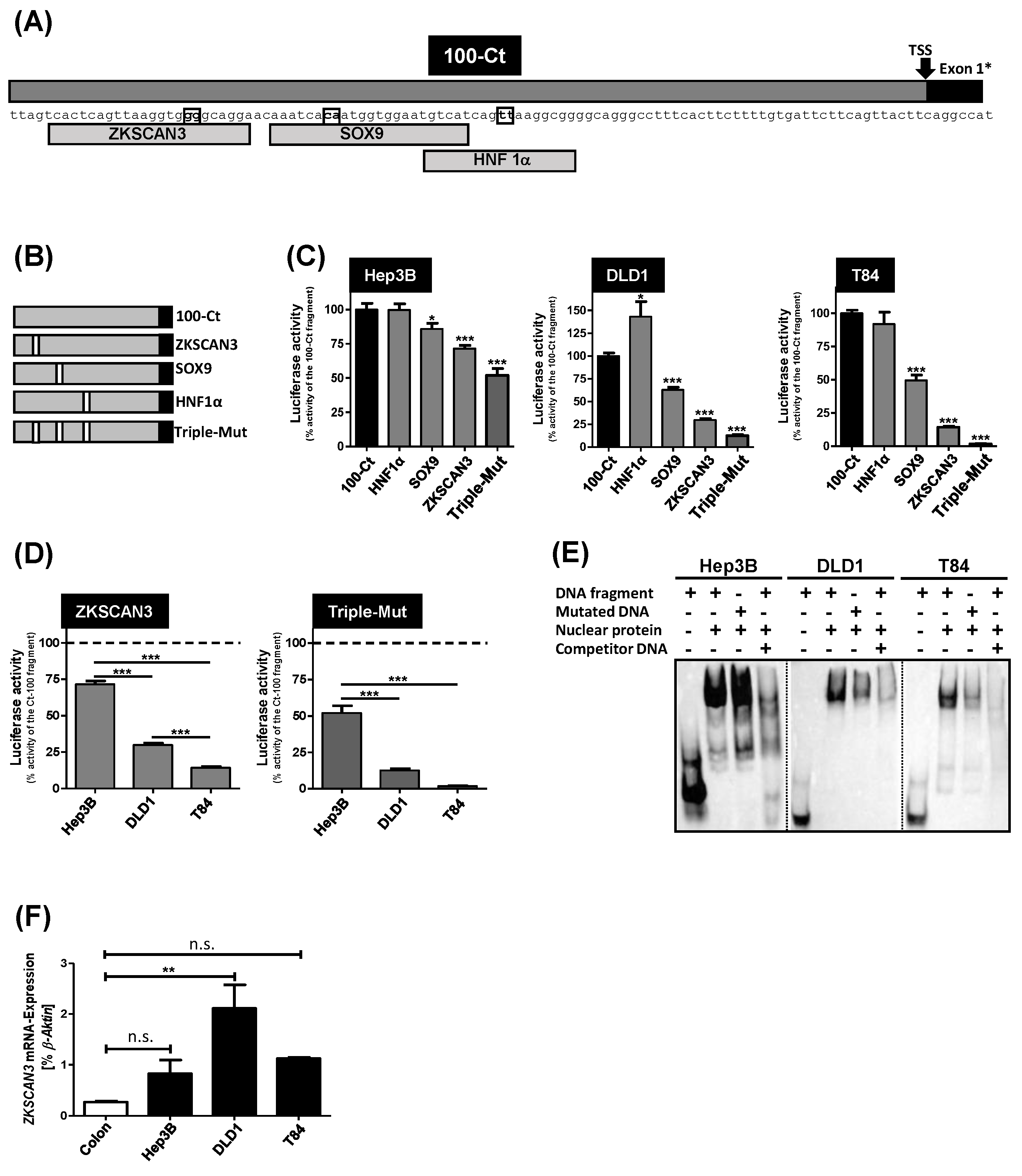
3.4. Mutational Analysis of Transcription Factor Binding Sites in the 100-Ct DNA Fragment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pizzagalli, M.D.; Bensimon, A.; Superti-Furga, G. A guide to plasma membrane solute carrier proteins. FEBS J. 2021, 288, 2784–2835. [Google Scholar] [CrossRef] [PubMed]
- Locher, K.P. Mechanistic diversity in ATP-binding cassette (ABC) transporters. Nat. Struct. Mol. Biol. 2016, 23, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Ahlin, G.; Hilgendorf, C.; Karlsson, J.; Szigyarto, C.A.; Uhlén, M.; Artursson, P. Endogenous gene and protein expression of drug-transporting proteins in cell lines routinely used in drug discovery programs. Drug Metab. Dispos. 2009, 37, 2275–2283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drozdzik, M.; Busch, D.; Lapczuk, J.; Müller, J.; Ostrowski, M.; Kurzawski, M.; Oswald, S. Protein abundance of clinically relevant drug transporters in the human liver and intestine: A comparative analysis in paired tissue specimens. Clin. Pharm. Ther. 2019, 105, 1204–1212. [Google Scholar] [CrossRef] [PubMed]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Ying, Y.; Tang, Z.; Liu, T.; Shi, F.; Li, H.; Guo, T.; Huang, S.; Lai, R. The emerging role of the SLCO1B3 protein in cancer resistance. Protein Pept. Lett. 2020, 27, 17–29. [Google Scholar] [CrossRef]
- Krchniakova, M.; Skoda, J.; Neradil, J.; Chlapek, P.; Veselska, R. Repurposing tyrosine kinase inhibitors to overcome multidrug resistance in cancer: A focus on transporters and lysosomal sequestration. Int. J. Mol. Sci. 2020, 21, 3157. [Google Scholar] [CrossRef]
- Yamagishi, T.; Sahni, S.; Sharp, D.M.; Arvind, A.; Jansson, P.J.; Richardson, D.R. P-glycoprotein mediates drug resistance via a novel mechanism involving lysosomal sequestration. J. Biol. Chem. 2013, 288, 31761–31771. [Google Scholar] [CrossRef] [Green Version]
- König, J.; Cui, Y.; Nies, A.T.; Keppler, D. Localization and genomic organization of a new hepatocellular organic anion transporting polypeptide. J. Biol. Chem. 2000, 275, 23161–23168. [Google Scholar] [CrossRef] [Green Version]
- König, J. Uptake transporters of the human OATP family: Molecular characteristics, substrates, their role in drug-drug interactions, and functional consequences of polymorphisms. In Handbook of Experimental Pharmacology 201, Drug Transporters, 26 November 2010 ed.; Fromm, M.F., Kim, R.B., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1–28. [Google Scholar] [CrossRef]
- Roth, M.; Obaidat, A.; Hagenbuch, B. OATPs, OATs and OCTs: The organic anion and cation transporters of the SLCO and SLC22A gene superfamilies. Br. J. Pharm. 2012, 165, 1260–1287. [Google Scholar] [CrossRef] [Green Version]
