
Phenolic Fingerprint, Bioactivity and Nanoformulation of Prunus spinosa L. Fruit Extract for Skin Delivery

 ,
,  , , ,
, , ,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extract Preparation
2.3. LC-High-Resolution MS Analysis (HRMS)
2.4. HPLC-DAD Analysis
2.5. DPPH Assay
2.6. MIC and MBC Determination
2.7. Phospholipid Vesicle Preparation and Characterization
2.8. Small-Angle X-ray Scattering
2.9. Hemolytic Activity
2.10. Skin Cells Viability
2.11. Statistical Analysis
3. Results
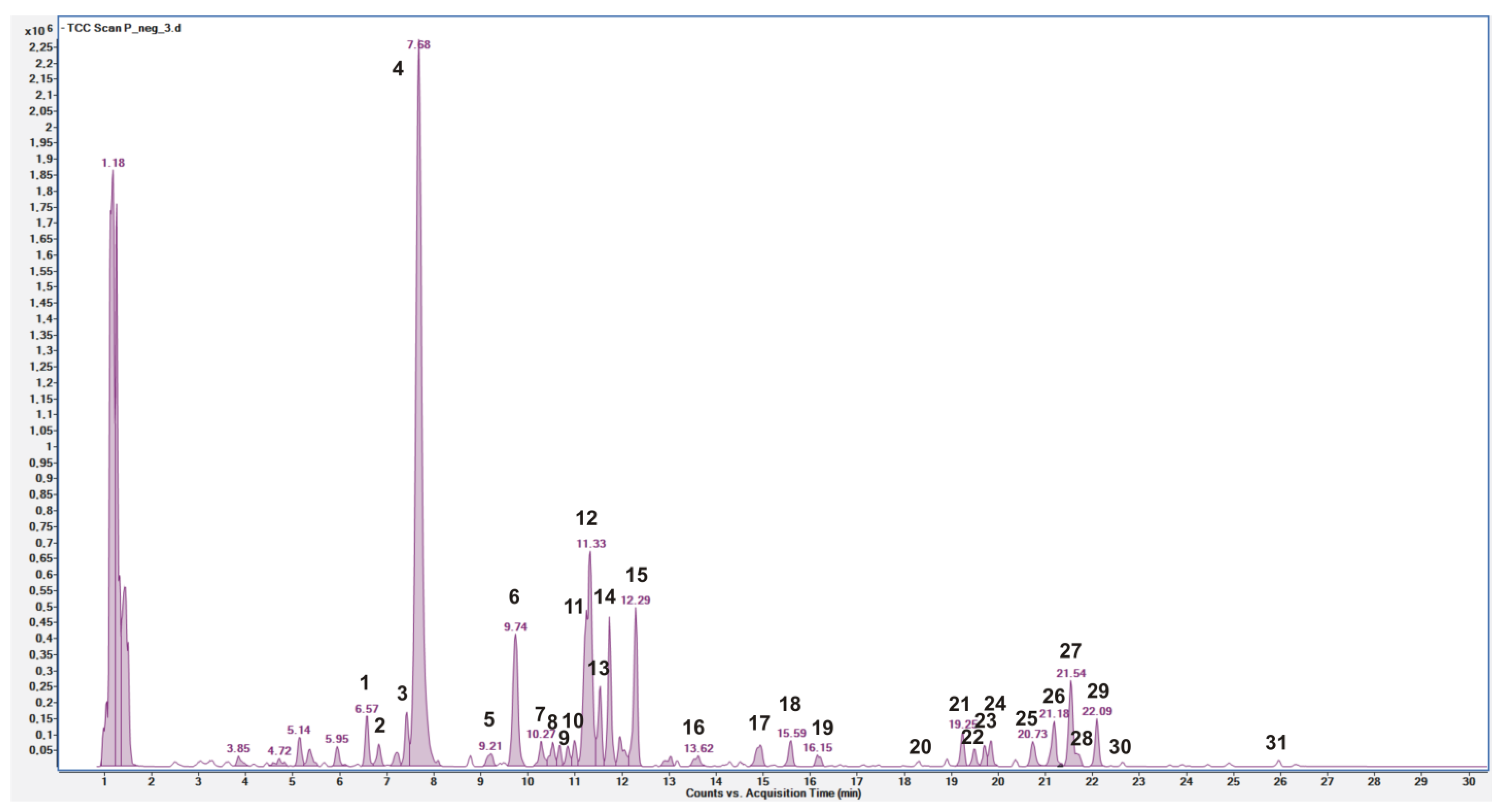
3.1. Quali-Quantitative Determination of Phenolic Compounds in P. spinosa Fruit Extract
3.2. Antioxidant Activity
3.3. Antibacterial Activity
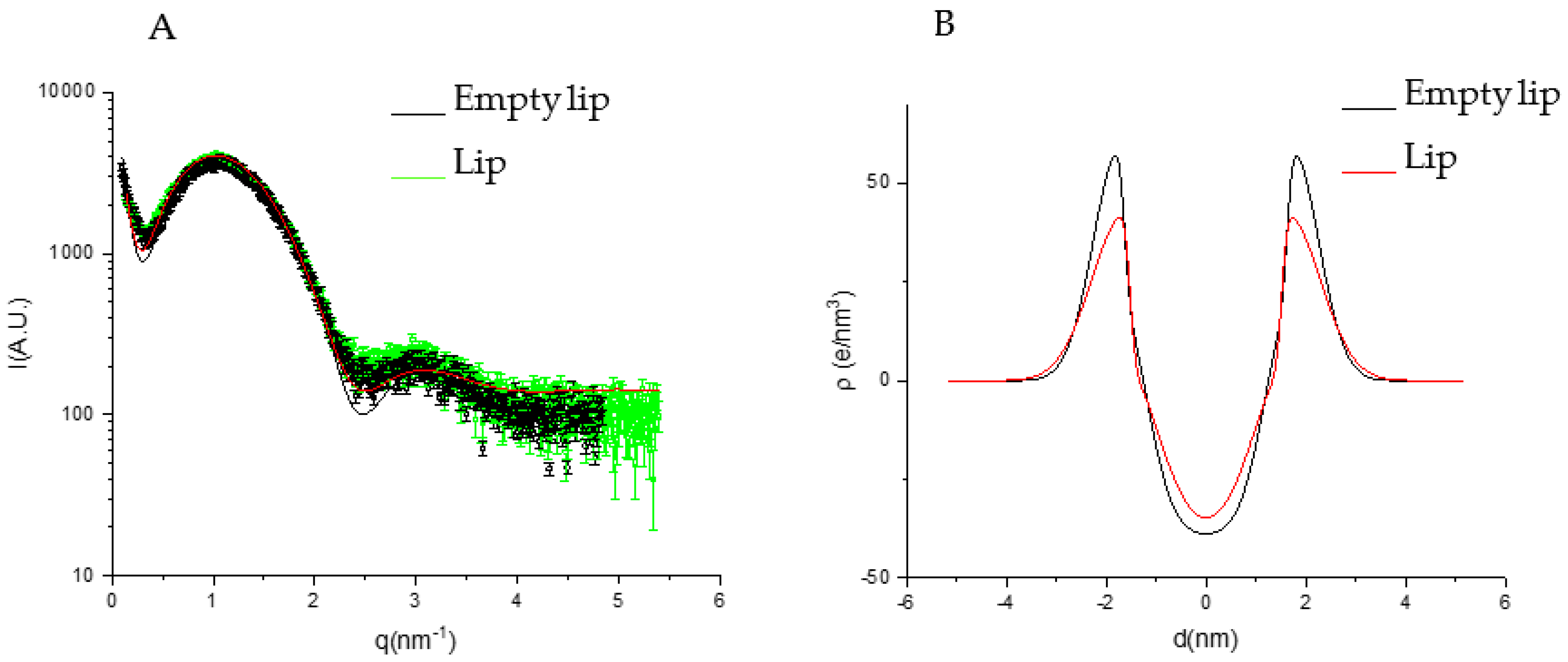
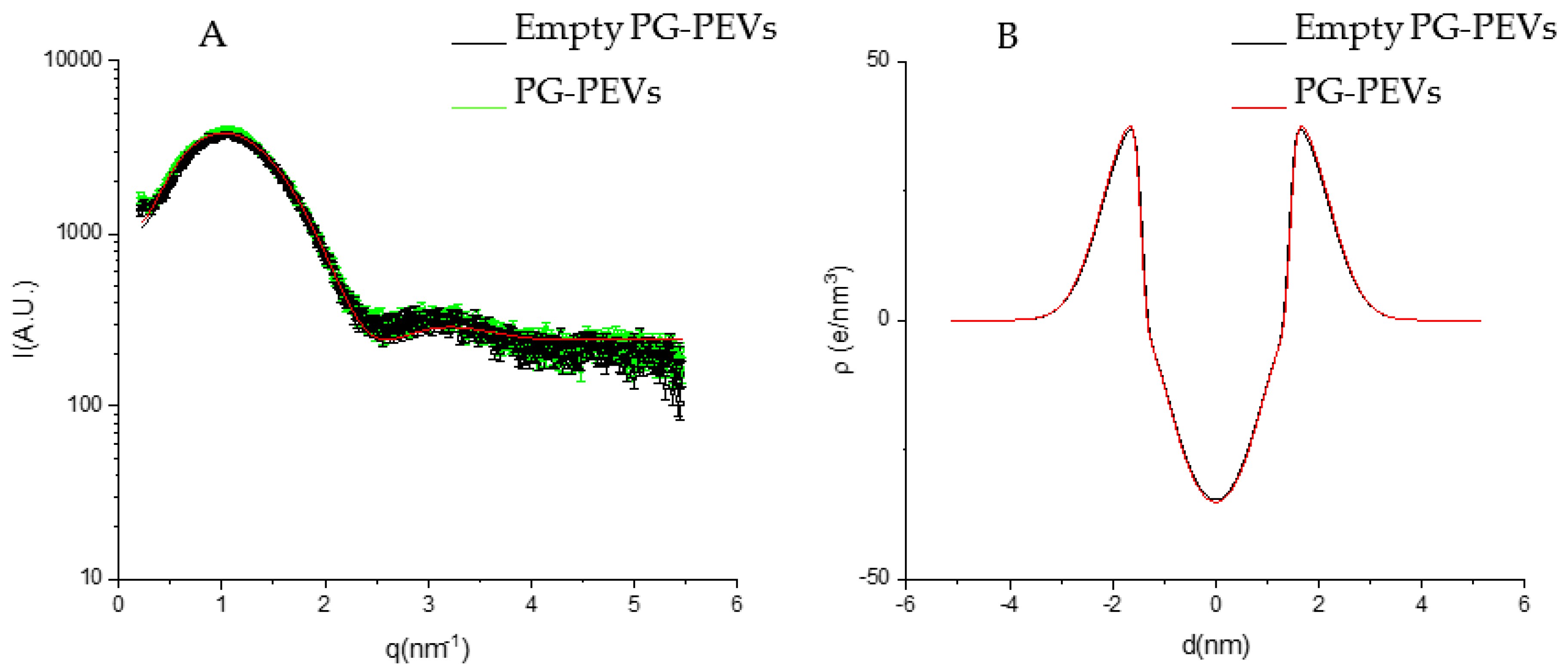
3.4. Phospholipid Vesicle Characterization
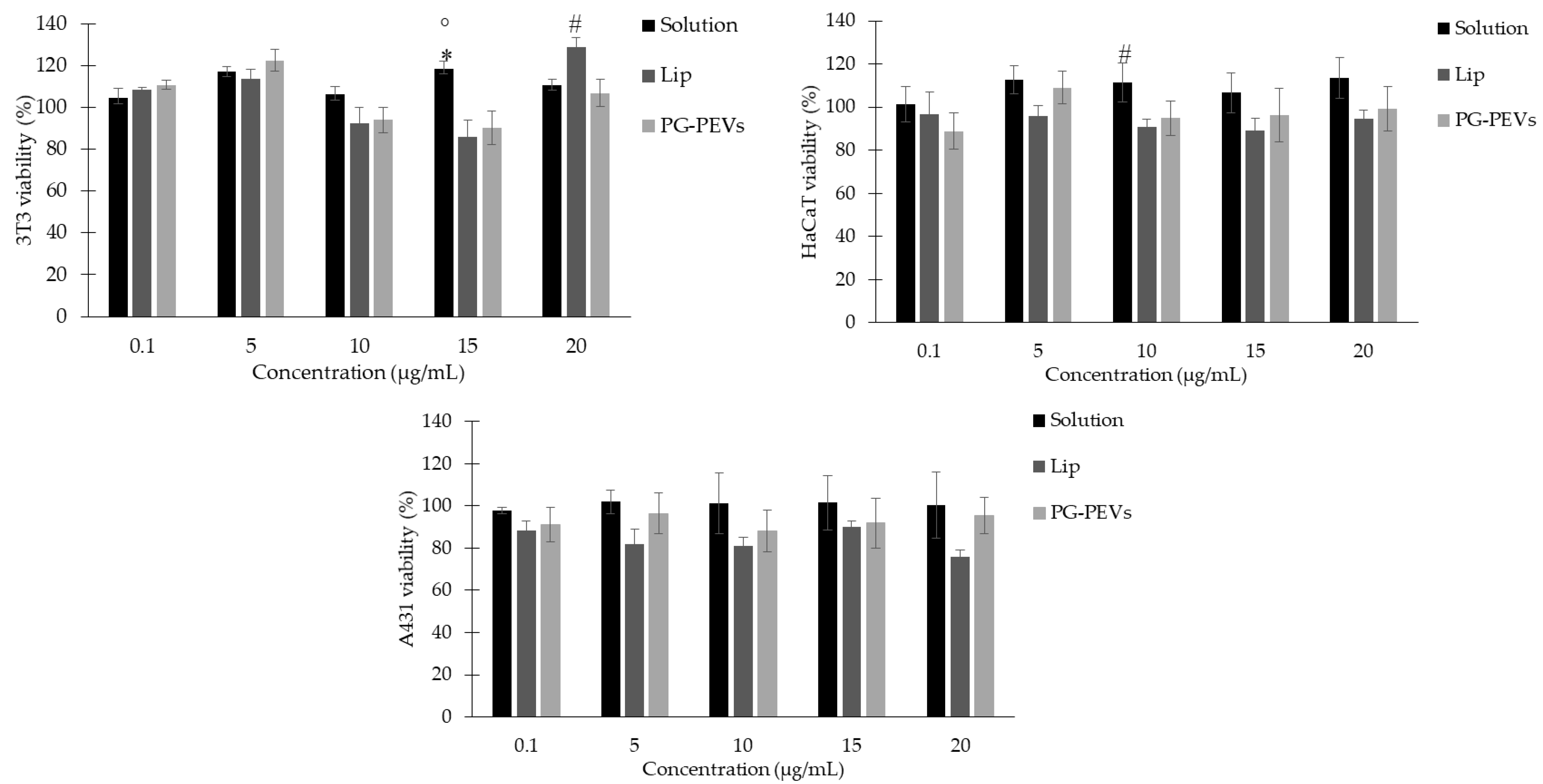
3.5. Biocompatibility Evaluation
4. Discussion
- -
- The extract is mainly localized in the hydrophilic core of the vesicles;
- -
- The extract does not affect the bilayer structure because of the low concentration or the low electron density contrast.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Magiera, A.; Czerwińska, M.E.; Owczarek, A.; Marchelak, A.; Granica, S.; Olszewska, M.A. Polyphenol-Enriched Extracts of Prunus spinosa Fruits: Anti-Inflammatory and Antioxidant Effects in Human Immune Cells Ex Vivo in Relation to Phytochemical Profile. Molecules 2022, 27, 1691. [Google Scholar] [CrossRef] [PubMed]
- Popović, B.M.; Blagojević, B.; Ždero Pavlović, R.; Mićić, N.; Bijelić, S.; Bogdanović, B.; Mišan, A.; Duarte, C.M.M.; Serra, A.T. Comparison between polyphenol profile and bioactive response in blackthorn (Prunus spinosa L.) genotypes from north Serbia-from raw data to PCA analysis. Food Chem. 2020, 302, 125373. [Google Scholar] [CrossRef] [PubMed]
