
Molluscicidal and Larvicidal Potency of N-Heterocylic Analogs against Biomophalaria alexandrina Snails and Schistosoma mansoni Larval Stages
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
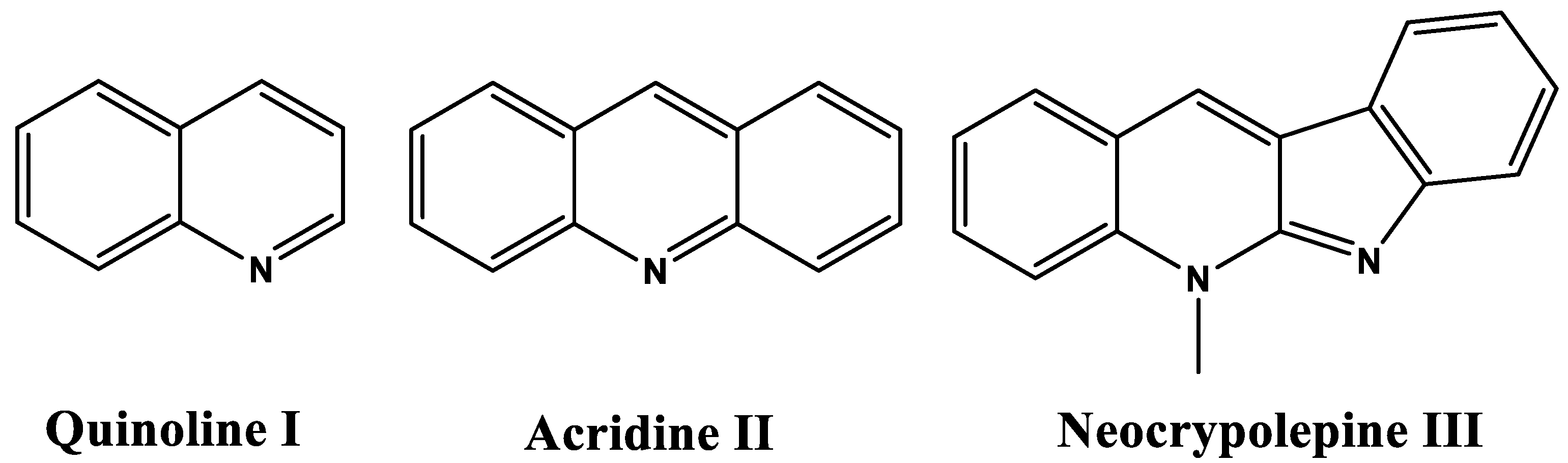
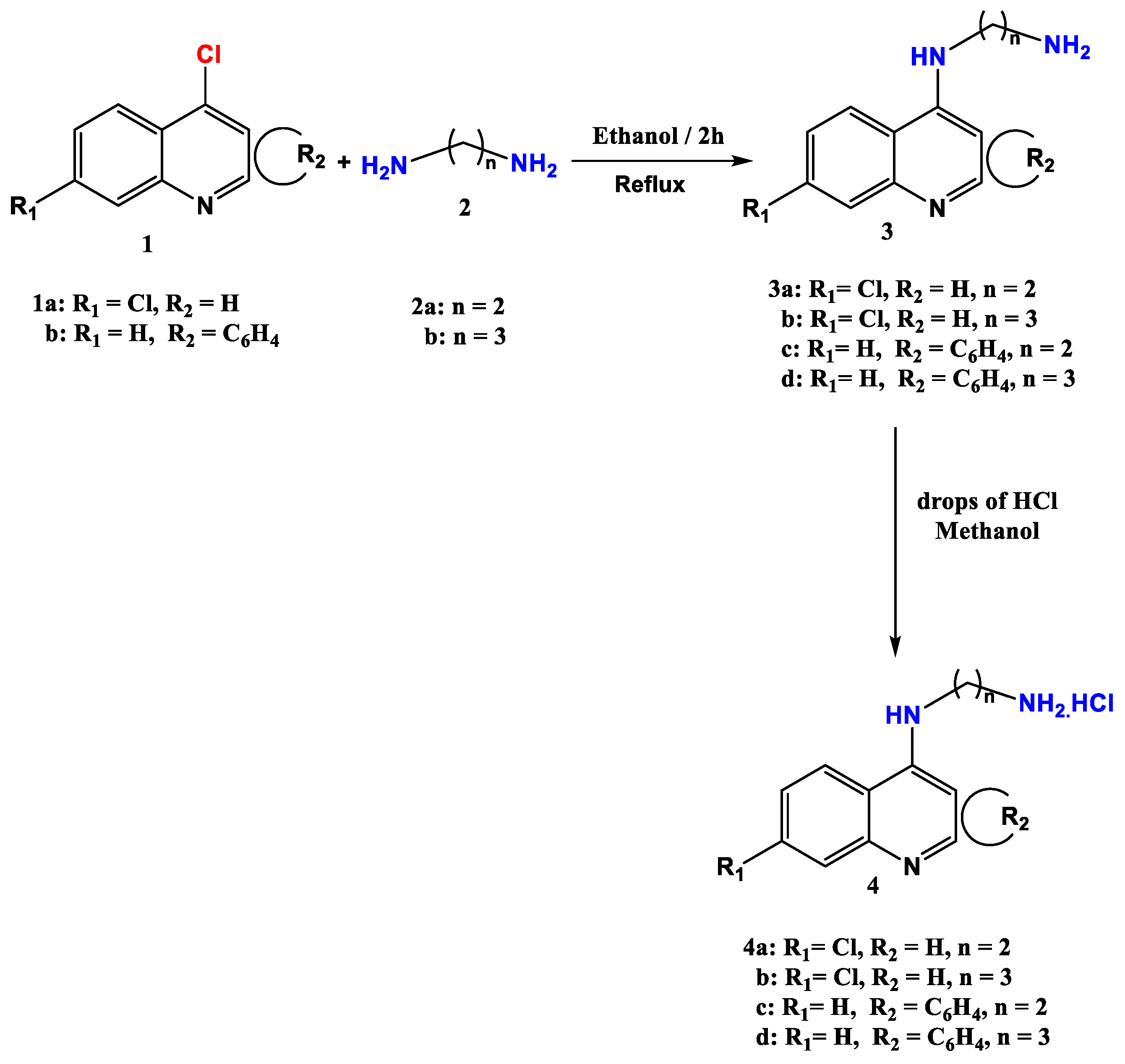
2.1. Chemistry
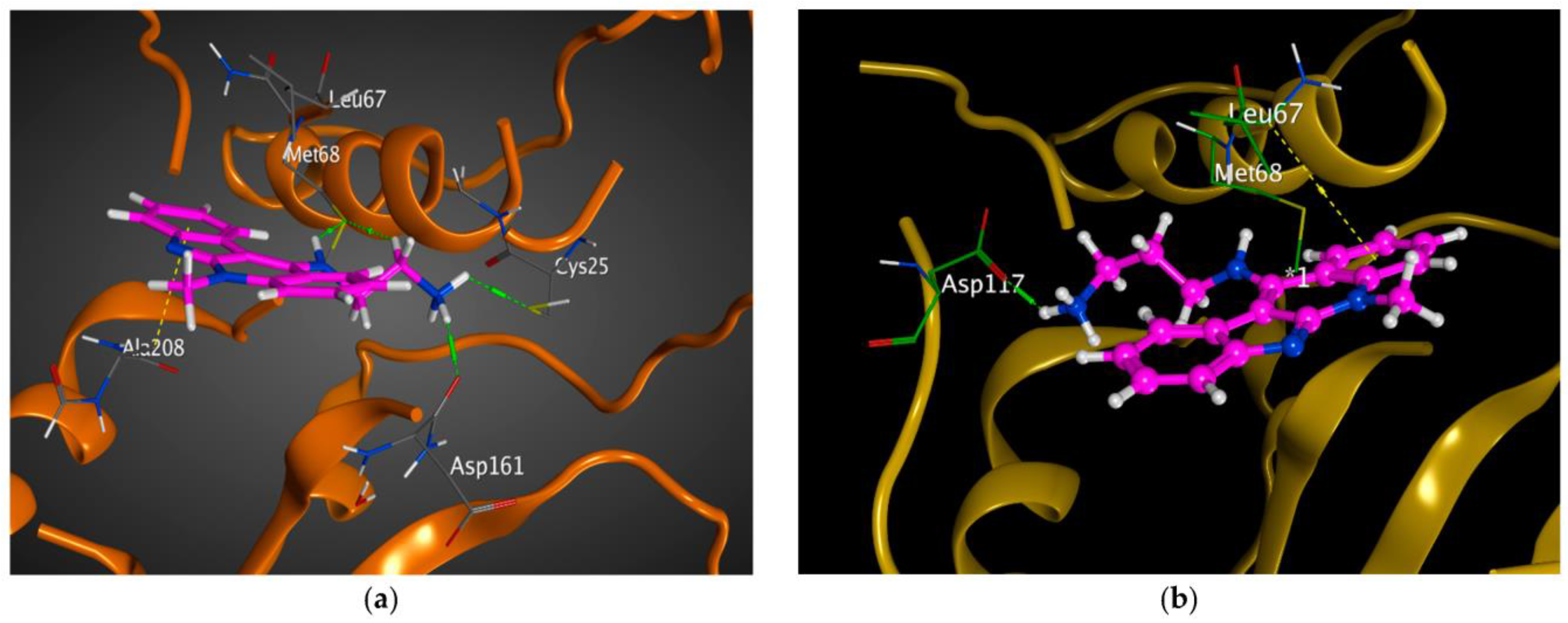
2.2. Modeling Studies
2.3. Biological Study
