
Biomimetic Silica Particles with Self-Loading BMP-2 Knuckle Epitope Peptide and Its Delivery for Bone Regeneration


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Silica Deposition
2.3. Silica Quantification
2.4. Co-Precipitation of P4 with Silica, Yielding P4-Encapsylated Silica Particle (P4@Si) by P4-Catalyzed Silicification and Measurement of Entrapment and Loading Efficiency
2.5. Determination of Release Kinetics
2.6. Determination of Silica Release from P4@Si
2.7. MC3T3 E1 Cell Culture
2.8. Flow Cytometry Analysis of Cellular Uptake of Free P4 and P4@Si
2.9. STED Microscopy and Confocal Images of MC3T3 E1 Cells
2.10. Preparation of HA Nanocomposite Powders (Scheme 1B)
2.11. Cell Proliferation
2.12. Osteogenic Gene Expression
2.13. Calcium Deposition and Quantification
2.14. SEM and EDX
2.15. Statistical Analysis
3. Results
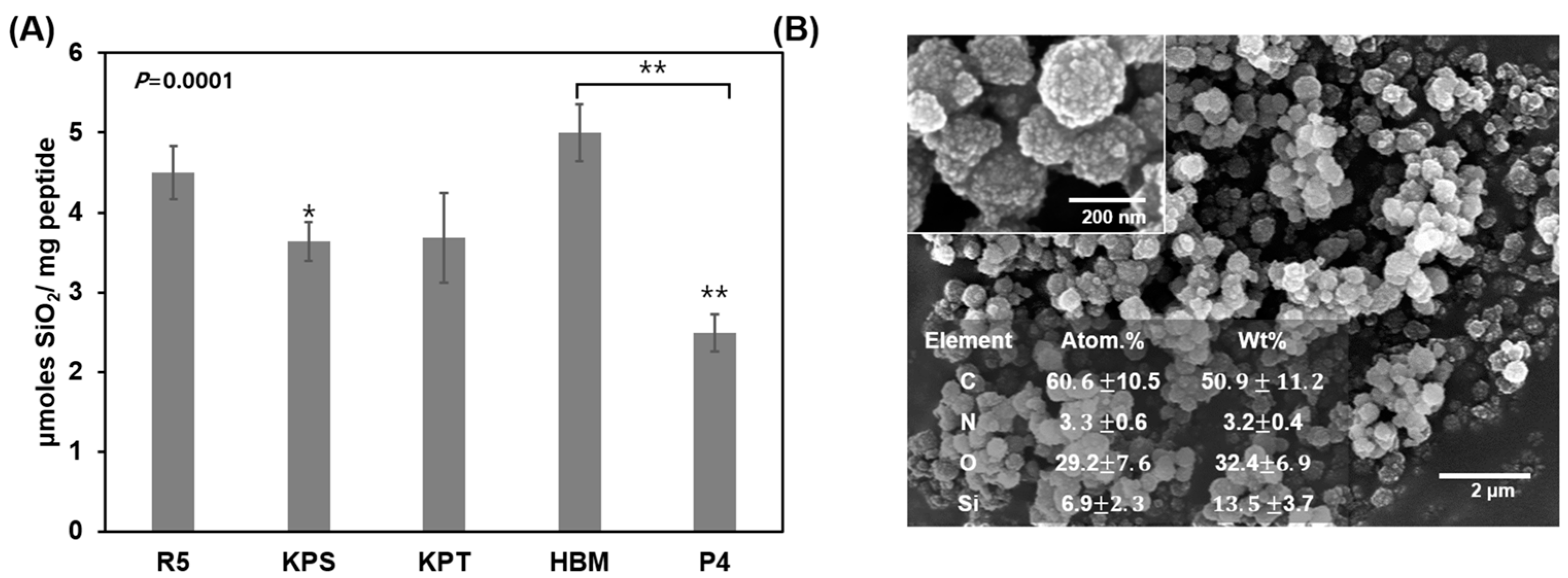
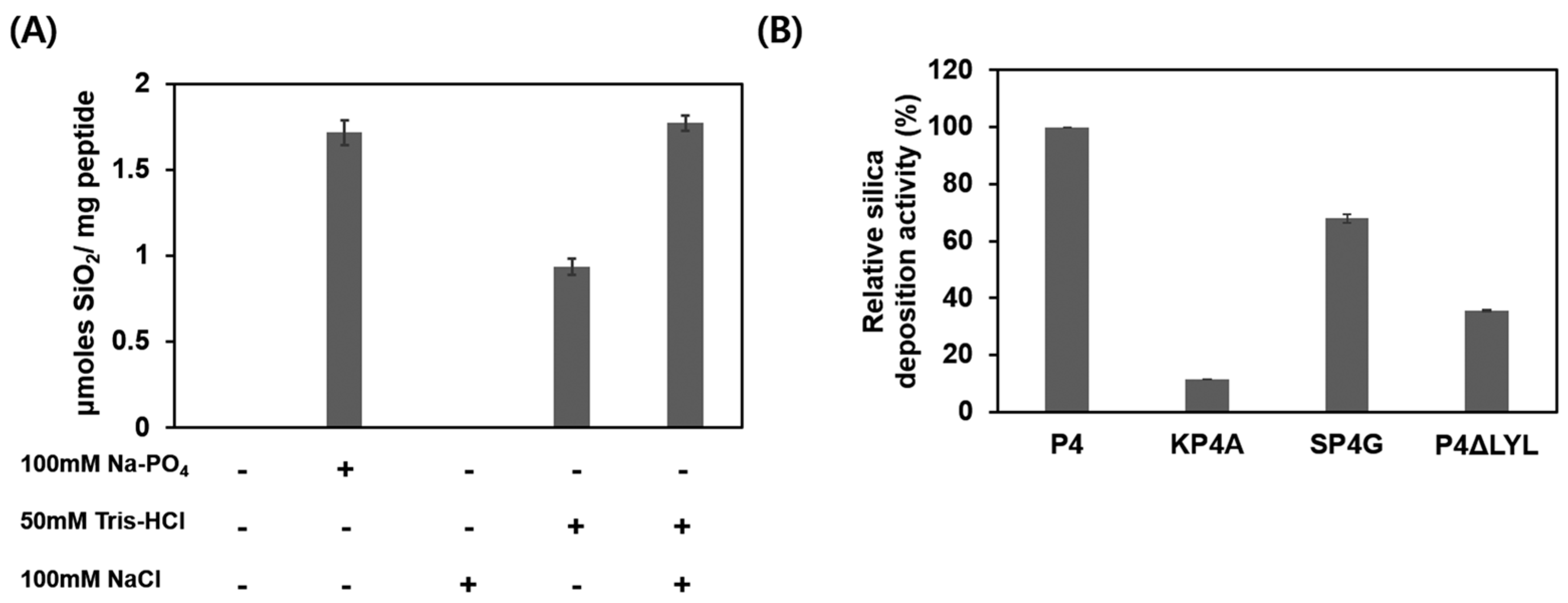
3.1. Silica Deposition Ability of Osteoinductive BMP-2 Knuckle Epitope Peptide (P4)
3.2. Characterization of P4-Mediated Silica Deposition
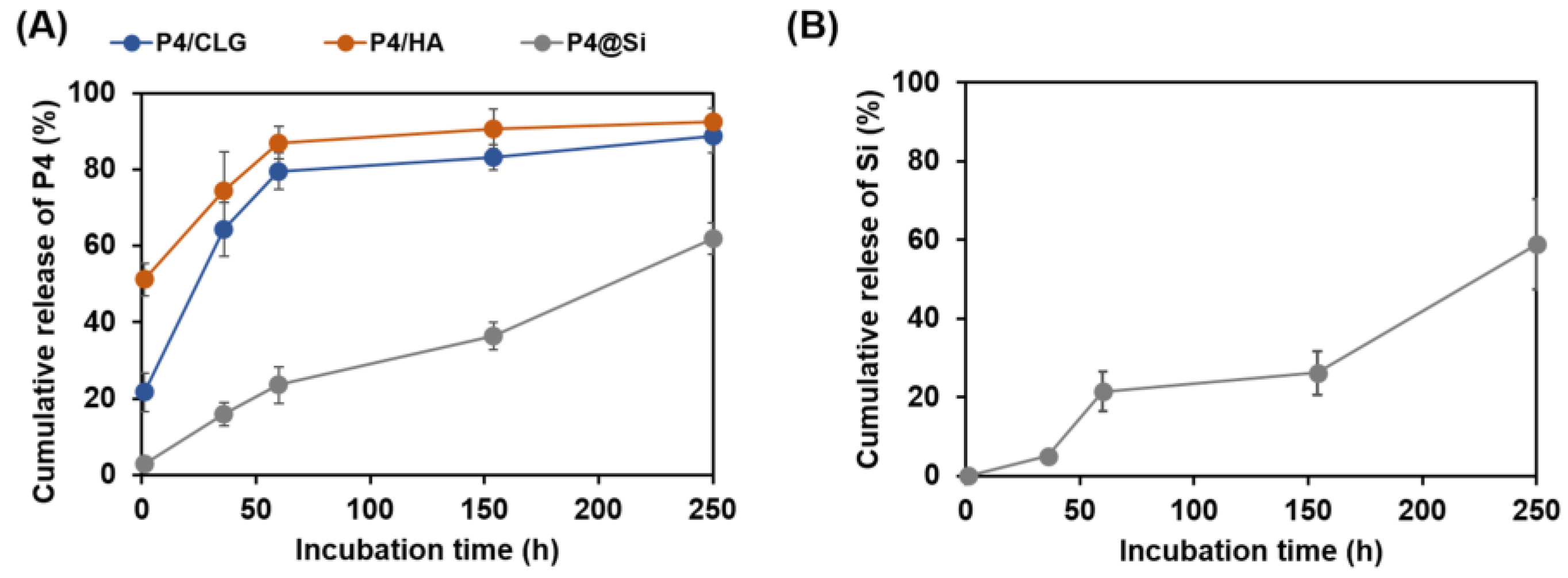
3.3. Peptide Release from Self-Entrapped Silica Particles
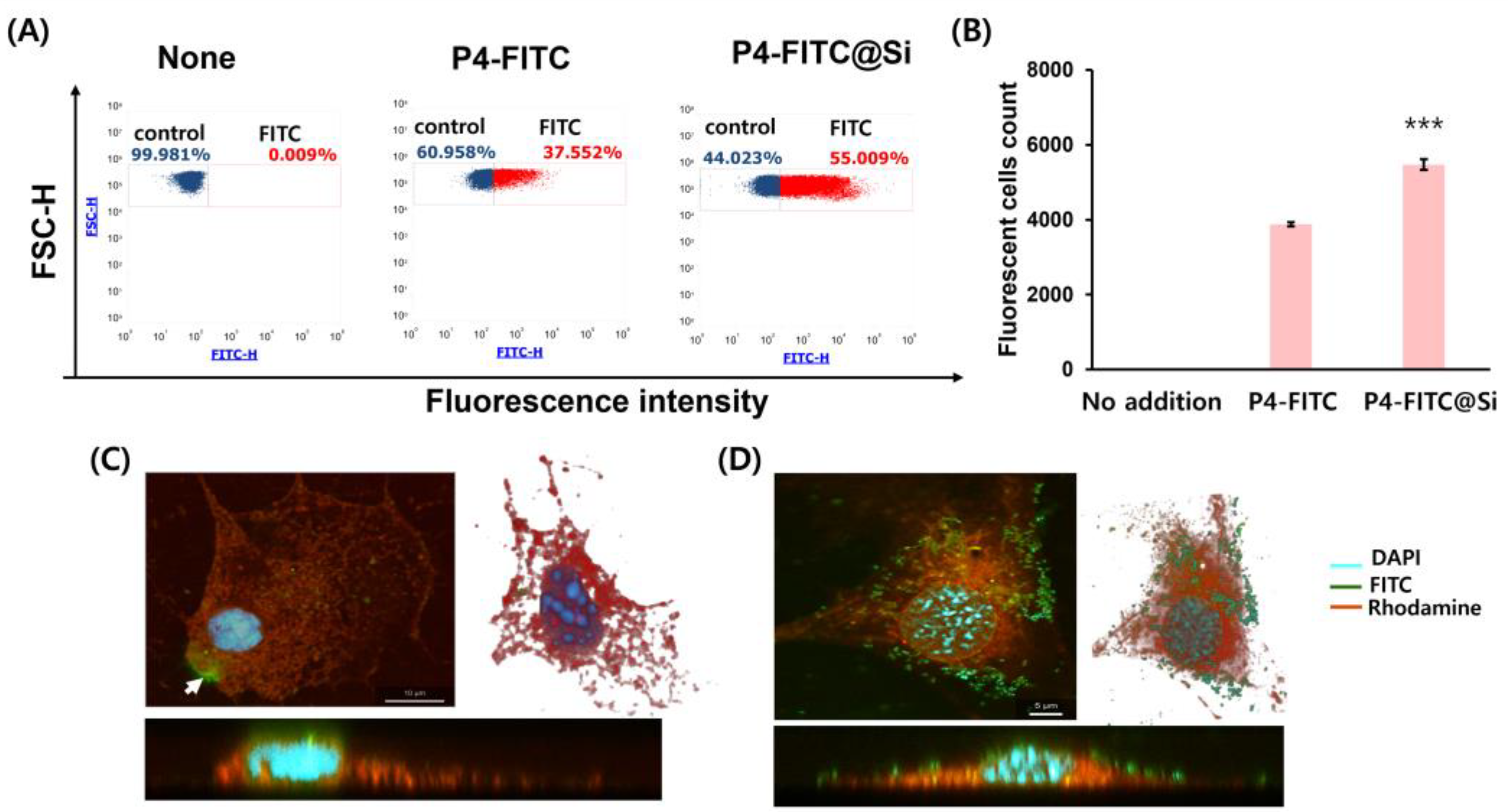
3.4. Enhancement of Cellular Uptake of P4 through Self-Entrapped Silica Carrier
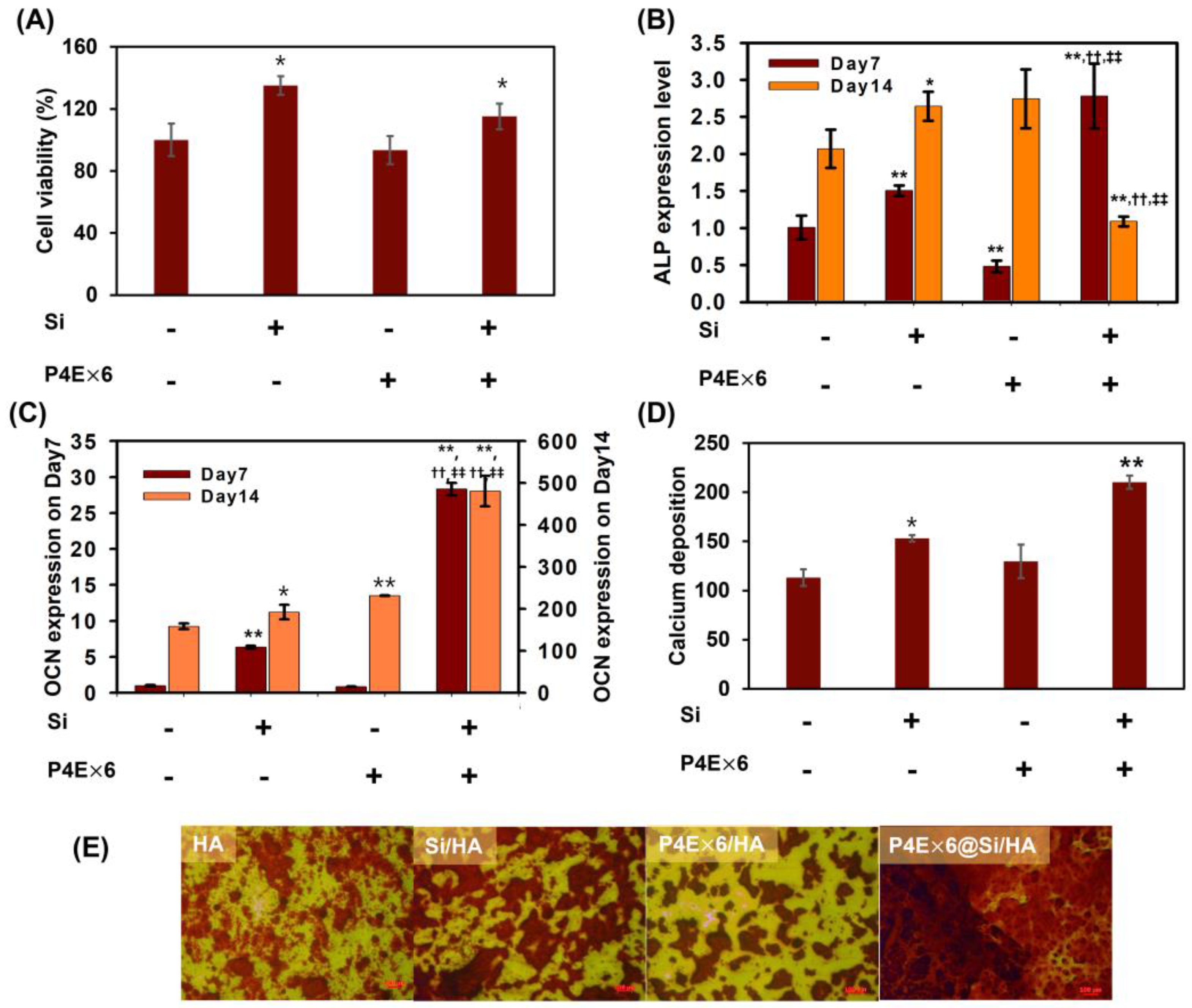
3.5. Synergistic Effect of the Combination of P4 with Silica on Osteogenesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- James, A.W.; LaChaud, G.; Shen, J.; Asatrian, G.; Nguyen, V.; Zhang, X.; Ting, K.; Soo, C. A Review of the Clinical Side Effects of Bone Morphogenetic Protein-2. Tissue Eng. Part B Rev. 2016, 22, 284–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Liu, Y.; Wang, A.; Zhu, Z.; Li, Y.; Zhu, C.; Che, Z.; Liu, T.; Liu, H.; Huang, L. Application of BMP in Bone Tissue Engineering. Front. Bioeng. Biotechnol. 2022, 10, 810880. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Coventry, B.; Goreshnik, I.; Huang, B.; Sheffler, W.; Park, J.S.; Jude, K.M.; Markovic, I.; Kadam, R.U.; Verschueren, K.H.G.; et al. Design of protein-binding proteins from the target structure alone. Nature 2022, 605, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Zhou, Q.; He, J.; Jiang, Z.; Peng, C.; Tong, R.; Shi, J. Recent advances in the development of protein-protein interactions modulators: Mechanisms and clinical trials. Signal Transduct. Target Ther. 2020, 5, 213. [Google Scholar] [CrossRef]
- Pelay-Gimeno, M.; Glas, A.; Koch, O.; Grossmann, T.N. Structure-Based Design of Inhibitors of Protein-Protein Interactions: Mimicking Peptide Binding Epitopes. Angew. Chem. Int. Ed. Engl. 2015, 54, 8896–8927. [Google Scholar] [CrossRef]