- Capek, P.; Košťálová, Z. Isolation, chemical characterization and antioxidant activity of Prunus spinosa L. fruit phenolic polysaccharide-proteins. Carbohydr. Res. 2022, 515, 108547. [Google Scholar] [CrossRef] [PubMed]
- Pozzo, L.; Russo, R.; Frassinetti, S.; Vizzarri, F.; Árvay, J.; Vornoli, A.; Casamassima, D.; Palazzo, M.; Della Croce, C.M.; Longo, V. Wild Italian Prunus spinosa L. Fruit exerts in vitro antimicrobial activity and protects against in vitro and in vivo oxidative stress. Foods 2020, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backes, E.; Leichtweis, M.G.; Pereira, C.; Carocho, M.; Barreira, J.C.M.; Kamal Genena, A.; Baraldi, I.J.; Barreiro, M.F.; Barros, L.; Ferreira, I.C.F.R. Ficus carica L. and Prunus spinosa L. extracts as new anthocyanin-based food colorants: A thorough study in confectionery products. Food Chem. 2020, 333, 127457. [Google Scholar] [CrossRef]
- Coppari, S.; Colomba, M.; Fraternale, D.; Brinkmann, V.; Romeo, M.; Rocchi, M.B.L.; Di Giacomo, B.; Mari, M.; Guidi, L.; Ramakrishna, S.; et al. Antioxidant and Anti-Inflammaging Ability of Prune (Prunus spinosa L.) Extract Result in Improved Wound Healing Efficacy. Antioxidants 2021, 10, 374. [Google Scholar] [CrossRef] [PubMed]
- Murati, T.; Miletić, M.; Kolarić, J.; Lovrić, V.; Kovačević, D.B.; Putnik, P.; Jurčević, I.L.; Đikić, D.; Dragović-Uzelac, V.; Kmetič, I. Toxic activity of Prunus spinosa L. flower extract in hepatocarcinoma cells. Arh. Hig. Rada Toksikol. 2019, 70, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Marchelak, A.; Kolodziejczyk-Czepas, J.; Wasielewska, P.; Nowak, P.; Olszewska, M.A. The Effects of Prunus spinosa L. Flower Extracts, Model Polyphenols and Phenolic Metabolites on Oxidative/Nitrative Modifications of Human Plasma Components with Particular Emphasis on Fibrinogen In Vitro. Antioxidants 2021, 10, 581. [Google Scholar] [CrossRef]
