

Emergence of Nontuberculous Mycobacteria at the Human–Livestock–Environment Interface in Zambia
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
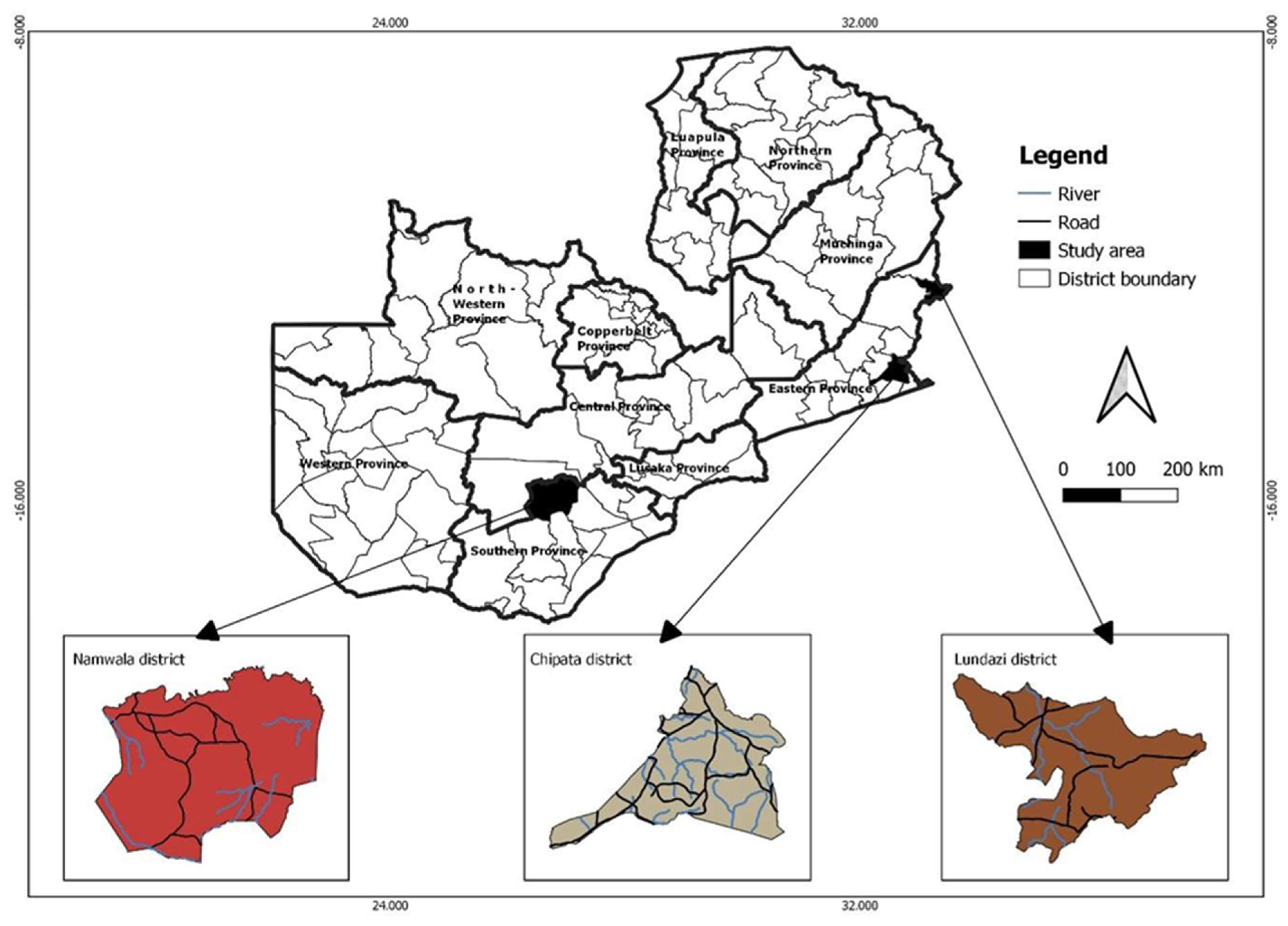
2.1. Study Site and Design
2.2. Participants Recruitment and Sample Collection
2.2.1. Human Sputum Samples
2.2.2. Cattle Tissues
2.2.3. Water Samples
2.3. Sample Processing
2.3.1. Sputum Samples
2.3.2. Tissue Sample
2.3.3. Water Sample Processing
2.4. Identification of NTM
2.5. DNA Extraction and Amplification
2.6. Purification of the PCR Products and Cycle Sequencing
2.7. Data Analysis
3. Results
3.1. Prevalence of Nontuberculous Mycobacteria in Humans
3.2. Prevalence of NTM in Cattle
3.3. Prevalence of NTM in Water
3.4. Overall Prevalence of NTM in Humans, Cattle, and Water
3.5. NTM Species Distribution at the Human–Livestock–Environment Interface
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmed, I.; Tiberi, S.; Farooqi, J.; Jabeen, K.; Yeboah-Manu, D.; Migliori, G.B.; Hasan, R. Non-tuberculous mycobacterial infections—A neglected and emerging problem. Int. J. Infect. Dis. 2020, 92, S46–S50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Roa, P.; Aznar, E.; Cacho, J.; Cogollos-Agruña, R.; Domingo, D.; García-Arata, M.; Millán, R.; Páez, M.; Perez-Cecilia, E.; Ruiz-Serrano, M.; et al. Epidemiology of Non-Tuberculous Mycobacteria isolated from clinical specimens in Madrid, Spain, from 2013 to 2017. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1089–1094. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.L.; Sarmiento, M.E.; Alvarez-Cabrera, N.; Norazmi, M.N.; Acosta, A. Pulmonary non-tuberculous mycobacterial infections: Current state and future management. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 39, 799–826. [Google Scholar] [CrossRef]
- Ratnatunga, C.N.; Lutzky, V.P.; Kupz, A.; Doolan, D.L.; Reid, D.W.; Field, M.; Bell, S.C.; Thomson, R.M.; Miles, J.J. The Rise of Non-Tuberculosis Mycobacterial Lung Disease. Front. Immunol. 2020, 11, 303. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.M.; Odell, J.A. Nontuberculous mycobacterial pulmonary infections. J. Thorac. Dis. 2014, 6, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Falkinham, J.O. Environmental Sources of Nontuberculous Mycobacteria. Clin. Chest Med. 2015, 36, 35–41. [Google Scholar] [CrossRef]
- Kankya, C.; Muwonge, A.; Djønne, B.; Munyeme, M.; Opuda-Asibo, J.; Skjerve, E.; Oloya, J.; Edvardsen, V.; Johansen, T.B. Isolation of non-tuberculous mycobacteria from pastoral ecosystems of Uganda: Public Health significance. BMC Public Health 2011, 11, 320. [Google Scholar] [CrossRef] [Green Version]