- Aoki, K.; Alles, N.; Soysa, N.; Ohya, K. Peptide-based delivery to bone. Adv. Drug Deliv. Rev. 2012, 64, 1220–1238. [Google Scholar] [CrossRef]
- Saito, A.; Suzuki, Y.; Ogata, S.; Ohtsuki, C.; Tanihara, M. Activation of osteo-progenitor cells by a novel synthetic peptide derived from the bone morphogenetic protein-2 knuckle epitope. Biochim. Biophys. Acta 2003, 1651, 60–67. [Google Scholar] [CrossRef]
- Zhang, A.; Chen, Z.; Yu, X.; Zhang, L.; Song, Q. Rational Derivation of Osteogenic Peptides from Bone Morphogenetic Protein-2 Knuckle Epitope by Integrating in silico Analysis and in vitro Assay. Int. J. Pept. Res. Ther. 2020, 27, 25–35. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, J.; Shan, H.; Yin, F.; Zhong, B.; Zhang, C.; Yu, X. Machine learning-guided evolution of BMP-2 knuckle Epitope-Derived osteogenic peptides to target BMP receptor II. J. Drug Target. 2020, 28, 802–810. [Google Scholar] [CrossRef]
- Miyazono, K.; Kamiya, Y.; Morikawa, M. Bone morphogenetic protein receptors and signal transduction. J. Biochem. 2010, 147, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Deng, C.; Li, Y.P. TGF-beta and BMP signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, Y.L.; Lin, L.D.; Chang, H.H.; Wang, T.M. Preliminary evaluation of BMP-2-derived peptide in repairing a peri-implant critical size defect: A canine model. J. Formos. Med. Assoc. 2021, 120, 1212–1220. [Google Scholar] [CrossRef] [PubMed]
- Moeinzadeh, S.; Barati, D.; Sarvestani, S.K.; Karimi, T.; Jabbari, E. Experimental and computational investigation of the effect of hydrophobicity on aggregation and osteoinductive potential of BMP-2-derived peptide in a hydrogel matrix. Tissue Eng. Part A 2015, 21, 134–146. [Google Scholar] [CrossRef] [Green Version]
- Bruno, B.J.; Miller, G.D.; Lim, C.S. Basics and recent advances in peptide and protein drug delivery. Ther. Deliv. 2013, 4, 1443–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culpepper, B.K.; Bonvallet, P.P.; Reddy, M.S.; Ponnazhagan, S.; Bellis, S.L. Polyglutamate directed coupling of bioactive peptides for the delivery of osteoinductive signals on allograft bone. Biomaterials 2013, 34, 1506–1513. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Yu, B.; Huang, Q.; Liu, R.; Feng, Q.; Cai, Q.; Mi, S. In vitro BMP-2 peptide release from thiolated chitosan based hydrogel. Int. J. Biol. Macromol. 2016, 93, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, X.; Liu, R.; Gong, Y.; Wang, M.; Huang, Q.; Feng, Q.; Yu, B. Zero-order controlled release of BMP2-derived peptide P24 from the chitosan scaffold by chemical grafting modification technique for promotion of osteogenesis in vitro and enhancement of bone repair in vivo. Theranostics 2017, 7, 1072–1087. [Google Scholar] [CrossRef]