- Meschini, S.; Pellegrini, E.; Condello, M.; Occhionero, G.; Delfine, S.; Condello, G.; Mastrodonato, F. Cytotoxic and Apoptotic Activities of Prunus spinosa Trigno Ecotype Extract on Human Cancer Cells. Molecules 2017, 22, 1578. [Google Scholar] [CrossRef] [Green Version]
- Magiera, A.; Czerwińska, M.E.; Owczarek, A.; Marchelak, A.; Granica, S.; Olszewska, M.A. Polyphenols and Maillard Reaction Products in Dried Prunus spinosa Fruits: Quality Aspects and Contribution to Anti-Inflammatory and Antioxidant Activity in Human Immune Cells Ex Vivo. Molecules 2022, 27, 3302. [Google Scholar] [CrossRef]
- Tiboni, M.; Coppari, S.; Casettari, L.; Guescini, M.; Colomba, M.; Fraternale, D.; Gorassini, A.; Verardo, G.; Ramakrishna, S.; Guidi, L.; et al. Prunus spinosa Extract Loaded in Biomimetic Nanoparticles Evokes In Vitro Anti-Inflammatory and Wound Healing Activities. Nanomaterials 2021, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Karakas, N.; Okur, M.E.; Ozturk, I.; Ayla, S.; Karadag, A.E.; Polat, D.Ç. Antioxidant Activity of Blackthorn (Prunus spinosa L.) Fruit extract and Cytotoxic Effects on Various Cancer Cell Lines. Medeni. Med. J. 2019, 34, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Condello, M.; Pellegrini, E.; Spugnini, E.P.; Baldi, A.; Amadio, B.; Vincenzi, B.; Occhionero, G.; Delfine, S.; Mastrodonato, F.; Meschini, S. Anticancer activity of “Trigno M”, extract of Prunus spinosa drupes, against in vitro 3D and in vivo colon cancer models. Biomed. Pharmacother. 2019, 118, 109281. [Google Scholar] [CrossRef] [PubMed]
- Balta, I.; Sevastre, B.; Mireşan, V.; Taulescu, M.; Raducu, C.; Longodor, A.L.; Marchiş, Z.; Mariş, C.S.; Coroian, A. Protective effect of blackthorn fruits (Prunus spinosa) against tartrazine toxicity development in albino Wistar rats. BMC Chem. 2019, 13, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alharbi, W.S.; Almughem, F.A.; Almehmady, A.M.; Jarallah, S.J.; Alsharif, W.K.; Alzahrani, N.M.; Alshehri, A.A. Phytosomes as an Emerging Nanotechnology Platform for the Topical Delivery of Bioactive Phytochemicals. Pharmaceutics 2021, 13, 1475. [Google Scholar] [CrossRef] [PubMed]