- Ahn, K.; Kim, Y.K.; Hwang, G.Y.; Cho, H.; Uh, Y. Continued Upward Trend in Non-Tuberculous Mycobacteria Isolation over 13 Years in a Tertiary Care Hospital in Korea. Yonsei Med. J. 2021, 62, 903–910. [Google Scholar] [CrossRef]
- Stout, J.E.; Koh, W.-J.; Yew, W.W. Update on pulmonary disease due to non-tuberculous mycobacteria. Int. J. Infect. Dis. 2016, 45, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Halstrom, S.; Price, P.; Thomson, R. Review: Environmental mycobacteria as a cause of human infection. Int. J. Mycobacteriology 2015, 4, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Briancesco, R.; Semproni, M.; Paradiso, R.; Bonadonna, L. Nontuberculous mycobacteria: An emerging risk in engineered environmental habitats. Ann. Microbiol. 2013, 64, 735–740. [Google Scholar] [CrossRef]
- Xu, J.; Li, P.; Zheng, S.; Shu, W.; Pang, Y. Prevalence and risk factors of pulmonary nontuberculous mycobacterial infections in the Zhejiang Province of China. Epidemiology Infect. 2019, 147, e269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prevots, D.R.; Marras, T.K. Epidemiology of Human Pulmonary Infection with Nontuberculous Mycobacteria. Clin. Chest Med. 2014, 36, 13–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agizew, T.; Basotli, J.; Alexander, H.; Boyd, R.; Letsibogo, G.; Auld, A.; Nyirenda, S.; Tedla, Z.; Mathoma, A.; Mathebula, U.; et al. Higher-than-expected prevalence of non-tuberculous mycobacteria in HIV setting in Botswana: Implications for diagnostic algorithms using Xpert MTB/RIF assay. PLoS ONE 2017, 12, e0189981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliyu, G.; El-Kamary, S.S.; Abimiku, A.; Brown, C.; Tracy, K.; Hungerford, L.; Blattner, W. Prevalence of Non-Tuberculous Mycobacterial Infections among Tuberculosis Suspects in Nigeria. PLoS ONE 2013, 8, e63170. [Google Scholar] [CrossRef] [Green Version]
- Bjerrum, S.; Oliver-Commey, J.; Kenu, E.; Lartey, M.; Newman, M.J.; Addo, K.K.; Hilleman, D.; Andersen, A.B.; Johansen, I.S. Tuberculosis and non-tuberculous mycobacteria among HIV -infected individuals in Ghana. Trop. Med. Int. Health 2016, 21, 1181–1190. [Google Scholar] [CrossRef]
- Maiga, M.; Siddiqui, S.; Diallo, S.; Diarra, B.; Traoré, B.; Shea, Y.R.; Zelazny, A.M.; Dembele, B.P.P.; Goita, D.; Kassambara, H.; et al. Failure to Recognize Nontuberculous Mycobacteria Leads to Misdiagnosis of Chronic Pulmonary Tuberculosis. PLoS ONE 2012, 7, e36902. [Google Scholar] [CrossRef]