- Luo, J.; Zhang, H.; Zhu, J.; Cui, X.; Gao, J.; Wang, X.; Xiong, J. 3-D mineralized silk fibroin/polycaprolactone composite scaffold modified with polyglutamate conjugated with BMP-2 peptide for bone tissue engineering. Colloids Surf. B Biointerfaces 2018, 163, 369–378. [Google Scholar] [CrossRef]
- Kim, M.J.; Lee, B.; Yang, K.; Park, J.; Jeon, S.; Um, S.H.; Kim, D.I.; Im, S.G.; Cho, S.W. BMP-2 peptide-functionalized nanopatterned substrates for enhanced osteogenic differentiation of human mesenchymal stem cells. Biomaterials 2013, 34, 7236–7246. [Google Scholar] [CrossRef] [PubMed]
- Bain, J.L.; Bonvallet, P.P.; Abou-Arraj, R.V.; Schupbach, P.; Reddy, M.S.; Bellis, S.L. Enhancement of the Regenerative Potential of Anorganic Bovine Bone Graft Utilizing a Polyglutamate-Modified BMP2 Peptide with Improved Binding to Calcium-Containing Materials. Tissue Eng. Part A 2015, 21, 2426–2436. [Google Scholar] [CrossRef] [Green Version]
- Boda, S.K.; Almoshari, Y.; Wang, H.; Wang, X.; Reinhardt, R.A.; Duan, B.; Wang, D.; Xie, J. Mineralized nanofiber segments coupled with calcium-binding BMP-2 peptides for alveolar bone regeneration. Acta Biomater. 2019, 85, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Feng, W.; Qiu, K.; Chen, L.; Wang, W.; Nie, W.; Mo, X.; He, C. BMP-2 Derived Peptide and Dexamethasone Incorporated Mesoporous Silica Nanoparticles for Enhanced Osteogenic Differentiation of Bone Mesenchymal Stem Cells. ACS Appl. Mater. Interfaces 2015, 7, 15777–15789. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Qu, Y.; Cui, W.; Yang, L.; Ji, Y.; Yu, W.; Navinduth, R.; Shao, Z.; Yang, H.; Guo, X. Evaluation of osteogenic inductivity of a novel BMP2-mimicking peptide P28 and P28-containing bone composite. J. Biomed. Mater. Res A 2018, 106, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Perikamana, S.K.M.; Ahmad, T.; Lee, M.S.; Yang, H.S.; Kim, D.G.; Kim, K.; Kwon, B.; Shin, H. Controlled Retention of BMP-2-Derived Peptide on Nanofibers Based on Mussel-Inspired Adhesion for Bone Formation. Tissue Eng. Part A 2017, 23, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Guo, X.; Zheng, Q.; Wu, Y.; Cui, F.; Wu, B. Improving osteogenesis of three-dimensional porous scaffold based on mineralized recombinant human-like collagen via mussel-inspired polydopamine and effective immobilization of BMP-2-derived peptide. Colloids Surf. B Biointerfaces 2017, 152, 124–132. [Google Scholar] [CrossRef]
- Gudivada, V.N.; Huang, C.J.; Luo, Y.H.; Dong, G.C. A Cyclic BMP-2 Peptide Upregulates BMP-2 Protein-Induced Cell Signaling in Myogenic Cells. Polymers 2021, 13, 2549. [Google Scholar] [CrossRef]