- Gugleva, V.; Ivanova, N.; Sotirova, Y.; Andonova, V. Dermal Drug Delivery of Phytochemicals with Phenolic Structure via Lipid-Based Nanotechnologies. Pharmaceuticals 2021, 14, 837. [Google Scholar] [CrossRef]
- Yang, B.; Dong, Y.; Wang, F.; Zhang, Y. Nanoformulations To Enhance the Bioavailability and Physiological Functions of Polyphenols. Molecules 2020, 25, 4613. [Google Scholar] [CrossRef] [PubMed]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: A rapid tool for turning tandem mass spectra into metabolite structure information. Nat. Methods 2019, 16, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, M.A.; Nothias, L.F.; Ludwig, M.; Fleischauer, M.; Gentry, E.C.; Witting, M.; Dorrestein, P.C.; Dührkop, K.; Böcker, S. High-confidence structural annotation of metabolites absent from spectral libraries. Nat. Biotechnol. 2022, 40, 411–421. [Google Scholar] [CrossRef]
- De Luca, M.; Lucchesi, D.; Tuberoso, C.I.G.; Fernàndez-Busquets, X.; Vassallo, A.; Martelli, G.; Fadda, A.M.; Pucci, L.; Caddeo, C. Liposomal Formulations to Improve Antioxidant Power of Myrtle Berry Extract for Potential Skin Application. Pharmaceutics 2022, 14, 910. [Google Scholar] [CrossRef]
- Garcia, M.T.; Ribosa, I.; González, J.J.; Comelles, F. Surface activity, self-aggregation and antimicrobial activity of catanionic mixtures of surface active imidazolium- or pyridinium-based ionic liquids and sodium bis(2-ethylhexyl) sulfosuccionate. J. Mol. Liq. 2020, 303, 112637. [Google Scholar] [CrossRef]
- Lennette, E.H.; Spaulding, E.H.; Truant, J.P. Manual of Clinical Microbiology; American Society for Microbiology Press: Washington, DC, USA, 1974; 970p. [Google Scholar] [CrossRef]