- Wagner, D.; Lipman, M.; Cooray, S.; Ringshausen, F.C.; Morimoto, K.; Koh, W.; Thomson, R. Nontuberculous Mycobacterial Disease; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Hoza, A.S.; Mfinanga, S.G.M.; Rodloff, A.C.; Moser, I.; König, B. Increased isolation of nontuberculous mycobacteria among TB suspects in Northeastern, Tanzania: Public health and diagnostic implications for control programmes. BMC Res. Notes 2016, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Pokam, B.T.; Yeboah-Manu, D.; Ofori, S.; Guemdjom, P.; Teyim, P.; Lawson, L.; Amiteye, D.; Yhiler, N.; Djuikoue, I.; Asuquo, A. Prevalence of non-tuberculous mycobacteria among previously treated TB patients in the Gulf of Guinea, Africa. IJID Reg. 2022, 3, 287–292. [Google Scholar] [CrossRef]
- Santos, A.; Carneiro, S.; Silva, A.; Gomes, J.; Macedo, R. Nontuberculous Mycobacteria in Portugal: Trends from the last decade. Pulmonology 2022. [Google Scholar] [CrossRef]
- Monteiro, N.F.; Peres, S.; Mansinho, K. Non-Tuberculous Mycobacteria: Seven-Year Experience of a Tertiary Hospital. Acta Méd. Port. 2019, 32, 208–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.; Malhotra, B.; Tiwari, J.; Bhargava, S. Profile of Nontuberculous Mycobacteria in Patients Suspected of Tuberculosis and Drug-Resistant Tuberculosis. J. Lab. Physicians 2020, 12, 203–211. [Google Scholar] [CrossRef]
- Griffith, D.E.; Aksamit, T.; Brown-Elliott, B.A.; Catanzaro, A.; Daley, C.; Gordin, F.; Holland, S.M.; Horsburgh, R.; Huitt, G.; Iademarco, M.F.; et al. An Official ATS/IDSA Statement: Diagnosis, Treatment, and Prevention of Nontuberculous Mycobacterial Diseases. Am. J. Respir. Crit. Care Med. 2007, 175, 367–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busatto, C.; Vianna, J.S.; Silva, A.B.S.; Basso, R.; Silveira, J.; Von Groll, A.; Ramis, I.B.; Da Silva, P.E.A. Nontuberculous mycobacteria in patients with suspected tuberculosis and the genetic diversity of Mycobacterium avium in the extreme south of Brazil. J. Bras. de Pneumol. 2020, 46, e20190184. [Google Scholar] [CrossRef] [PubMed]
- Zulu, M.; Monde, N.; Nkhoma, P.; Malama, S.; Munyeme, M. Nontuberculous Mycobacteria in Humans, Animals, and Water in Zambia: A Systematic Review. Front. Trop. Dis. 2021, 2. [Google Scholar] [CrossRef]
- Philley, J.V.; Griffith, D.E. Treatment of slowly growing mycobacteria. Clin. Chest Med. 2015, 36, 79–90. [Google Scholar] [CrossRef]
- Cheng, L.-P.; Chen, S.-H.; Lou, H.; Gui, X.-W.; Shen, X.-N.; Cao, J.; Sha, W.; Sun, Q. Factors Associated with Treatment Outcome in Patients with Nontuberculous Mycobacterial Pulmonary Disease: A Large Population-Based Retrospective Cohort Study in Shanghai. Trop. Med. Infect. Dis. 2022, 7, 27. [Google Scholar] [CrossRef]
- Lubungu, M.; Mofya-mukuka, R. The Status of the Smallholder Livestock Sector in Zambia; IAPRI: Lusaka, Zambia, 2012. [Google Scholar]