- Kullak-Ublick, G.A.; Ismair, M.G.; Stieger, B.; Landmann, L.; Huber, R.; Pizzagalli, F.; Fattinger, K.; Meier, P.J.; Hagenbuch, B. Organic anion-transporting polypeptide B (OATP-B) and its functional comparison with three other OATPs of human liver. Gastroenterology 2001, 120, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Seithel, A.; Eberl, S.; Singer, K.; Auge, D.; Heinkele, G.; Wolf, N.B.; Dörje, F.; Fromm, M.F.; König, J. The influence of macrolide antibiotics on the uptake of organic anions and drugs mediated by OATP1B1 and OATP1B3. Drug Metab. Dispos. 2007, 35, 779–786. [Google Scholar] [CrossRef] [Green Version]
- Abe, T.; Unno, M.; Onogawa, T.; Tokui, T.; Kondo, T.N.; Nakagomi, R.; Adachi, H.; Fujiwara, K.; Okabe, M.; Suzuki, T.; et al. LST-2, a human liver-specific organic anion transporter, determines methotrexate sensitivity in gastrointestinal cancers. Gastroenterology 2001, 120, 1689–1699. [Google Scholar] [CrossRef]
- Nagai, M.; Furihata, T.; Matsumoto, S.; Ishii, S.; Motohashi, S.; Yoshino, I.; Ugajin, M.; Miyajima, A.; Matsumoto, S.; Chiba, K. Identification of a new organic anion transporting polypeptide 1B3 mRNA isoform primarily expressed in human cancerous tissues and cells. Biochem. Biophys. Res. Commun. 2012, 418, 818–823. [Google Scholar] [CrossRef]
- Sun, Y.; Furihata, T.; Ishii, S.; Nagai, M.; Harada, M.; Shimozato, O.; Kamijo, T.; Motohashi, S.; Yoshino, I.; Kamiichi, A.; et al. Unique expression features of cancer-type organic anion transporting polypeptide 1B3 mRNA expression in human colon and lung cancers. Clin. Transl. Med. 2014, 3, 37. [Google Scholar] [CrossRef] [Green Version]
- Haberkorn, B.; Oswald, S.; Kehl, N.; Gessner, A.; Taudte, R.V.; Dobert, J.P.; Zunke, F.; Fromm, M.F.; König, J. Cancer-type organic anion transporting polypeptide 1B3 (Ct-OATP1B3) is localized in lysosomes and mediates resistance against kinase inhibitors. Mol. Pharm. 2022, 102, 248–258. [Google Scholar] [CrossRef]
- Lee, W.; Belkhiri, A.; Lockhart, A.C.; Merchant, N.; Glaeser, H.; Harris, E.I.; Washington, M.K.; Brunt, E.M.; Zaika, A.; Kim, R.B.; et al. Overexpression of OATP1B3 confers apoptotic resistance in colon cancer. Cancer Res. 2008, 68, 10315–10323. [Google Scholar] [CrossRef] [Green Version]
- Lockhart, A.C.; Harris, E.; Lafleur, B.J.; Merchant, N.B.; Washington, M.K.; Resnick, M.B.; Yeatman, T.J.; Lee, W. Organic anion transporting polypeptide 1B3 (OATP1B3) is overexpressed in colorectal tumors and is a predictor of clinical outcome. Clin. Exp. Gastroenterol. 2008, 1, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Muto, M.; Onogawa, T.; Suzuki, T.; Ishida, T.; Rikiyama, T.; Katayose, Y.; Ohuchi, N.; Sasano, H.; Abe, T.; Unno, M. Human liver-specific organic anion transporter-2 is a potent prognostic factor for human breast carcinoma. Cancer Sci. 2007, 98, 1570–1576. [Google Scholar] [CrossRef] [PubMed]