- Präbst, K.; Engelhardt, H.; Ringgeler, S.; Hübner, H. Basic Colorimetric Proliferation Assays: MTT, WST, and Resazurin. In Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2017; Volume 1601, pp. 1–17. [Google Scholar]
- Heftberger, P.; Kollmitzer, B.; Heberle, F.A.; Pan, J.; Rappolt, M.; Amenitsch, H.; Kučerka, N.; Katsaras, J.; Pabst, G. Global small-angle X-ray scattering data analysis for multilamellar vesicles: The evolution of the scattering density profile model. J. Appl. Crystallogr. 2014, 47, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Pabst, G.; Rappolt, M.; Amenitsch, H.; Laggner, P. Structural information from multilamellar liposomes at full hydration: Full q-range fitting with high quality X-ray data. Phys. Rev. E Stat. Phys. Plasmas Fluids Relat. Interdiscip. Top. 2000, 62, 4000–4009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, J.S. Analysis of small-angle scattering data from colloids and polymer solutions: Modeling and least-squares fitting. Adv. Colloid Interface Sci. 1997, 70, 171–210. [Google Scholar] [CrossRef]
- Caddeo, C.; Pucci, L.; Gabriele, M.; Carbone, C.; Fernàndez-Busquets, X.; Valenti, D.; Pons, R.; Vassallo, A.; Fadda, A.M.; Manconi, M. Stability, biocompatibility and antioxidant activity of PEG-modified liposomes containing resveratrol. Int. J. Pharm. 2018, 538, 40–47. [Google Scholar] [CrossRef]
- Caddeo, C.; Díez-Sales, O.; Pons, R.; Fernàndez-Busquets, X.; Fadda, A.M.; Manconi, M. Topical Anti-Inflammatory Potential of Quercetin in Lipid-Based Nanosystems: In Vivo and In Vitro Evaluation. Pharm. Res. 2014, 31, 959–968. [Google Scholar] [CrossRef] [Green Version]
- Pérez, L.; Pinazo, A.; Morán, M.C.; Pons, R. Aggregation Behavior, Antibacterial Activity and Biocompatibility of Catanionic Assemblies Based on Amino Acid-Derived Surfactants. Int. J. Mol. Sci. 2020, 21, 8912. [Google Scholar] [CrossRef]
- KNApSAcK Core System. Available online: http://www.knapsackfamily.com/knapsack_core/top.php (accessed on 11 December 2022).