- Monde, N.; Munyeme, M.; Chongwe, G.; Wensman, J.J.; Zulu, M.; Siziya, S.; Tembo, R.; Siame, K.K.; Shambaba, O.; Malama, S. First and Second-Line Anti-Tuberculosis Drug-Resistance Patterns in Pulmonary Tuberculosis Patients in Zambia. Antibiotics 2023, 12, 166. [Google Scholar] [CrossRef]
- Kumar, A.; Kebede, E.; Kassaye, E. Evaluation of quality of beef produced and sold in parts of Tigray Region of Ethiopia. Trop. Anim. Health Prod. 2009, 42, 445–449. [Google Scholar] [CrossRef]
- Monde, N.; Munyeme, M.; Muwonge, A.; Muma, J.B.; Malama, S. Characterization of non-tuberculous mycobacterium from humans and water in an Agropastoral area in Zambia. BMC Infect. Dis. 2018, 18, 20. [Google Scholar] [CrossRef] [Green Version]
- Katale, B.; Mbugi, E.; Botha, L.; Keyyu, J.; Kendall, S.; Dockrell, H.; Michel, A.; Kazwala, R.; Rweyemamu, M.; van Helden, P.; et al. Species diversity of non-tuberculous mycobacteria isolated from humans, livestock and wildlife in the Serengeti ecosystem, Tanzania. BMC Infect. Dis. 2014, 14, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addo, K.K.; Addo, S.O.; Mensah, G.I.; Mosi, L.; Bonsu, F.A. Genotyping and drug susceptibility testing of mycobacterial isolates from population-based tuberculosis prevalence survey in Ghana. BMC Infect. Dis. 2017, 17, 743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, M.; San, K.C.; Pho, Y.; Sok, C.; Dousset, J.-P.; Brant, W.; Hurtado, N.; Eam, K.K.; Ardizzoni, E.; Heng, S.; et al. Nontuberculous Mycobacteria Infections at a Provincial Reference Hospital, Cambodia. Emerg. Infect. Dis. 2017, 23, 1139–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kone, B.; Sarro, Y.S.; Maiga, M.; Sanogo, M.; Somboro, A.M.; Diarra, B.; Togo, A.C.G.; Coulibaly, N.; Dembele, B.P.P.; Goita, D.; et al. Clinical characteristics of non-tuberculous mycobacterial pulmonary infections in Bamako, Mali. Epidemiol. Infect. 2018, 146, 354–358. [Google Scholar] [CrossRef] [Green Version]
- Okoi, C.; Anderson, S.T.B.; Antonio, M.; Mulwa, S.N.; Gehre, F.; Adetifa, I.M.O. Non-tuberculous Mycobacteria isolated from Pulmonary samples in sub-Saharan Africa—A Systematic Review and Meta Analyses. Sci. Rep. 2017, 7, 12002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monde, N.; Munyeme, M.; Muwonge, A.; Muma, J.B.; Malama, S. Non-Tuberculous Mycobacteria Recovered from Suspected TB Cases in Namwala District of Zambia. ARC J. Clin. Case Rep. 2017, 3, 4–10. [Google Scholar] [CrossRef]
- Buijtels, P.C.A.M.; Van Der Sande, M.A.B.; Parkinson, S.; Verbrugh, H.A.; Petit, P.L.C.; van Soolingen, D. Isolation of non-tuberculous mycobacteria at three rural settings in Zambia; a pilot study. Clin. Microbiol. Infect. 2010, 16, 1142–1148. [Google Scholar] [CrossRef] [Green Version]