2.3.1. Snails
2.3.2. Miracidia
2.3.3. Cercariae
2.3.4. Snails Breeding and Maintenance
2.3.5. Snails Infection and Larval Stages Maintenance
2.4. Toxicity Test
2.4.1. Molluscicidal Activity
2.4.2. Larvicidal Activity
Miracidicidal Activity
Cercaricidal Activity
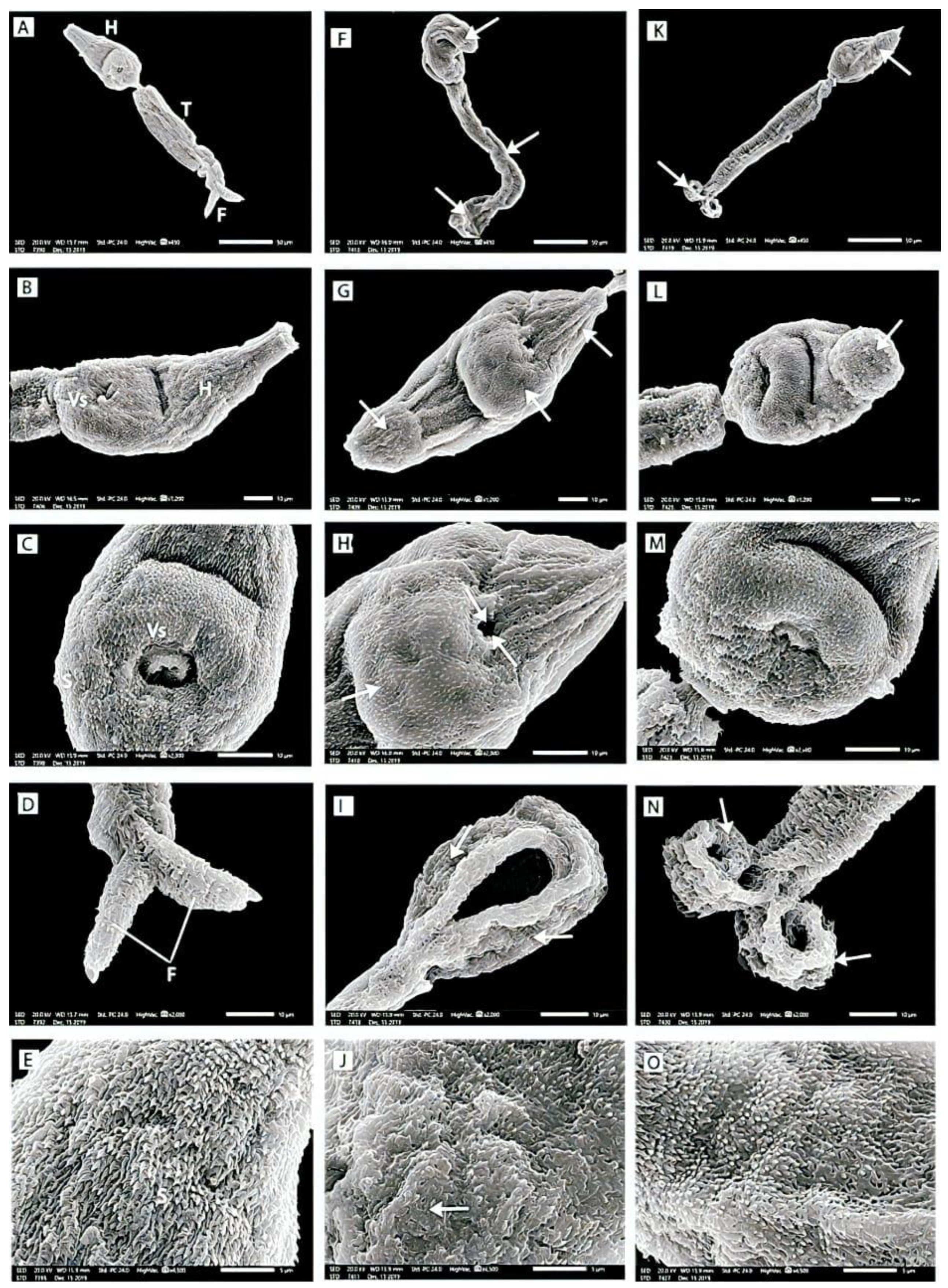
2.5. Scanning Electron Microscope Study of S. mansoni Cercariae
2.6. Experimental Design