- Blaženović, I.; Kind, T.; Ji, J.; Fiehn, O. Software Tools and Approaches for Compound Identification of LC-MS/MS Data in Metabolomics. Metabolites 2018, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Liu, M.; Lin, Z.; Jiang, H.; Niu, Y.; Wang, H.; Chen, S. Comprehensive identification of 125 multifarious constituents in Shuang-huang-lian powder injection by HPLC-DAD-ESI-IT-TOF-MS. J. Pharm. Biomed. Anal. 2015, 115, 86–106. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Stampar, F.; Veberic, R.; Sircelj, H. Wild Prunus Fruit Species as a Rich Source of Bioactive Compounds. J. Food Sci. 2016, 81, C1928–C1937. [Google Scholar] [CrossRef]
- Guimarães, R.; Barros, L.; Dueñas, M.; Carvalho, A.M.; Queiroz, M.J.R.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Characterisation of phenolic compounds in wild fruits from Northeastern Portugal. Food Chem. 2013, 141, 3721–3730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Vorsa, N.; Harrington, P.D.B.; Chen, P. Nontargeted Metabolomic Study on Variation of Phenolics in Different Cranberry Cultivars Using UPLC-IM—HRMS. J. Agric. Food Chem. 2018, 66, 12206–12216. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Lin, X.; Yang, C.H.; Tang, X.; Chang, Y.W.; Zheng, W.; Luo, L.; Xu, C.; Chen, Y.H. Study on Chemical Profile and Neuroprotective Activity of Myrica rubra Leaf Extract. Molecules 2017, 22, 1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaiswal, R.; Matei, M.F.; Ullrich, F.; Kuhnert, N. How to distinguish between cinnamoylshikimate esters and chlorogenic acid lactones by liquid chromatography-tandem mass spectrometry. J. Mass Spectrom. 2011, 46, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.R.; Liu, X.H.; Chen, L.; Huang, J.H.; Liang, X.J.; Wan, D.; Zhang, S.H.; Huang, L.Q. Comparison of the Antioxidant Activities and Phenolic Content of Five Lonicera Flowers by HPLC-DAD/MS-DPPH and Chemometrics. Int. J. Anal. Chem. 2020, 2020, 2348903. [Google Scholar] [CrossRef] [Green Version]
- Bottone, A.; Montoro, P.; Masullo, M.; Pizza, C.; Piacente, S. Metabolomics and antioxidant activity of the leaves of Prunus dulcis Mill. (Italian cvs. Toritto and Avola). J. Pharm. Biomed. Anal. 2018, 158, 54–65. [Google Scholar] [CrossRef]
- Sabatini, L.; Fraternale, D.; Di Giacomo, B.; Mari, M.; Albertini, M.C.; Gordillo, B.; Rocchi, M.B.L.; Sisti, D.; Coppari, S.; Semprucci, F.; et al. Chemical composition, antioxidant, antimicrobial and anti-inflammatory activity of Prunus spinosa L. fruit ethanol extract. J. Funct. Foods 2020, 67, 103885. [Google Scholar] [CrossRef]
- Luna-Vázquez-gómez, R.; Arellano-García, M.E.; Toledano-Magaña, Y.; García-Ramos, J.C.; Radilla-Chávez, P.; Salas-Vargas, D.S.; Casillas-Figueroa, F.; Ruiz-Ruiz, B.; Pestryakov, A.; Bogdanchikova, N. Bell Shape Curves of Hemolysis Induced by Silver Nanoparticles: Review and Experimental Assay. Nanomaterials 2022, 12, 1066. [Google Scholar] [CrossRef]
- Fischer, D.; Li, Y.; Ahlemeyer, B.; Krieglstein, J.; Kissel, T. In vitro cytotoxicity testing of polycations: Influence of polymer structure on cell viability and hemolysis. Biomaterials 2003, 24, 1121–1131. [Google Scholar] [CrossRef]
- Nagaraju, P.G.; Sengupta, P.; Chicgovinda, P.P.; Rao, P.J. Nanoencapsulation of clove oil and study of physicochemical properties, cytotoxic, hemolytic, and antioxidant activities. J. Food Process Eng. 2021, 44, e13645. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lecithin | Extract | PG | Water | |
|---|---|---|---|---|
| Lip | 180 mg | 20 mg | 1 mL | |
| Empty lip | 180 mg | 1 mL | ||
| PG-PEVs | 180 mg | 20 mg | 100 µL | 900 µL |
| Empty PG-PEVs | 180 mg | 100 µL | 900 µL |
| Compound No. | Rt min | Identity | [M-H]− § m/z | Molecular Formula | Δ ppm | MS/MS §,* m/z | References | Identification Confidencelevel # |
|---|---|---|---|---|---|---|---|---|