- Mumcuoglu, D.; Siverino, C.; Tabisz, B.; Kluijtmans, B.; Nickel, J. How to use BMP-2 for clinical applications? A review on pros and cons of existing delivery strategies. J. Transl. Sci. 2017, 3, 1–11. [Google Scholar] [CrossRef]
- Singh, P.; Srivastava, S.; Singh, S.K. Nanosilica: Recent Progress in Synthesis, Functionalization, Biocompatibility, and Biomedical Applications. ACS Biomater. Sci. Eng. 2019, 5, 4882–4898. [Google Scholar] [CrossRef]
- Hudson, S.P.; Padera, R.F.; Langer, R.; Kohane, D.S. The biocompatibility of mesoporous silicates. Biomaterials 2008, 29, 4045–4055. [Google Scholar] [CrossRef] [Green Version]
- Lechner, C.C.; Becker, C.F. Modified silaffin R5 peptides enable encapsulation and release of cargo molecules from biomimetic silica particles. Bioorg. Med. Chem. 2013, 21, 3533–3541. [Google Scholar] [CrossRef]
- Ki, M.R.; Nguyen, T.K.M.; Jun, H.S.; Pack, S.P. Biosilica-enveloped ferritin cage for more efficient drug deliveries. Process. Biochem. 2018, 68, 182–189. [Google Scholar] [CrossRef]
- Ki, M.R.; Kim, J.K.; Kim, S.H.; Nguyen, T.K.M.; Kim, K.H.; Pack, S.P. Compartment-restricted and rate-controlled dual drug delivery system using a biosilica-enveloped ferritin cage. J. Ind. Eng. Chem. 2020, 81, 367–374. [Google Scholar] [CrossRef]
- Nguyen, T.K.M.; Ki, M.R.; Lee, C.S.; Pack, S.P. Nanosized and tunable design of biosilica particles using novel sili-ca-forming peptide-modified chimeric ferritin templates. J. Ind. Eng. Chem. 2019, 73, 198–204. [Google Scholar] [CrossRef]
- Abdelhamid, M.A.A.; Pack, S.P. Biomimetic and bioinspired silicifications: Recent advances for biomaterial design and applications. Acta Biomater. 2021, 120, 38–56. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, M.A.A.; Ki, M.R.; Abd El-Hafeez, A.A.; Son, R.G.; Pack, S.P. Tailored Functionalized Protein Nanocarriers for Cancer Therapy: Recent Developments and Prospects. Pharmaceutics 2023, 15, 168. [Google Scholar] [CrossRef]
- Ki, M.-R.; Kim, S.H.; Nguyen, T.K.M.; Son, R.G.; Jun, S.H.; Pack, S.P. BMP2-Mediated Silica Deposition: An Effective Strategy for Bone Mineralization. ACS Biomater. Sci. Eng. 2022. [Google Scholar] [CrossRef]
- Yeo, K.B.; Ki, M.R.; Park, K.S.; Pack, S.P. Novel silica-forming peptides derived from Ectocarpus siliculosus. Process Biochem 2017, 58, 193–198. [Google Scholar] [CrossRef]
- Naik, R.R.; Brott, L.L.; Clarson, S.J.; Stone, M.O. Silica-precipitating peptides isolated from a combinatorial phage display peptide library. J. Nanosci. Nanotechnol. 2002, 2, 95–100. [Google Scholar] [CrossRef]
- Nguyen, T.K.M.; Ki, M.R.; Son, R.G.; Kim, K.H.; Hong, J.; Pack, S.P. A dual-functional peptide, Kpt from Ruegeria pomeroyi DSS-3 for protein purification and silica precipitation. Biochem. Eng. J. 2020, 163, 107726. [Google Scholar] [CrossRef]