2.6.1. Egg-Laying Capacity
2.6.2. Hatchability of Eggs of B. alexandrina In Vitro
2.6.3. Statistical Analysis
3. Results
3.1. Chemistry
3.2. Modeling Studies
3.3. The Toxic Effects of Synthesized N-Heteroarene Derivatives
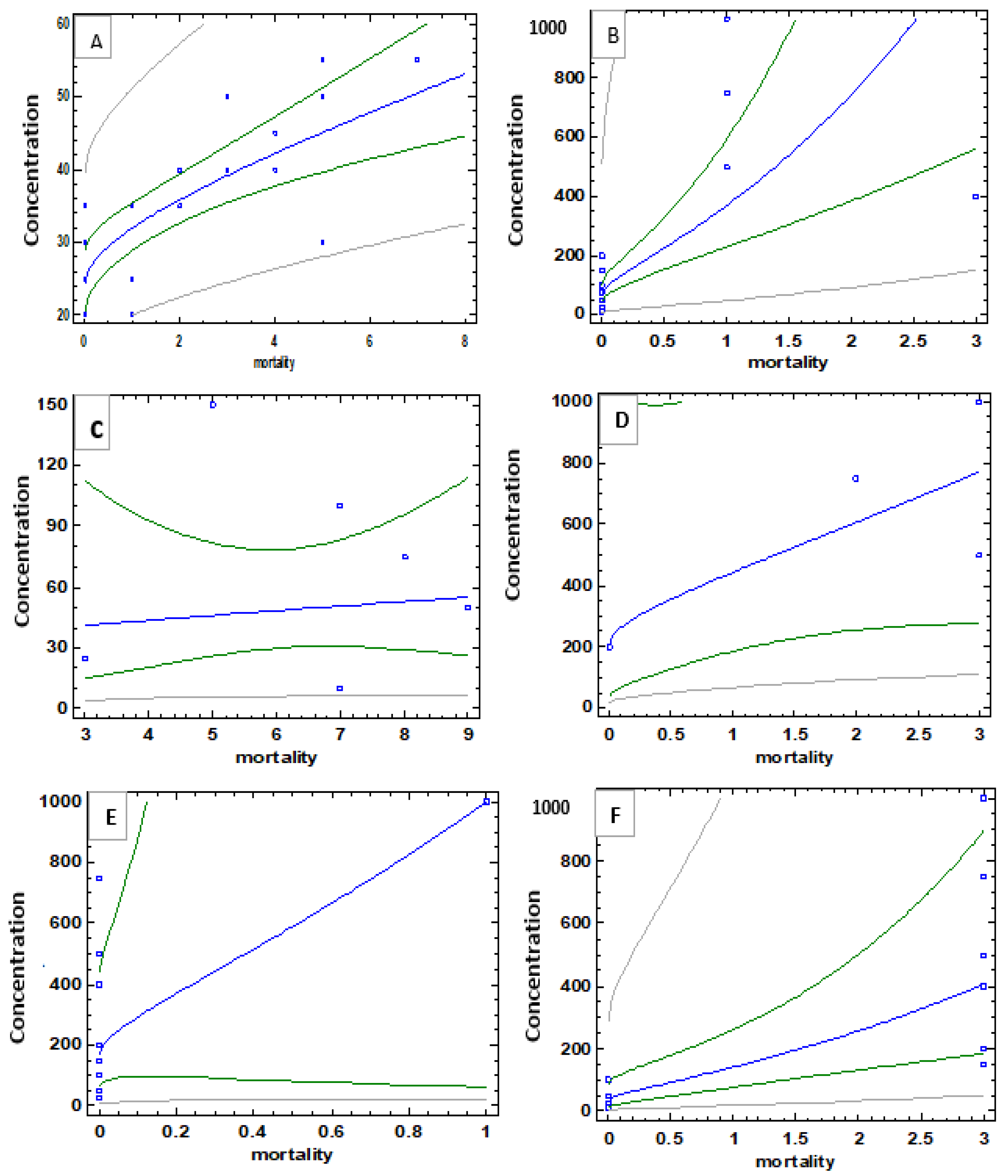
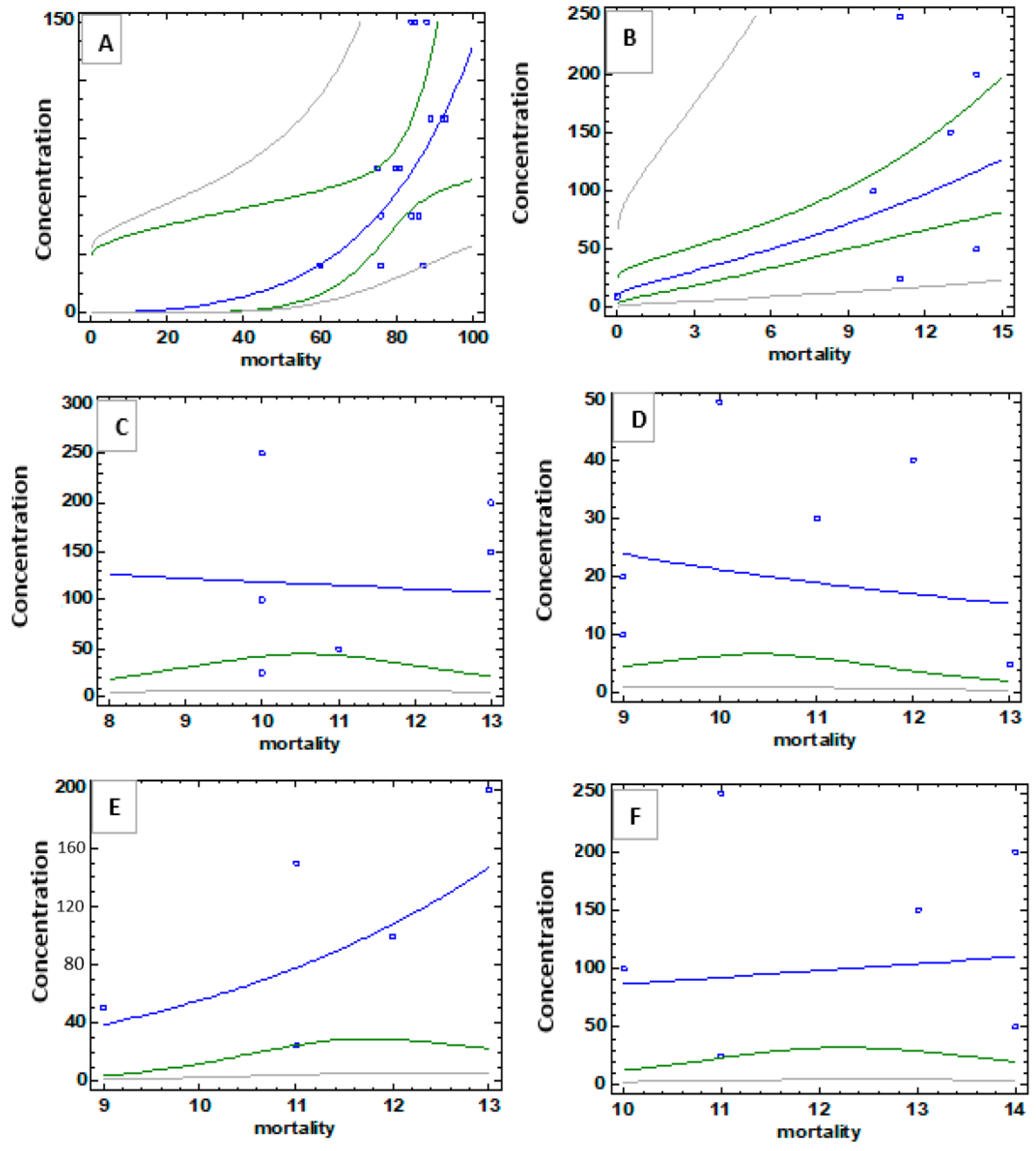
3.3.1. Molluscicidal Activity
3.3.2. Larvicidal Activity
3.3.3. Effect of CAAQ on Biological Activities of B. alexandrina Snails
Egg-Laying Capacity
3.3.4. Effect of CAAQ on Eggs’ Hatchability of B. alexandrina In Vitro
3.3.5. Scanning Electron Microscopy (SEM) of S. mansoni Cercariae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- W.H.O. Schistosomiasis. 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/schistosomiasis (accessed on 3 February 2023).