| 1 | 6.59 | vanillic acid-O-glucopyranoside | 329.0877 | C14H18O9 | −0.2134 | 167.0349(100) | [1,32] | 2 |
| 2 | 6.83 | caffeoylquinic acid | 353.0879 | C16H18O9 | −0.0047 | 191.0561(78)/179.0342(44) | [1] | 2 |
| 3 | 7.43 | phenolic glycoside | 313.0932 | C14H18O8 | - | 59.0140(100)/101.0245(39) | [18] | 3 |
| 4 | 7.68 | 3-O-caffeoylquinic acid | 353.0885 | C16H18O9 | −0.3257 | 191.0563(100)/179.0353(53) | [1,32,33,34] | 1 |
| 5 | 9.21 | coumaroylquinic acid isomer | 337.0930 | C16H18O8 | −0.3320 | 163.0401(100)/119.0505(64) | [18] | 2 |
| 6 | 9.74 | 3-p-coumaroylquinic acid | 337.0940 | C16H18O8 | −0.2511 | 163.0403(100)/119.0504(48) | [1,32,33,34] | 2 |
| 7 | 10.27 | unknown | 469.0979 | C20H22O13 | - | 191.0558(100)/59.0140(11) | - | 4 |
| 8 | 10.53 | caffeic acid hexoside | 341.0878 | C15H18O9 | 0.0021 | 179.0346(100)/135.0448(35) | [1] | 2 |
| 9 | 10.67 | unknown | 391.1251 | C16H24O11 | - | 44.9995(100) | - | 4 |
| 10 | 10.84 | caffeoylquinic acid hexoside | 515.1399 | C22H28O14 | −1.370 | 179.0339(100)/341.0853(35)/191.0554(17) | [18] | 3 |
| 11 | 11.18 | 3-O-feruloylquinic acid | 367.1043 | C17H20O9 | −0.4858 | 193.0516(100)/134.0372(77) | [1,32,33,34] | 2 |
| 12 | 11.33 | 5-O-caffeoylquinic acid | 353.0884 | C16H18O9 | 0.5943 | 191.0561(100)/179.0347(55) | [1,32,34] | 1 |
| 13 | 11.55 | hydroxybenzoyl-hexosyl-hexoside | 461.1302 | C19H26O13 | −0.4844 | 121.0295(100) | [32,35] | 3 |
| 14 | 11.73 | barlerin isomer I | 447.1508 | C19H28O12 | −0.7799 | 71.0141(100)/101.0242(41) | [32,36] | 3 |
| 15 | 12.29 | barlerin isomer II | 447.1508 | C19H28O12 | −0.7799 | 101.0242(100)/71.0146(46) | [32,36] | 3 |
| 16 | 13.62 | 4-p-coumaroylquinic acid | 337.0940 | C16H18O8 | −0.2511 | 173.0461(100)/163.0403(32) | [1,32,33,34] | 2 |
| 17 | 14.94 | caffeoylshikimic acid isomer I | 335.0773 | C16H16O8 | 0.0590 | 179.0352(100)/135.0448(75)/161.0247(43) | [1,32,37] | 2 |
| 18 | 15.59 | caffeoylshikimic acid isomer II | 335.0773 | C16H16O8 | 0.1590 | 161.0247(100)/135.0448(20)/179.0339(15) | [1,32,37] | 2 |
| 19 | 16.15 | ethyl caffeoylquinate | 381.1186 | C18H22O9 | −0.5058 | 161.0246(100) | [32,38] | 3 |
| 20 | 18.89 | quercetin hexosyl-rhamnoside | 609.1468 | C27H30O16 | 0.6916 | 300.0272(100)/301.0344(39)/343.0440(21) | [1,32,34] | 2 |
| 21 | 19.25 | quercetin-3-O-rutinoside | 609.1458 | C27H30O16 | −0.3084 | 300.0269(100)/301.0342(60)/343.0458(17) | [1,32,33,34] | 1 |
| 22 | 19.48 | quercetin-3-O-glucoside | 463.0886 | C21H20O12 | 0.4004 | 300.0270(100)/301.0322(41) | [1,32,33,34] | 1 |
| 23 | 19.70 | quercetin pentosyl-hexoside | 595.1303 | C26H28O16 | −0.1584 | 300.0280 (100)/301.0322(15)/415.0632(12) | [1,32,33,34] | 2 |
| 24 | 19.83 | quercetin hexoside | 463.0879 | C21H20O12 | −0.2996 | 300.0260(100)/301.0363(37) | [32,34] | 2 |
| 25 | 20.73 | quercetin pentoside | 433.0773 | C20H18O11 | −0.3349 | 300.0277(100)/301.0366(37) | [1,32,34] | 2 |
| 26 | 21.18 | quercetin pentoside | 433.0779 | C20H18O11 | −0.3249 | 300.0280(100) | [1,32,33] | 2 |
| 27 | 21.54 | quercetin pentoside | 433.0784 | C20H18O11 | 0.7651 | 300.0274(100)/301.0354(72) | [1,32,33] | 2 |
| 28 | 21.68 | quercetin hexosyl-rhamnoside | 609.1457 | C27H30O16 | −0.4084 | 301.0360(100)/300.0255(98) | [1,32,33] | 2 |
| 29 | 22.09 | quercetin-3-O-rhamnoside | 447.1006 | C21H20O11 | 0.0427 | 300.0267(100)/301.0351(71) | [1,32,33,34] | 1 |
| 30 | 22.63 | quercetin acetyl hexoside | 505.0992 | C23H22O13 | 0.3318 | 300.0261(100)/271.0227(30) | [1,32,34] | 2 |
| 31 | 25.96 | quercetin acetyl hexosyl-rhamnoside | 651.1561 | C29H32O17 | −0.5137 | 301.0349(100)/300.0267(50)/609.1430(15) | [1,32,34] | 2 |
| Compound | Peak No. § | P. spinosa Extract (mg/g dr) | |
|---|---|---|---|
| Mean | ±SD | ||
| Total Anthocyanins | 1.72 | 0.05 | |
| cyanidin-3-O-glucoside | A1 | 0.43 | 0.01 |
| cyanidin-3-O-rutinoside | A2 | 0.74 | 0.03 |
| peonidin-3-O-glucoside | A3 | 0.11 | 0.00 |
| peonidin-3-O-rutinoside | A4 | 0.44 | 0.01 |