- Luckarift, H.R.; Spain, J.C.; Naik, R.R.; Stone, M.O. Enzyme immobilization in a biomimetic silica support. Nat. Biotechnol. 2004, 22, 211–213. [Google Scholar] [CrossRef]
- Spinthaki, A.; Skordalou, G.; Stathoulopoulou, A.; Demadis, K.D. Modified macromolecules in the prevention of silica scale. Pure Appl. Chem. 2016, 88, 1037–1047. [Google Scholar] [CrossRef]
- Korsmeyer, R.W.; Gurny, R.; Doelker, E.; Buri, P.; Peppas, N.A. Mechanisms of solute release from porous hydrophilic polymers. Int. J. Pharm. 1983, 15, 25–35. [Google Scholar] [CrossRef]
- Dash, S.; Murthy, P.N.; Nath, L.; Chowdhury, P. Kinetic modeling on drug release from controlled drug delivery systems. Acta Pol. Pharm. 2010, 67, 217–223. [Google Scholar] [PubMed]
- Yourek, G.; McCormick, S.M.; Mao, J.J.; Reilly, G.C. Shear stress induces osteogenic differentiation of human mesenchymal stem cells. Regen. Med. 2010, 5, 713–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thu, H.E.; Mohamed, I.N.; Hussain, Z.; Shuid, A.N. Exploring molecular mechanism of bone-forming capacity of Eurycoma longifolia: Evidence of enhanced expression of bone-related biomarkers. J. Ayurveda Integr. Med. 2018, 9, 272–280. [Google Scholar] [CrossRef]
- Cheng, S.L.; Yang, J.W.; Rifas, L.; Zhang, S.F.; Avioli, L.V. Differentiation of human bone marrow osteogenic stromal cells in vitro: Induction of the osteoblast phenotype by dexamethasone. Endocrinology 1994, 134, 277–286. [Google Scholar] [CrossRef]
- Lee, S.; Lee, T.A.; Song, S.J.; Park, T.; Park, B. Hyperproduction of IL-6 caused by aberrant TDP-43 overexpression in high-fat diet-induced obese mice. FEBS Lett. 2015, 589, 1825–1831. [Google Scholar] [CrossRef] [Green Version]
- Lechner, C.C.; Becker, C.F. A sequence-function analysis of the silica precipitating silaffin R5 peptide. J. Pept. Sci. 2014, 20, 152–158. [Google Scholar] [CrossRef]
- Lechner, C.C.; Becker, C.F. Silaffins in Silica Biomineralization and Biomimetic Silica Precipitation. Mar. Drugs 2015, 13, 5297–5333. [Google Scholar] [CrossRef] [Green Version]
- Beederman, M.; Lamplot, J.D.; Nan, G.; Wang, J.; Liu, X.; Yin, L.; Li, R.; Shui, W.; Zhang, H.; Kim, S.H.; et al. BMP signaling in mesenchymal stem cell differentiation and bone formation. J. Biomed. Sci. Eng. 2013, 6, 32–52. [Google Scholar] [CrossRef] [Green Version]
- Abdelhamid, M.A.A.; Yeo, K.B.; Ki, M.R.; Pack, S.P. Self-encapsulation and controlled release of recombinant proteins using novel silica-forming peptides as fusion linkers. Int. J. Biol. Macromol. 2019, 125, 1175–1183. [Google Scholar] [CrossRef]
- Seyfaee, A.; Moreno-Atanasio, R.; Neville, F. Effect of physical conditioning of pH responsive polyamine nuclei and their subsequent silication. Colloids Surf. A Physicochem. Eng. Asp. 2018, 539, 243–251. [Google Scholar] [CrossRef]