- Chanda-Kapata, P.; Kapata, N.; Klinkenberg, E.; Mulenga, L.; Tembo, M.; Katemangwe, P.; Sunkutu, V.; Mwaba, P.; Grobusch, M.P. Non-tuberculous mycobacteria (NTM) in Zambia: Prevalence, clinical, radiological and microbiological characteristics. BMC Infect. Dis. 2015, 15, 500. [Google Scholar] [CrossRef] [Green Version]
- Shafipour, M.; Shirzad-Aski, H.; Ghaemi, E.A.; Sohrabi, A.; Taziki, M.; Kochkaksaraei, M.B.; Rahimi, S. Occurrence and risk factors of nontuberculous mycobacteria in tuberculosis-suspected patients in the north of Iran. Iran. J. Microbiol. 2021. [Google Scholar] [CrossRef]
- Shahraki, A.H.; Heidarieh, P.; Bostanabad, S.Z.; Khosravi, A.D.; Hashemzadeh, M.; Khandan, S.; Biranvand, M.; Schraufnagel, D.; Mirsaeidi, M. “Multidrug-resistant tuberculosis” may be nontuberculous mycobacteria. Eur. J. Intern. Med. 2015, 26, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Faverio, P.; De Giacomi, F.; Bodini, B.D.; Stainer, A.; Fumagalli, A.; Bini, F.; Luppi, F.; Aliberti, S. Nontuberculous mycobacterial pulmonary disease: An integrated approach beyond antibiotics. ERJ Open Res. 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Gopalaswamy, R.; Shanmugam, S.; Mondal, R.; Subbian, S. Of tuberculosis and non-tuberculous mycobacterial infections—A comparative analysis of epidemiology, diagnosis and treatment. J. Biomed. Sci. 2020, 27, 74. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-J.; Li, Y.-X.; Zhao, Y.; Yang, W.-H.; Xiao, M.; Kudinha, T.; Xu, Y.-C. Prevalence of nontuberculous mycobacteria in a tertiary hospital in Beijing, China, January 2013 to December 2018. BMC Microbiol. 2020, 20, 158. [Google Scholar] [CrossRef] [PubMed]
- Szturmowicz, M.; Siemion-Szcześniak, I.; Wyrostkiewicz, D.; Klatt, M.; Brzezińska, S.; Zabost, A.; Lewandowska, A.; Filipczak, D.; Oniszh, K.; Skoczylas, A.; et al. Factors Predisposing to Non-Tuberculous Mycobacterial Lung Disease in the Patients with Respiratory Isolates of Non-Tuberculous Mycobacteria. Adv. Respir. Med. 2018, 86, 261–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donohue, M.J. Epidemiological risk factors and the geographical distribution of eight Mycobacterium species. BMC Infect. Dis. 2021, 21, 258. [Google Scholar] [CrossRef] [PubMed]
- Mbeha, B.; Mine, M.; Motswaledi, M.S.; Dewar, J.; Study, T. Nontuberculous Mycobacteria, Botswana, 2011–2014. Emerg. Infect. Dis. 2019, 25, 1401–1403. [Google Scholar] [CrossRef] [PubMed]
- Saptawati, L.; Mashuri, Y.A.; Suryawati, B.; Harsono, H.; Pradiptakirana, R. Risk Factors and Prognosis of Nontuberculous Mycobacteria Infection in a High Prevalence of Tuberculosis Setting. J. Epidemiol. Public Health 2020, 5, 79–87. [Google Scholar] [CrossRef]