| Total Flavonols | 1.33 | 0.01 | |
| quercetin-3-O-rutinoside | 21 | 0.31 | 0.00 |
| quercetin-3-O-glucoside | 22 | 0.06 | 0.00 |
| quercetin-3-O-rhamnoside | 29 | 0.12 | 0.00 |
| quercetin pentosides a | (25, 26, 27) | 0.49 | 0.00 |
| Other quercetin derivatives a | (20, 23, 24, 28, 30, 31) | 0.35 | 0.00 |
| Total Hydroxycinnamic acids | 2.72 | 0.02 | |
| 3-O-caffeoylquinic acid | 4 | 2.38 | 0.02 |
| 3-p-coumaroylquinic acid b | 6 | 0.13 | 0.00 |
| 3-O-feruoilquinic acid c | 11 | 0.07 | 0.00 |
| 5-O-caffeoylquinic acid | 12 | 0.13 | 0.00 |
| Total Hydroxybenzoic acids | 0.12 | 0.00 | |
| vanillic acid-O-glucopyranoside d | 1 | 0.12 | 0.00 |
| Total polyphenols | 5.92 | 0.08 | |
| Bacteria Strains | MIC (mg/mL) | MBC (mg/mL) |
|---|---|---|
| Bacillus subtilis | / | / |
| Staphylococcus epidermidis | 16 | 16 |
| Staphylococcus aureus | 16 | 16 |
| Listeria monocytogenes | / | / |
| Enterococcus faecalis | / | / |
| Escherichia coli | / | / |
| Acinetobacter baumannii | / | / |
| Klebsiella aerogenes | / | / |
| Lip | Empty Lip | PG-PEVs | Empty PG-PEVs | |
|---|---|---|---|---|
| Mean diameter (nm ± SD) | ** 94 ± 2.0 | 70 ± 2.6 | **,# 86 ± 6.4 | 59 ± 3.3 |
| Polydispersity index (± SD) | 0.19 ± 0.01 | 0.19 ± 0.03 | 0.21 ± 0.02 | 0.21 ± 0.03 |
| Zeta potential (mV ± SD) | −48 ± 1.4 | −46 ± 2.3 | −47 ± 1.7 | −47 ± 1.1 |
| Peak No. § | Compound | EE% ± SD | EE% ± SD |
|---|---|---|---|
| Lip | PG-PEVs | ||
| 2 | 3-O-caffeoylquinic acid | 33.6 ± 3.4 | 24.2 ± 1.4 |
| 4 | 3-O-feruoilquinic acid a | 35.3 ± 4.3 | 28.2 ± 1.7 |
| 5 | 5-O-caffeoylquinic acid | 38.0 ± 3.4 | 28.2 ± 1.5 |
| A1 | cyanidin-3-O-glucoside | 98.6 ± 0.9 | 84.9 ± 1.7 |
| A2 | cyanidin-3-O-rutinoside | 96.3 ± 0.9 | 74.4 ± 1.7 |
| A4 | peonidin-3-O-rutinoside | 97.0 ± 0.5 | 77.4 ± 0.9 |
| 14 | quercetin-3-O-rutinoside | 82.6 ± 6.1 | 64.5 ± 3.2 |
| 15 | quercetin-3-O-glucoside | 85.1 ± 7.8 | 72.8 ± 1.7 |
| 20 | quercetin pentoside b | 87.5 ± 7.5 | 70.7 ± 4.8 |
| Lip | Empty Lip | PG-PEVs | Empty PG-PEVs | |
|---|---|---|---|---|
| χ2 | 6.36 | 5.07 | 3.42 | 3.72 |
| ZH (Å) | 18.40 ± 0.5 | 17.27 ± 0.5 | 15.49 ± 0.5 | 15.29 ± 0.5 |
| σH (Å) | 5.91 ± 0.5 | 5.30 ± 0.5 | 6.43 ± 0.5 | 6.47 ± 0.5 |
| σC (Å) | 17.73 ± 10 | 4.62 × 10−5 ± 1 | 4.27 × 10−4 ± 1 | 4.27 × 10−4 ± 1 |
| Extract Concentration (µg/mL) | Hemolytic Activity (% ± SD) | |
|---|---|---|
| Solution | 1000 | 2.00 ± 0.32 |
| Lip | 1000 | 0.66 ± 0.46 |
| PEVs | 1000 | 0.66 ± 0.52 |
| Solution | 2000 | 2.61 ± 0.71 |
| Lip | 2000 | 1.20 ± 0.62 |
| PEVs | 2000 | * 0.65 ± 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Luca, M.; Tuberoso, C.I.G.; Pons, R.; García, M.T.; Morán, M.d.C.; Ferino, G.; Vassallo, A.; Martelli, G.; Caddeo, C. Phenolic Fingerprint, Bioactivity and Nanoformulation of Prunus spinosa L. Fruit Extract for Skin Delivery. Pharmaceutics 2023, 15, 1063. https://doi.org/10.3390/pharmaceutics15041063
De Luca M, Tuberoso CIG, Pons R, García MT, Morán MdC, Ferino G, Vassallo A, Martelli G, Caddeo C. Phenolic Fingerprint, Bioactivity and Nanoformulation of Prunus spinosa L. Fruit Extract for Skin Delivery. Pharmaceutics. 2023; 15(4):1063. https://doi.org/10.3390/pharmaceutics15041063
Chicago/Turabian StyleDe Luca, Maria, Carlo Ignazio Giovanni Tuberoso, Ramon Pons, María Teresa García, María del Carmen Morán, Giulio Ferino, Antonio Vassallo, Giuseppe Martelli, and Carla Caddeo. 2023. "Phenolic Fingerprint, Bioactivity and Nanoformulation of Prunus spinosa L. Fruit Extract for Skin Delivery" Pharmaceutics 15, no. 4: 1063. https://doi.org/10.3390/pharmaceutics15041063
APA StyleDe Luca, M., Tuberoso, C. I. G., Pons, R., García, M. T., Morán, M. d. C., Ferino, G., Vassallo, A., Martelli, G., & Caddeo, C. (2023). Phenolic Fingerprint, Bioactivity and Nanoformulation of Prunus spinosa L. Fruit Extract for Skin Delivery. Pharmaceutics, 15(4), 1063. https://doi.org/10.3390/pharmaceutics15041063