- Choi, K.M.; Kuroda, K. Double function of tris(hydroxymethyl)aminomethane (THAM) for the preparation of colloidal silica nanospheres and the conversion to ordered mesoporous carbon. Chem. Commun. 2011, 47, 10933–10935. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, L.; Xie, J.; Wang, Y.; Wang, T.J. Surface Amination of Silica Nanoparticles Using Tris(hydroxymethyl)aminomethane. Ind. Eng. Chem. Res. 2020, 59, 21383–21392. [Google Scholar] [CrossRef]
- Richthammer, P.; Bormel, M.; Brunner, E.; van Pee, K.H. Biomineralization in diatoms: The role of silacidins. Chembiochem 2011, 12, 1362–1366. [Google Scholar] [CrossRef]
- Zhou, Y.; Shimizu, K.; Cha, J.N.; Stucky, G.D.; Morse, D.E. Efficient Catalysis of Polysiloxane Synthesis by Silicatein alpha Requires Specific Hydroxy and Imidazole Functionalities. Angew. Chem. Int. Ed. Engl. 1999, 38, 779–782. [Google Scholar] [CrossRef]
- von Burkersroda, F.; Schedl, L.; Gopferich, A. Why degradable polymers undergo surface erosion or bulk erosion. Biomaterials 2002, 23, 4221–4231. [Google Scholar] [CrossRef]
- Muller, S. Molecular dissection of protein antigens and the prediction of epitopes. In Synthetic Peptides as Antigens; Elsevier: Amsterdam, The Netherlands, 1999; pp. 1–78. [Google Scholar]
- Bharti, C.; Nagaich, U.; Pal, A.K.; Gulati, N. Mesoporous silica nanoparticles in target drug delivery system: A review. Int. J. Pharm. Investig. 2015, 5, 124–133. [Google Scholar] [CrossRef] [Green Version]
- Patra, J.K.; Das, G.; Fraceto, L.F.; Campos, E.V.R.; Rodriguez-Torres, M.d.P.; Acosta-Torres, L.S.; Diaz-Torres, L.A.; Grillo, R.; Swamy, M.K.; Sharma, S.; et al. Nano based drug delivery systems: Recent developments and future prospects. J. Nanobiotechnol. 2018, 16, 71. [Google Scholar] [CrossRef] [Green Version]
- Gessner, A.; Waicz, R.; Lieske, A.; Paulke, B.; Mader, K.; Muller, R.H. Nanoparticles with decreasing surface hydrophobicities: Influence on plasma protein adsorption. Int. J. Pharm. 2000, 196, 245–249. [Google Scholar] [CrossRef]
- Liu, T.; Zheng, Y.; Wu, G.; Wismeijer, D.; Pathak, J.L.; Liu, Y. BMP2-coprecipitated calcium phosphate granules enhance osteoinductivity of deproteinized bovine bone, and bone formation during critical-sized bone defect healing. Sci. Rep. 2017, 7, 41800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelhamid, M.A.A.; Son, R.G.; Park, K.S.; Pack, S.P. Oriented multivalent silaffin-affinity immobilization of recom-binant lipase on diatom surface: Reliable loading and high performance of biocatalyst. Colloids Surf. B Biointerfaces 2022, 219, 112830. [Google Scholar] [CrossRef] [PubMed]