- Pressler, H.; Sissung, T.M.; Venzon, D.; Price, D.K.; Figg, W.D. Expression of OATP family members in hormone-related cancers: Potential markers of progression. PLoS ONE 2011, 6, e20372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.; Kim, K.; Thakkar, N.; Kim, D.; Lee, W. Role of hypoxia inducible factor-1α in the regulation of the cancer-specific variant of organic anion transporting polypeptide 1B3 (OATP1B3), in colon and pancreatic cancer. Biochem. Pharm. 2013, 86, 816–823. [Google Scholar] [CrossRef]
- Łanuszewska, J.; Cudak, A.; Rzeszowska-Wolny, J.; Widłak, P. Detection of damage-recognition proteins from human lymphocytes. Acta Biochim. Pol. 2000, 47, 443–450. [Google Scholar] [CrossRef]
- Ohtsuka, H.; Abe, T.; Onogawa, T.; Kondo, N.; Sato, T.; Oshio, H.; Mizutamari, H.; Mikkaichi, T.; Oikawa, M.; Rikiyama, T.; et al. Farnesoid X receptor, hepatocyte nuclear factors 1α and 3β are essential for transcriptional activation of the liver-specific organic anion transporter-2 gene. J. Gastroenterol. 2006, 41, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Hays, A.; Apte, U.; Hagenbuch, B. Organic anion transporting polypeptides expressed in pancreatic cancer may serve as potential diagnostic markers and therapeutic targets for early stage adenocarcinomas. Pharm. Res. 2013, 30, 2260–2269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorsen, K.; Schepeler, T.; Øster, B.; Rasmussen, M.H.; Vang, S.; Wang, K.; Hansen, K.Q.; Lamy, P.; Pedersen, J.S.; Eller, A.; et al. Tumor-specific usage of alternative transcription start sites in colorectal cancer identified by genome-wide exon array analysis. BMC Genom. 2011, 12, 505. [Google Scholar] [CrossRef] [Green Version]
- Carninci, P.; Sandelin, A.; Lenhard, B.; Katayama, S.; Shimokawa, K.; Ponjavic, J.; Semple, C.A.; Taylor, M.S.; Engström, P.G.; Frith, M.C.; et al. Genome-wide analysis of mammalian promoter architecture and evolution. Nat. Genet. 2006, 38, 626–635. [Google Scholar] [CrossRef]