- Moore, J.; Kruijshaar, M.E.; Ormerod, L.P.; Drobniewski, F.; Abubakar, I. Increasing reports of non-tuberculous mycobacteria in England, Wales and Northern Ireland, 1995–2006. BMC Public Health 2010, 10, 612. [Google Scholar] [CrossRef]
- Karamat, A.; Ambreen, A.; Ishtiaq, A.; Tahseen, S.; Rahman, M.A.; Mustafa, T. Isolation of non-tuberculous mycobacteria among tuberculosis patients, a study from a tertiary care hospital in Lahore, Pakistan. BMC Infect. Dis. 2021, 21, 381. [Google Scholar] [CrossRef]
- Henkle, E.; Hedberg, K.; Schafer, S.; Novosad, S.; Winthrop, K.L. Population-based Incidence of Pulmonary Nontuberculous Mycobacterial Disease in Oregon 2007 to 2012. Ann. Am. Thorac. Soc. 2015, 12, 642–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, C.-C.; Wu, M.-F.; Pan, S.-W.; Wu, T.-S.; Lai, H.-C.; Lin, M.-C. Host immune response against environmental nontuberculous mycobacteria and the risk populations of nontuberculous mycobacterial lung disease. J. Formos. Med Assoc. 2020, 119, S13–S22. [Google Scholar] [CrossRef] [PubMed]
- Slathia, P.; Narang, D.; Chandra, M. Detection of Nontuberculous Mycobacterial Species from Tissue Samples of Cattle and Buffaloes by PCR and PRA (PCR-RFLP). Indian J. Anim. Res. 2021. [Google Scholar] [CrossRef]
- Katale, B.Z.; Mbugi, E.V.; Karimuribo, E.D.; Keyyu, J.D.; Kendall, S.; Kibiki, G.S.; Godfrey-Faussett, P.; Michel, A.L.; Kazwala, R.R.; van Helden, P.; et al. Prevalence and risk factors for infection of bovine tuberculosis in indigenous cattle in the Serengeti ecosystem, Tanzania. BMC Vet. Res. 2013, 9, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ntivuguruzwa, J.B.; Michel, A.L.; Kolo, F.B.; Mwikarago, I.E.; Ngabonziza, J.C.S.; van Heerden, H. Prevalence of bovine tuberculosis and characterization of the members of the Mycobacterium tuberculosis complex from slaughtered cattle in Rwanda. PLoS Neglected Trop. Dis. 2022, 16, e0009964. [Google Scholar] [CrossRef]
- Tingan, T.K.; Mensah, G.I.; Agyekum, E.B.; Amanor, I.B.; Addo, S.O.; Ayamdoo, Y.I.; Duah, M.S.; Mosi, L.; Addo, K.K. Non-tuberculous mycobacteria, not Mycobacterium bovis, are a significant cause of TB-like lesions observed in slaughtered cattle in Ghana. IJID Reg. 2022, 3, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Jarguín, A.; Martínez-Burnes, J.; Molina-Salinas, G. Isolation Nontuberculous Mycobacteria and Histopathological Changes in Lymph Nodes Collected at the Abattoir in Cattle Reactive-Positive to Tuberculin Dermal Test. 2020, pp. 1–17. Available online: https://www.researchsquare.com/article/rs-15889/latest.pdf (accessed on 25 August 2022).