- Ki, M.R.; Yeo, K.B.; Pack, S.P. Surface immobilization of protein via biosilification catalyzed by silicatein fused to glutathione S-transferase (GST). Bioprocess. Biosyst. Eng. 2013, 36, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, A.P.S.; Venkatasubbu, G.D. Sustained release of amoxicillin from hydroxyapatite nanocomposite for bone infections. Prog. Biomater. 2018, 7, 289–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ki, M.-R.; Park, K.S.; Abdelhamid, M.A.A.; Pack, S.P. Novel silicatein-like protein for biosilica production from Amphimedon queenslandica and its use in osteogenic composite fabrication. Korean J. Chem. Eng. 2023, 40, 419–428. [Google Scholar] [CrossRef]
- Nie, R.; Liu, Y.; Liu, Z. The calcium-binding activity of fish scale protein hydrolysates. J. Agric. Chem. Environ. 2014, 3, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Vimalraj, S. Alkaline phosphatase: Structure, expression and its function in bone mineralization. Gene 2020, 754, 144855. [Google Scholar] [CrossRef]
- Rawadi, G.; Vayssière, B.; Dunn, F.; Baron, R.; Roman-Roman, S. BMP-2 Controls Alkaline Phosphatase Expression and Osteoblast Mineralization by a Wnt Autocrine Loop. J. Bone Miner. Res. 2003, 18, 1842–1853. [Google Scholar] [CrossRef]
- Link, T.; Wang, X.; Schloßmacher, U.; Feng, Q.; Schröder, H.C.; Müller, W.E.G. An approach to a biomimetic bone scaffold: Increased expression of BMP-2 and of osteoprotegerin in SaOS-2 cells grown onto silica-biologized 3D printed scaffolds. RSC Adv. 2013, 3, 11140–11147. [Google Scholar] [CrossRef]
- Madl, C.M.; Mehta, M.; Duda, G.N.; Heilshorn, S.C.; Mooney, D.J. Presentation of BMP-2 mimicking peptides in 3D hydrogels directs cell fate commitment in osteoblasts and mesenchymal stem cells. Biomacromolecules 2014, 15, 445–455. [Google Scholar] [CrossRef]
- Kirsch, T.; Nickel, J.; Sebald, W. BMP-2 antagonists emerge from alterations in the low-affinity binding epitope for receptor BMPR-II. EMBO J. 2000, 19, 3314–3324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, H.C.; Larrouture, Q.C.; Li, Y.; Lin, H.; Beer-Stoltz, D.; Liu, L.; Tuan, R.S.; Robinson, L.J.; Schlesinger, P.H.; Nelson, D.J. Osteoblast Differentiation and Bone Matrix Formation in vivo and in vitro. Tissue Eng. Part B Rev. 2017, 23, 268–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallas, S.L.; Prideaux, M.; Bonewald, L.F. The osteocyte: An endocrine cell … and more. Endocr. Rev. 2013, 34, 658–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Medio, L.; Brandi, M.L. Advances in bone turnover markers. Adv. Clin. Chem. 2021, 105, 101–140. [Google Scholar] [CrossRef] [PubMed]
- Ressler, A.; Žužić, A.; Ivanišević, I.; Kamboj, N.; Ivanković, H. Ionic substituted hydroxyapatite for bone regeneration applications: A review. Open Ceram. 2021, 6, 100122. [Google Scholar] [CrossRef]