- Yamashita, R.; Sathira, N.P.; Kanai, A.; Tanimoto, K.; Arauchi, T.; Tanaka, Y.; Hashimoto, S.; Sugano, S.; Nakai, K.; Suzuki, Y. Genome-wide characterization of transcriptional start sites in humans by integrative transcriptome analysis. Genome Res. 2011, 21, 775–789. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; König, J.; Nies, A.T.; Pfannschmidt, M.; Hergt, M.; Franke, W.W.; Alt, W.; Moll, R.; Keppler, D. Detection of the human organic anion transporters SLC21A6 (OATP2) and SLC21A8 (OATP8) in liver and hepatocellular carcinoma. Lab. Investig. 2003, 83, 527–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vavricka, S.R.; Jung, D.; Fried, M.; Grützner, U.; Meier, P.J.; Kullak-Ublick, G.A. The human organic anion transporting polypeptide 8 (SLCO1B3) gene is transcriptionally repressed by hepatocyte nuclear factor 3β in hepatocellular carcinoma. J. Hepatol. 2004, 40, 212–218. [Google Scholar] [CrossRef]
- Hilgendorf, C.; Ahlin, G.; Seithel, A.; Artursson, P.; Ungell, A.L.; Karlsson, J. Expression of thirty-six drug transporter genes in human intestine, liver, kidney, and organotypic cell lines. Drug Metab. Dispos. 2007, 35, 1333–1340. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, Y.; Wang, W.; Ninomiya, T.; Nagano, H.; Ohta, K.; Itoh, H. Liver enriched transcription factors and differentiation of hepatocellular carcinoma. Mol. Pathol. 1999, 52, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Ichihara, S.; Kikuchi, R.; Kusuhara, H.; Imai, S.; Maeda, K.; Sugiyama, Y. DNA methylation profiles of organic anion transporting polypeptide 1B3 in cancer cell lines. Pharm. Res. 2010, 27, 510–516. [Google Scholar] [CrossRef]
- Imai, S.; Kikuchi, R.; Tsuruya, Y.; Naoi, S.; Nishida, S.; Kusuhara, H.; Sugiyama, Y. Epigenetic regulation of organic anion transporting polypeptide 1B3 in cancer cell lines. Pharm. Res. 2013, 30, 2880–2890. [Google Scholar] [CrossRef] [PubMed]
- Tsuchihara, K.; Suzuki, Y.; Wakaguri, H.; Irie, T.; Tanimoto, K.; Hashimoto, S.; Matsushima, K.; Mizushima-Sugano, J.; Yamashita, R.; Nakai, K.; et al. Massive transcriptional start site analysis of human genes in hypoxia cells. Nucleic Acids Res. 2009, 37, 2249–2263. [Google Scholar] [CrossRef] [PubMed]
- Cartharius, K.; Frech, K.; Grote, K.; Klocke, B.; Haltmeier, M.; Klingenhoff, A.; Frisch, M.; Bayerlein, M.; Werner, T. MatInspector and beyond: Promoter analysis based on transcription factor binding sites. Bioinformatics 2005, 21, 2933–2942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Hamilton, S.R.; Sood, A.; Kuwai, T.; Ellis, L.; Sanguino, A.; Lopez-Berestein, G.; Boyd, D.D. The previously undescribed ZKSCAN3 (ZNF306) is a novel “driver” of colorectal cancer progression. Cancer Res. 2008, 68, 4321–4330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.W.; Roh, S.A.; Tak, K.H.; Koh, B.M.; Ha, Y.J.; Cho, D.H.; Kim, S.Y.; Kim, Y.S.; Kim, J.C. ZKSCAN3 facilitates liver metastasis of colorectal cancer associated with CEA-expressing tumor. Anticancer Res. 2016, 36, 2397–2406. [Google Scholar] [PubMed]