- King, C.H.; Bertsch, D. Historical perspective: Snail control to prevent schistosomiasis. PLoS Negl. Trop. Dis. 2015, 9, e0003657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coustau, C.; Théron, A. Resistant or resisting: Seeking consensus terminology. Trends Parasitol. 2004, 20, 209–210. [Google Scholar] [CrossRef]
- El-Atti, A.; Elsheakh, A.; Khalil, A.E.; Elgohary, W. Control of the glassy clover snails Monacha cartusiana using Zingiber officinale extract as an ecofriendly molluscicide. Afr. J. Biol. Sci. 2019, 15, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.M.; Ahmed, A.K.; Hammam, O.A.; Abdel-Ghaffar, F. Immunotoxical, neurotoxical, histopathological and immunohistopathological alterations of Nerium oleander and Tecoma stans methanolic extract on Biomphalaria alexandrina snails. Acta Trop. 2022, 230, 106405. [Google Scholar] [CrossRef] [PubMed]
- Femoe, U.M.; Fassi, J.B.K.; Jatsa, H.B.; Tchoffo, Y.L.; Nna, D.C.A.; Kamdoum, B.C.; Dimo, T. In Vitro Assessment of the Cercaricidal Activity of Sida acuta Burm. F. and Sida rhombifolia Linn. (Malvaceae) Hydroethanolic Extracts, Cytotoxicity, and Phytochemical Studies. J. Evid. Based Complement. Altern. Med. 2022, 2022, 7281144. [Google Scholar]
- Sheir, S.K.; Mohamed, A.H.; Osman, G.Y.; El-Nabi, S.E.H.; Allam, S.A. Efficiency of Citrus limon extract on biological and molecular activities of Biomphalaria alexandrina snails. Delta J. Sci. 2013, 36, 186–200. [Google Scholar] [CrossRef]
- Elmongy, E.I.; Ahmed, A.A.; El Sayed, I.E.T.; Fathy, G.; Awad, H.M.; Salman, A.U.; Hamed, M.A. Synthesis, Biocidal and Antibiofilm Activities of New Isatin–Quinoline Conjugates against Multidrug-Resistant Bacterial Pathogens along with Their In Silico Screening. J. Antibiot. 2022, 11, 1507. [Google Scholar] [CrossRef]
- De la Guardia, C.; Stephens, D.E.; Dang, H.T.; Quijada, M.; Larionov, O.V.; Lleonart, R. Antiviral Activity of Novel Quinoline Derivatives against Dengue Virus Serotype 2. Molecules 2018, 23, 672. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, E.A.; Ferreira, A.T.; Pinz, M.P.; REIS, A.S.; Vogt, A.G.; Stein, A.L.; Luchese, C. Antioxidant effect of quinoline derivatives containing or not selenium: Relationship with antinociceptive action quinolines are antioxidant and antinociceptive. An. Acad. Bras. Cienc. 2017, 89, 457–467. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Mishra, A. Synthesis, Characterization & Screening for Anti-Inflammatory & Analgesic Activity of Quinoline Derivatives Bearing Azetidinones Scaffolds. Antiinflamm. Antiallergy Agents Med. Chem. 2016, 15, 31–43. [Google Scholar]
- Lauria, A.; La Monica, G.; Bono, A.; Martorana, A. Quinoline anticancer agents active on DNA and DNA-interacting proteins: From classical to emerging therapeutic targets. Eur. J. Med. Chem. 2021, 220, 113555. [Google Scholar] [CrossRef] [PubMed]
- Solomon, V.R.; Pundir, S.; Lee, H. Examination of Novel 4-Aminoquinoline Derivatives Designed and Synthesized by a Conjugates Pharmacophore Approach to Enhance Their Anticancer Activities. Sci. Rep. 2019, 9, 6315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taman, A.; Alhusseiny, S.M.; Saleh, N.E.; Youssef, M.Y.; Mansour, B.; Massoud, M.; El-Beshbishi, S.N. Effect of a newly synthesized quinoline-based compound (PPQ-8) on murine schistosomiasis mansoni. J. Helminthol. 2020, 94, e123. [Google Scholar] [CrossRef] [PubMed]
- Kudryavtseva, T.N.; Lamanov, A.Y.; Klimova, L.G.; Nazarov, G.V. Synthesis and Antimicrobial Activity of AcridineCarboxylic Acid Derivatives Containing a Piperazine Moiety. Russ. Chem. Bull. 2017, 66, 123–128. [Google Scholar] [CrossRef]
- Wynn, J.E.; Zhang, W.; Falkinham, J.O.; Santos, W.L. Branched Peptides: Acridine and Boronic Acid Derivatives as Antimicrobial Agent A.C.S. Med. Chem. Lett. 2017, 8, 820–823. [Google Scholar] [CrossRef]
- Kumar, R.; Kaur, M.; Kumari, M. Acridine: A Versatile Heterocyclic Nucleus. Acta Pol. Pharm.-Drug Res. 2012, 69, 3–9. [Google Scholar]
- Pereira, E.; Quental, L.D.; Palma, E.; Oliveira, M.C.; Mendes, F.; Raposinho, P.; Correia, I.; Lavrado, J.; Di Maria, S.; Belchior, A.; et al. Evaluation of Acridine Orange Derivatives as DNA-Targeted Radiopharmaceuticals for Auger Therapy: Influence of the Radionuclide and Distance to DNA. Sci. Rep. 2017, 7, 42544. [Google Scholar] [CrossRef] [Green Version]
- Caffrey, C.R.; Steverding, D.; Swenerton, R.K.; Kelly, B.; Walshe, D.; Debnath, A.; May, B.C. Bis-acridines as lead antiparasitic agents: Structure-activity analysis of a discrete compound library in vitro. Antimicrob. Agents Chemother. 2007, 51, 2164–2172. [Google Scholar] [CrossRef] [Green Version]