- Akinseye, V.O.; Adebayo, M.D.; Genesis, O.O.; Adelakun, O.D.; Cadmus, S.I.B. Prevalence and risk factors of mycobacterial infections in farm and trade cattle in southwestern Nigeria. Trop. Anim. Health Prod. 2017, 50, 761–772. [Google Scholar] [CrossRef]
- Nuru, A.; Zewude, A.; Mohammed, T.; Wondale, B.; Teshome, L.; Getahun, M.; Mamo, G.; Medhin, G.; Pieper, R.; Ameni, G. Nontuberculosis mycobacteria are the major causes of tuberculosis like lesions in cattle slaughtered at Bahir Dar Abattoir, northwestern Ethiopia. BMC Veter-Res. 2017, 13, 237. [Google Scholar] [CrossRef] [Green Version]
- Chaabila, K.S. Socio-economic transformation Among traditional Cattle Keepers in Kafue Flood Plain in Namwala District, Zambia. Agric. JABS 2012, 2, 90. [Google Scholar]
- Malama, S.; Munyeme, M.; Mwanza, S.; Muma, J.B. Isolation and characterization of non tuberculous mycobacteria from humans and animals in Namwala District of Zambia. BMC Res. Notes 2014, 7, 622. [Google Scholar] [CrossRef] [Green Version]
- Munyeme, M.; Muma, J.B.; Samui, K.L.; Skjerve, E.; Nambota, A.M.; Phiri, I.G.K.; Rigouts, L.; Tryland, M. Prevalence of bovine tuberculosis and animal level risk factors for indigenous cattle under different grazing strategies in the livestock/wildlife interface areas of Zambia. Trop. Anim. Health Prod. 2008, 41, 345–352. [Google Scholar] [CrossRef]
- Muwonge, A.; Kankya, C.; Godfroid, J.; Djonne, B.; Opuda-Asibo, J.; Biffa, D.; Ayanaw, T.; Munyeme, M.; Skjerve, E. Prevalence and associated risk factors of mycobacterial infections in slaughter pigs from Mubende district in Uganda. Trop. Anim. Health Prod. 2009, 42, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Kanyala, E.; Shuaib, Y.A.; Schwarz, N.G.; Andres, S.; Richter, E.; Sawadogo, B.; Sawadogo, M.; Germaine, M.; Lassina, O.; Poppert, S.; et al. Prevalence and Molecular Characterization of Mycobacterium bovis in Slaughtered Cattle Carcasses in Burkina Faso; West Africa. Microorganisms 2022, 10, 1378. [Google Scholar] [CrossRef]
- Kazwala, R.; Kambarage, D.; Daborn, C.; Nyange, J.; Jiwa, S.; Sharp, J. Risk factors associated with the occurrence of bovine tuberculosis in cattle in the Southern Highlands of Tanzania. Vet. Res. Commun. 2001, 25, 609–614. [Google Scholar] [CrossRef]
- Dejene, S.W.; Heitkönig, I.M.A.; Prins, H.H.T.; Lemma, F.A.; Mekonnen, D.A.; Alemu, Z.E.; Kelkay, T.Z.; de Boer, W.F. Risk Factors for Bovine Tuberculosis (bTB) in Cattle in Ethiopia. PLoS ONE 2016, 11, e0159083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kankya, C.; Muwonge, A.; Olet, S.; Munyeme, M.; Biffa, D.; Opuda-Asibo, J.; Skjerve, E.; Oloya, J. Factors associated with pastoral community knowledge and occurrence of mycobacterial infections in Human-Animal Interface areas of Nakasongola and Mubende districts, Uganda. BMC Public Health 2010, 10, 471. [Google Scholar] [CrossRef] [Green Version]
- Rahbar, M.; Lamei, A.; Babazadeh, H.; Yavari, S.A. Isolation of rapid growing mycobacteria from soil and water in Iran. African J. Biotechnol. 2010, 9, 3618–3621. [Google Scholar]
- Velayati, A.A.; Farnia, P.; Mozafari, M.; Malekshahian, D.; Seif, S.; Rahideh, S.; Mirsaeidi, M. Molecular Epidemiology of Nontuberculous Mycobacteria Isolates from Clinical and Environmental Sources of a Metropolitan City. PLoS ONE 2014, 9, e114428. [Google Scholar] [CrossRef] [Green Version]
- Perez-Martinez, I.; Aguilar-Ayala, D.A.; Fernandez-Rendon, E.; Carrillo-Sanchez, A.K.; Helguera-Repetto, A.C.; Rivera-Gutierrez, S.; Estrada-García, T.; Cerna-Cortes, J.F.; Gonzalez-Y-Merchand, J.A. Occurrence of potentially pathogenic nontuberculous mycobacteria in Mexican household potable water: A pilot study. BMC Res. Notes 2013, 6, 531. [Google Scholar] [CrossRef] [Green Version]