- Matheu, A.; Collado, M.; Wise, C.; Manterola, L.; Cekaite, L.; Tye, A.J.; Canamero, M.; Bujanda, L.; Schedl, A.; Cheah, K.S.; et al. Oncogenicity of the developmental transcription factor Sox9. Cancer Res. 2012, 72, 1301–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Z.; Deng, H.; Fang, Y.; Zhu, X.; Ye, G.T.; Yan, L.; Liu, H.; Li, G. Identification of the interplay between SOX9 and S100P in the metastasis and invasion of colon carcinoma. Oncotarget 2015, 6, 20672–20684. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| 2000-Ct.for | 5′-tggaggcaaggaattgcaact-3′ |
| 1000-Ct.for | 5′-gaaggaccaaggcaggcatc-3′ |
| 500-Ct.for | 5′-ttcatgtgtgtccatgtgaagag-3′ |
| 200-Ct.for | 5′-cagtcaaagggggttgttctct-3′ |
| 100-Ct.for | 5′-ttagtcactcagttaaggtgggg-3′ |
| Ct-Upstream.rev | 5′-taactgaccattcccttacctgc-3′ |
| 2000-Lt.for | 5′-gctgtcaagtagcagagacattgg-3′ |
| 1000-Lt.for | 5′-acaactgtcctgtcagtgataagg-3′ |
| 500-Lt.for | 5′-ccatgtgagatatccagtgtccatg-3′ |
| 200-Lt.for | 5′-gataggcttctggggtgaactcc-3′ |
| 100-Lt.for | 5′-ctgtttgcctaggacaatgacct-3′ |
| Lt-Upstream.rev | 5′-gcaactgcaacaagtccatcctt-3′ |
| Primer | Sequence |
|---|---|
| oHNF1α-Mut | 5′-ggtggaatgtcatcagccaaggcggggcaggg-3′ |
| oSOX9-Mut | 5′-aggtggggcaggaacaaatcaacatggtggaatgtcatcag-3′ |
| oZKSCAN3-mut | 5′-gttagtcactcagttaaggtgaagcaggaacaaatcacaatggt-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haberkorn, B.; Löwen, D.; Meier, L.; Fromm, M.F.; König, J. Transcriptional Regulation of Liver-Type OATP1B3 (Lt-OATP1B3) and Cancer-Type OATP1B3 (Ct-OATP1B3) Studied in Hepatocyte-Derived and Colon Cancer-Derived Cell Lines. Pharmaceutics 2023, 15, 738. https://doi.org/10.3390/pharmaceutics15030738
Haberkorn B, Löwen D, Meier L, Fromm MF, König J. Transcriptional Regulation of Liver-Type OATP1B3 (Lt-OATP1B3) and Cancer-Type OATP1B3 (Ct-OATP1B3) Studied in Hepatocyte-Derived and Colon Cancer-Derived Cell Lines. Pharmaceutics. 2023; 15(3):738. https://doi.org/10.3390/pharmaceutics15030738
Chicago/Turabian StyleHaberkorn, Bastian, Dennis Löwen, Lukas Meier, Martin F. Fromm, and Jörg König. 2023. "Transcriptional Regulation of Liver-Type OATP1B3 (Lt-OATP1B3) and Cancer-Type OATP1B3 (Ct-OATP1B3) Studied in Hepatocyte-Derived and Colon Cancer-Derived Cell Lines" Pharmaceutics 15, no. 3: 738. https://doi.org/10.3390/pharmaceutics15030738
APA StyleHaberkorn, B., Löwen, D., Meier, L., Fromm, M. F., & König, J. (2023). Transcriptional Regulation of Liver-Type OATP1B3 (Lt-OATP1B3) and Cancer-Type OATP1B3 (Ct-OATP1B3) Studied in Hepatocyte-Derived and Colon Cancer-Derived Cell Lines. Pharmaceutics, 15(3), 738. https://doi.org/10.3390/pharmaceutics15030738