- Kožurková, M.; Sabolová, D.; Kristian, P. A review on acridinylthioureas and its derivatives: Biological and cytotoxic activity. J. Appl. Toxicol. 2017, 37, 1132–1139. [Google Scholar] [CrossRef]
- Wang, N.; Wicht, K.J.; Wang, L.; Lu, W.J.; Misumi, R.; Wang, M.Q.; Inokuchi, T. Synthesis and in vitro testing of antimalarial activity of non-natural-type neocryptolepines: Structure–activity relationship study of 2, 11-and 9, 11-disubstituted 6-methylindolo [2, 3-b] quinolines. Chem. Pharm. Bull. 2013, 61, 1282–1290. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.A.; Awad, H.M.; El-Sayed, I.E.T.; El Gokha, A.A. Synthesis and antiproliferative activity of new hybrids bearing neocryptolepine, acridine and α-aminophosphonate scaffolds. J. Iran. Chem. Soc. 2020, 17, 1211–1221. [Google Scholar] [CrossRef]
- Mei, Z.; Wang, L.; Lu, W.; Pang, C.; Maeda, T.; Peng, W.; Kaiser, M.; El-Sayed, I.; Inokuchi, T. Synthesis and in vitro antimalarial testing of neocryptolepines: SAR study for improved activity by introduction and modification of side chains at C2 and C11 on 5H-indolo[2,3-b]quinolones. J. Med. Chem. 2013, 56, 1431–1442. [Google Scholar] [CrossRef]
- Lu, W.J.; Świtalska, M.; Wang, L.; Yonezawa, M.; El-Sayed, I.E.T.; Wietrzyk, J.; Inokuchi, T. In vitro antiproliferative activity of 11-aminoalkylamino-substituted 5H-indolo[2,3-b]quinolines; improving activity of neocryptolepines by installation of ester substituent. Med Chem Res. 2013, 22, 4492–4504. [Google Scholar] [CrossRef]
- EL Sayed, I.; Ramzy, F.; William, S.; Bahanasawy, M.E.; El-Sattar, A.; Mostafa, M. Neocryptolepine analogues containing N-substituted side-chains at C-11: Synthesis and antischistosomicidal activity. Med. Chem. Res. 2012, 21, 4219–4229. [Google Scholar] [CrossRef]
- El Bardicy, S.; El Sayed, I.; Yousif, F.; Van der Veken, P.; Haemers, A.; Augustyns, K.; Pieters, L. Schistosomicidal and molluscicidal activities of aminoalkylamino substituted neo-and norneocryptolepine derivatives. Pharm. Biol. 2012, 50, 134–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayed, I.E.T.E.; Ullah, S.; Al-Hartomy, O.A.; Hasanein, A.M.; Ahmed, A.A.; Kahilo, K.A.; El-Naggar, M.E. Synthesis, Nanoformulations, and In Vitro Anticancer Activity of N-Substituted Side Chain Neocryptolepine Scaffolds. Molecules 2022, 27, 1024. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, F.A.R.; García-Sánchez, R.N.; Estupiñan, S.V.; Gómez-Barrio, A.; Amado, D.F.T.; Pérez-Solórzano, B.M.; Kouznetsov, V.V. Synthesis and antimalarial activity of new heterocyclic hybrids based on chloroquine and thiazolidinone scaffolds. Bioorganic Med. Chem. 2011, 19, 4562–4573. [Google Scholar] [CrossRef]
- Iain, D.K.; Lee, J.H.; Farady, C.J.; Marion, R.; Rickert, M.; Sajid, M.; Brinen, L.S. Vinyl sulfones as antiparasitic agents and a structural basis for drug design. J. Biol. Chem. 2009, 284, 25697–25703. [Google Scholar]
- Brinen, L.S.; Marion, R. PDB Entry—2P7U. The Crystal Structure of Rhodesain, the Major Cysteine Protease of T. brucei Rhodesiense, Bound to Inhibitor K777; wwPDB: New York, NY, USA, 2007. [Google Scholar] [CrossRef]
- Elmongy, E.I.; Altwaijry, N.; Attallah, N.G.; AlKahtani, M.M.; Henidi, H.A. In-Silico Screening of Novel Synthesized Thienopyrimidines Targeting Fms Related Receptor Tyrosine Kinase-3 and Their In-Vitro Biological Evaluation. Pharmaceuticals 2022, 15, 170. [Google Scholar] [CrossRef]
- Elmongy, E.I.; Henidi, H.A. In Silico Evaluation of a Promising Key Intermediate Thieno [2, 3-d] Pyrimidine Derivative with Expected JAK2 Kinase Inhibitory Activity. Molbank 2022, 2022, M1352. [Google Scholar] [CrossRef]
- Oliver, L.; Haskins, W.T.; Gurian, J. Action of very low concentration of Na pentachlorophenate on freshly laid eggs of Australorbis glabratus. Bull. WHO 1962, 27, 87–94. [Google Scholar]
- Youssef, F.; El-Emam, M.; El-Gindy, H.; El-Sayed, K. Life table and population parameters of Biomphalaria Alexandria (Pulmonata: Planobidae), the intermediate host of Schistosoma mansoni in Egypt. J. Egypt. Soc. Parasitol. 1999, 79, 273–279. [Google Scholar]
- Chernin, E. Behavioral responses of miracidia of Schistosoma mansoni and other trematodes to substances emitted by snails. J. Parasitol. 1970, 56, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.M.; Stibbs, H.H.; Chernin, E.; Ward, S. A simple quantitative technique for testing behavioral responses of Schistosoma mansoni miracidia to chemicals. J. Parasitol. 1978, 64, 277–282. [Google Scholar] [CrossRef]
- Anderson, R.M.; Mercer, J.G.; Wilson, R.A.; Carter, N.P. Transmission of Schistosoma mansoni from man to snail: Experimental studies of miracidial survival and infectivity in relation to larval age, water temperature, host size and host age. J. Parasitol. 1982, 85, 339–360. [Google Scholar] [CrossRef]