- Kim, B.S.; Yang, S.S.; Yoon, J.H.; Lee, J. Enhanced bone regeneration by silicon-substituted hydroxyapatite derived from cuttlefish bone. Clin. Oral Implants Res. 2017, 28, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.K.; Ki, M.R.; Kim, E.H.; Park, C.J.; Ryu, J.J.; Jang, H.S.; Pack, S.P.; Jo, Y.K.; Jun, S.H. Biosilicated collagen/β-tricalcium phosphate composites as a BMP-2-delivering bone-graft substitute for accelerated craniofacial bone regeneration. Biomater. Res. 2021, 25, 13. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (N-C) | AA # | Mass (Da) | pI | Net Charge |
|---|---|---|---|---|---|
| R5 | SSKKSGSYSGSKGSRRIL | 18 | 1885 | 11.64 | +5 |
| KPS | KPSHHHHHTGAN | 12 | 1359 | 9.81 | +1 |
| KPT | KPTHHHHHHDG | 11 | 1339 | 7.95 | +1 |
| HBM | RKRLKSSSKR | 10 | 1245 | 12.80 | +6 |
| P4 | KIPKASSVPTELSAISTLYL | 20 | 2118 | 9.41 | +1 |
| KP4A | AIPAASSVPTELSAISTLYL | 20 | 2003 | 3.20 | −1 |
| SP4G | KIPKAGGVPGELGAIGGLYL | 20 | 1910 | 9.41 | +1 |
| P4ΔLYL | KIPKASSVPTELSAIST | 17 | 1729 | 9.63 | +1 |
| 6 | KIPKASSVPTELSAISTLYLGSEEEEEE | 28 | 3036 | 3.68 | −5 |
| Initial P4 (μg) | Bound P4 (μg) | Carrier Mass (μg) | LE (%) | EE (%) | |
|---|---|---|---|---|---|
| P4/CLG | 100 | 77.30 ± 3.62 | 1000 | 7.15 | 77.30 |
| P4/HA | 100 | 51.58 ± 7.80 | 1000 | 4.91 | 51.58 |
| P4@Si | 100 | 68.00 ± 2.83 | 10.04 ± 0.06 | 87.18 | 68.00 |
| n | k (−n) | R2 | |
|---|---|---|---|
| P4/CLG | 0.2681 | 23.017 ± 5.350 | 0.9637 ± 0.001 |
| P4/HA | 0.1122 | 51.615 ± 5.190 | 0.9534 ± 0.019 |
| P4@Si | 0.5377 | 2.636 ± 0.178 | 0.9860 ± 0.011 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ki, M.-R.; Nguyen, T.K.M.; Park, T.-I.; Park, H.-M.; Pack, S.P. Biomimetic Silica Particles with Self-Loading BMP-2 Knuckle Epitope Peptide and Its Delivery for Bone Regeneration. Pharmaceutics 2023, 15, 1061. https://doi.org/10.3390/pharmaceutics15041061
Ki M-R, Nguyen TKM, Park T-I, Park H-M, Pack SP. Biomimetic Silica Particles with Self-Loading BMP-2 Knuckle Epitope Peptide and Its Delivery for Bone Regeneration. Pharmaceutics. 2023; 15(4):1061. https://doi.org/10.3390/pharmaceutics15041061
Chicago/Turabian StyleKi, Mi-Ran, Thi Khoa My Nguyen, Tae-In Park, Hae-Min Park, and Seung Pil Pack. 2023. "Biomimetic Silica Particles with Self-Loading BMP-2 Knuckle Epitope Peptide and Its Delivery for Bone Regeneration" Pharmaceutics 15, no. 4: 1061. https://doi.org/10.3390/pharmaceutics15041061
APA StyleKi, M. -R., Nguyen, T. K. M., Park, T. -I., Park, H. -M., & Pack, S. P. (2023). Biomimetic Silica Particles with Self-Loading BMP-2 Knuckle Epitope Peptide and Its Delivery for Bone Regeneration. Pharmaceutics, 15(4), 1061. https://doi.org/10.3390/pharmaceutics15041061