- Dajem, S.M.B.; Mostafa, O.M. Scanning electron microscopical studies on Schistosoma mansoni cercariae exposed to ultraviolet irradiation. Aust. J. Basic Appl. Sci. 2007, 1, 776–784. [Google Scholar]
- El-Gindy, M.S.; Radhawy, I.A. Effect of low concentrations of sodium pentachlorophenate on the fecundity and egg viability of Bulinus truncatus from central Iraq. Bull. Endem. Dis. 1965, 7, 44–54. [Google Scholar]
- Frick, L.P.; De Jimenez, W.Q. Egg clutches as against individual eggs of Australorbis glabratus as test units in molluscicide evaluation. Bull. WHO 1963, 29, 286. [Google Scholar]
- Siqueira-Neto, J.L.; Debnath, A.; McCall, L.-I.; Bernatchez, J.A.; Ndao, M.; Reed, S.L.; Rosenthal, P.J. Cysteine proteases in protozoan parasites. PLoSNegl Trop. Dis. 2018, 12, e0006512. [Google Scholar] [CrossRef] [Green Version]
- El Ridi, R.; Tallima, H.; Selim, S.; Donnelly, S.; Cotton, S.; Santana, B.G.; Dalton, J.P. Cysteine Peptidases as Schistosomiasis Vaccines with Inbuilt Adjuvanticity. PLoS ONE 2014, 9, e85401. [Google Scholar] [CrossRef] [Green Version]
- Wright, C.W. Recent developments in naturally derived antimalarials: Cryptolepine analogues. J. Pharm. Pharmacol. 2007, 59, 899–904. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.M.; Abdel-Tawab, H. Cystoseira barbata marine algae have a molluscicidal activity against Biomphalaria alexandrina snails supported by scanning electron microscopy, hematological and histopathological alterations, and larvicidal activity against the infective stages of Schistosoma mansoni. Biologia 2020, 75, 1945–1954. [Google Scholar]
- Rizk, E.S. Schistosomiasis control: Evaluation of the molluscicidal activity of a plant extract (Sesbania sesban) against Biomphalaria alexandrina. J. Egypt. Ger. Socie. Zool. 1998, 27, 91–108. [Google Scholar]
- Rizk, E.S.; ElMehlawy, M.H.; Mona, M.H. Evaluation of the slow-release potency of polymeric niclosamide against Biomphalaria alexandrina eggs and Schistosoma mansoni larvae. J. Egypt. Ger. Socie. Zool. 2001, 35, 189–203. [Google Scholar]
- Santos, A.F.D.; Fonseca, S.A.; César, F.A.; Albuquerque, M.C.P.d.A.; Santana, J.V.; Santana, A.E.G. A penta-substituted pyridine alkaloid from the rhizome of Jatropha elliptica (Pohl) Muell. Arg. is active against Schistosoma mansoni and Biomphalaria glabrata. Parasitol. Res. 2014, 113, 1077–1084. [Google Scholar] [CrossRef] [Green Version]
- Njogu, M.K.; Matasyoh, J.C.; Kibor, A.C. Isolation of four furoquinoline alkaloids from Teclea nobilis and their activity against Schistosoma mansoni miracidia. J. Biomed. Pharm. Res. 2014, 3, 87–93. [Google Scholar]
- Eissa, M.M.; El Bardicy, S.; Tadros, M. Bioactivity of miltefosine against aquatic stages of Schistosoma mansoni, Schistosoma haematobium and their snail hosts, supported by scanning electron microscopy. Parasites Vectors 2011, 4, 73. [Google Scholar] [CrossRef] [Green Version]
- Manneck, T.; Haggenmüller, Y.; Keiser, J. Morphological effects and tegumental alterations induced by mefloquine on schistosomula and adult flukes of Schistosoma mansoni. Parasitology 2010, 137, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.M.; Coultas, K.A. Identification of plumbagin and sanguinarine as effective chemotherapeutic agents for treatment of schistosomiasis. Int. J. Parasitol. Drugs Drug Resist. 2013, 3, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Meshnick, S.R.; Thomas, A.; Ranz, A.; Xu, C.M.; Pan, H.Z. Artemisinin (qinghaosu): The role of intracellular hemin in its mechanism of antimalarial action. Mol. Biochem. Parasitol. 1991, 49, 181–189. [Google Scholar] [CrossRef]
- Xiao, S.H.; Keiser, J.; Xue, J.; Tanner, M.; Morson, G.; Utzinger, J. Effect of single-dose oral artemether and tribendimidine on the tegument of adult Clonorchis sinensis in rats. Parasitol. Res. 2009, 104, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Tantawy, A.A.; Sharaf El-Din, A.T.; Bakry, F.A. Molluscicidal effect of Solanum dubium (Solanaceae) against Biomphalaria alexandrina snails under laboratory conditions. Proc. Int. Conf. Biol. Sci. 2000, 1, 307–318. [Google Scholar]
- Rizk, M.Z.; Metwally, N.S.; Hamed, M.A.; Mohamed, A.M. Correlation between steroid sex hormones, egg laying capacity and cercarial shedding in Biomphalaria alexandrina snails after treatment with Haplophyllum tuberculatum. Exp. Parasitol. 2012, 132, 171–179. [Google Scholar] [CrossRef]
- Abdel Salam, H.A.; Maghrabi, O.A.; Al-Malki, E.S.; Albenasy, K.S. Thymol as Molluscicidal Agent against Biomphalaria alexandrina, Intermediate Host of Schistosoma mansoni. J. Essent. Oil-Bear. Plants 2021, 24, 1439–1450. [Google Scholar] [CrossRef]
- Hamdi, S.A.; Rawi, S.M. Biological and biochemical studies on Biomphalaria alexandrina snails, treated with low concentrations of certain molluscicides (synthetic and of plant origin). J. Egypt. Soc. Parasitol. 2005, 35, 841–858. [Google Scholar]
- Bakry, F.A.; Sakrane, A.A.; Ismail, N.M. Molluscicidal effect of fungicide, herbicide and plant extract on some biological and physiological parameters of Biomphalaria alexandrina. J. Egypt. Soc. Parasitol. 2002, 32, 821–835. [Google Scholar]
- Demetillo, M.T.; Baguio, M.L.; Limitares, D.E.; Madjos, G.G.; Abrenica-Adamat, L.R. Effect of Cymbopogon citratus (lemon grass) crude leaf extracts on the developmental stages of Pomacea canaliculata (Golden apple snail). Adv. Environ. Sci. 2015, 3, 460–467. [Google Scholar]
- Mostafa, S.S.; Gawish, F.A. Towards to control Biomphalaria alexandrina snails and the free living larval stages of Schistosoma mansoni using the microalga Spirulina platensis. Austr. J. Bas. Appl. Sci. 2009, 3, 4112–4119. [Google Scholar]
- Osman, G.Y.; Mohamed, A.Z.; Sheir, K.S.; Hassab, E.L.; Nabi, S.E.; Allam, A.S. Molluscidal activity of Mirazid on Biomphalaria alexandrina snails: Biological and molecular studies. Int. J. Adv. Res. Publ. 2014, 2, 977–989. [Google Scholar]
- Fatimatuzzahra, F.; Putriana, A.; Jumadi, J.; Lestari, D.F.; Anggoro, B.S. The Comparison between Tree Sorrel and Snake Plant on the Hatchability Inhibition of Apple Snail Eggs. Biosfer Jurnal Tadris Biologi 2020, 11, 43–50. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PDB Code | Compound | S | RMSD | Amino Acids and Type of Bonding |
|---|---|---|---|---|
| 2P7U | CAAQ | −4.8697 | 1.8618 | ASP117 (H-donor), ASP117 (ionic) |
| AEAN | −5.9386 | 1.4522 | MET68 (H-donor), MET68 (H-donor), CYS25 (H-donor), ASP161 (H-donor), ALA208 (Pi-H) | |
| APAN | −5.6194 | 1.7254 | ASP117 (H-donor), ASP117 (ionic), LEU67 (Pi-H) | |
| AEAA | −5.2061 | 2.0628 | CYS25 (H-donor), GLY66 (H-donor), MET68 (H-donor) | |
| APAA | −5.4576 | 1.5371 | ASP117 (H-donor), ASP117 (ionic) | |
| APACQ | −5.2330 | 1.7619 | ASP117 (H-donor), ASP117 (ionic), LEU67 (Pi-H) | |
| D1R | −5.678 | 1.4811 | GLY66 (H-donor), GLY66 (H-acceptor), ASP161 (H-donor), GLN19 (H-acceptor), TRP184 (H-acceptor) |
| Time | Exposure | Non-Infected Snails | Infected Snails |
|---|---|---|---|
| 24 h | Control | 11.0 ± 0.05 | 0.18 ± 0.08 |
| 0.25 ppm | 0.04 ± 0.05 | 0.08 ± 0.02 | |
| 1 ppm | 0.07± 0.09 | 0.18 ± 0.02 | |
| 1st week | Control | 2.80 ± 0.38 | 1.97 ± 0.46 |
| 0.25 ppm | 0.16 ± 0.02 * | 0.16 ± 0.02 * | |
| 1 ppm | 0.4 ± 0.28 * | 0.09 ± 0.08 * | |
| 2nd week | Control | 2.67 ± 0.33 | 1.96 ± 0.47 |
| 0.25 ppm | 0.17 ± 0.04 * | 0.18 ± 0.05 * | |
| 1 ppm | 0.06 0.05 * | 0.09 ± 0.12 * |
| Time | Treatments | Non-Infected Snails | Infected Snails | ||||
|---|---|---|---|---|---|---|---|
| Normal | Anucleated | Multi-Nucleated | Normal | Anucleated | Multi-Nucleated | ||
| 24 h | control | 94 | 1 | 1 | 84 | 5.2 | 15 |
| 0.25 ppm | 40.4 | 200 | 46 | 42 | 72 | 100 | |
| 1 ppm | 79.4 | 0 | 0 | 73.7 | 90 | 0 | |
| 1st week | control | 89 | 7.7 | 3.4 | 86.4 | 9.3 | 4.3 |
| 0.25 ppm | 4 | 7 | 0 | 5 | 17 | 0 | |
| 1 ppm | 18 | 12 | 8 | 3 | 4 | 2 | |
| 2nd week | control | 96.3 | 2.2 | 1.5 | 95.5 | 2.6 | 2 |
| 0.25 ppm | 5 | 3 | 0 | 6 | 4 | 1 | |
| 1 ppm | 2 | 4 | 9 | 4 | 17 | 0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheir, S.K.; Elmongy, E.I.; Mohamad, A.H.; Osman, G.Y.; Bendary, S.E.; Ahmed, A.A.S.; Binsuwaidan, R.; El-Sayed, I.E.-T. Molluscicidal and Larvicidal Potency of N-Heterocylic Analogs against Biomophalaria alexandrina Snails and Schistosoma mansoni Larval Stages. Pharmaceutics 2023, 15, 1200. https://doi.org/10.3390/pharmaceutics15041200
Sheir SK, Elmongy EI, Mohamad AH, Osman GY, Bendary SE, Ahmed AAS, Binsuwaidan R, El-Sayed IE-T. Molluscicidal and Larvicidal Potency of N-Heterocylic Analogs against Biomophalaria alexandrina Snails and Schistosoma mansoni Larval Stages. Pharmaceutics. 2023; 15(4):1200. https://doi.org/10.3390/pharmaceutics15041200
Chicago/Turabian StyleSheir, Sherin K., Elshaymaa I. Elmongy, Azza H. Mohamad, Gamalat Y. Osman, Shimaa E. Bendary, Abdullah A. S. Ahmed, Reem Binsuwaidan, and Ibrahim El-Tantawy El-Sayed. 2023. "Molluscicidal and Larvicidal Potency of N-Heterocylic Analogs against Biomophalaria alexandrina Snails and Schistosoma mansoni Larval Stages" Pharmaceutics 15, no. 4: 1200. https://doi.org/10.3390/pharmaceutics15041200
APA StyleSheir, S. K., Elmongy, E. I., Mohamad, A. H., Osman, G. Y., Bendary, S. E., Ahmed, A. A. S., Binsuwaidan, R., & El-Sayed, I. E. -T. (2023). Molluscicidal and Larvicidal Potency of N-Heterocylic Analogs against Biomophalaria alexandrina Snails and Schistosoma mansoni Larval Stages. Pharmaceutics, 15(4), 1200. https://doi.org/10.3390/pharmaceutics15041200