- Falkinham, J.O.; Rahbar, M.; Lamei, A.; Babazadeh, H.; Yavari, S.A.; Delghandi, M.R. Ecology of nontuberculous mycobacteria-where do human infections come from? PLoS ONE 2013, 9, 95–102. [Google Scholar]
- Falkinham, J.O. Reducing Exposure to Nontuberculous Mycobacteria. NTM-TB Insights 2015, 2015, 1–15. [Google Scholar]
- Gan, Y.; Rahmatika, I.; Kurisu, F.; Furumai, H.; Simazaki, D.; Fukano, H.; Hoshino, Y.; Kasuga, I. The fate and risk of nontuberculous mycobacteria in the water supply system: A review. H2Open J. 2022, 5, 180–197. [Google Scholar] [CrossRef]
- Chin’Ombe, N.; Muzividzi, B.; Ruhanya, V.; Nziramasanga, P. 16S-23S Ribosomal RNA Internal Transcribed Spacer Region-based Genetic Analysis of Nontuberculous Mycobacteria from Zimbabwe. Acta Sci. Med. Sci. 2021, 4, 64–69. [Google Scholar] [CrossRef]
- Dahl, V.N.; Mølhave, M.; Fløe, A.; van Ingen, J.; Schön, T.; Lillebaek, T.; Andersen, A.B.; Wejse, C. Global trends of pulmonary infections with nontuberculous mycobacteria: A systematic review. Int. J. Infect. Dis. 2022, 125, 120–131. [Google Scholar] [CrossRef]
- Tan, Y.; Su, B.; Shu, W.; Cai, X.; Kuang, S.; Kuang, H.; Liu, J.; Pang, Y. Epidemiology of pulmonary disease due to nontuberculous mycobacteria in Southern China, 2013–2016. BMC Pulm. Med. 2018, 18, 168. [Google Scholar] [CrossRef]
- Weygaerde, Y.V.; Cardinaels, N.; Bomans, P.; Chin, T.; Boelens, J.; André, E.; Van Braeckel, E.; Lorent, N. Clinical relevance of pulmonary non-tuberculous mycobacterial isolates in three reference centres in Belgium: A multicentre retrospective analysis. BMC Infect. Dis. 2019, 19, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mwangi, Z.M.; Mukiri, N.; Onyambu, F.G.; Bulimo, W. Genetic Diversity of Non-Tuberculous Mycobacteria among Symptomatic TB negative Patients in Kenya. Int. J. Mycobacteriol. 2022, 11, 60–69. [Google Scholar] [CrossRef]
- Gharbi, R.; Mhenni, B.; Ben Fraj, S.; Mardassi, H. Nontuberculous mycobacteria isolated from specimens of pulmonary tuberculosis suspects, Northern Tunisia: 2002–2016. BMC Infect. Dis. 2019, 19, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Furlaneto, I.P.; da Conceição, M.L.; Conceição, E.C.; Lopes, M.L.; Rodrigues, Y.C.; Macelino, B.R.; Gomes, H.M.; Suffys, P.N.; Guimarães, R.J.D.P.S.E.; da Silva, M.G.; et al. Molecular epidemiology of mycobacteria among herds in Marajó Island, Brazil, reveals strains genetically related and potential zoonotic risk of clinical relevance. Infect. Genet. Evol. 2019, 77, 104044. [Google Scholar] [CrossRef]
- Van Halsema, C.L.; Chihota, V.N.; Gey Van Pittius, N.C.; Fielding, K.L.; Lewis, J.J.; Van Helden, P.D.; Churchyard, G.; Grant, A. Clinical Relevance of Nontuberculous Mycobacteria Isolated from Sputum in a Gold Mining Workforce in South Africa: An Observational, Clinical Study. Biomed. Res. Int. 2015, 2015, 959107. [Google Scholar] [CrossRef] [Green Version]
- Degiacomi, G.; Sammartino, J.C.; Chiarelli, L.R.; Riabova, O.; Makarov, V.; Pasca, M.R. Mycobacterium abscessus, an Emerging and Worrisome Pathogen among Cystic Fibrosis Patients. Int. J. Mol. Sci. 2019, 20, 5868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, R.; Tolson, C.; Sidjabat, H.; Huygens, F.; Hargreaves, M. Mycobacterium abscessus isolated from municipal water—A potential source of human infection. BMC Infect. Dis. 2013, 13, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Variable | Category | Frequency | Prevalence (%) (n = 33) | 95% CI |
|---|---|---|---|---|
| Gender | Male | 421 | 19 (4.5) | 2.8–7.1 |
| Female | 421 | 14 (3.3) | 1.9–5.7 | |
| p-value | 0.899 | |||
| Gender specific | ||||
| Male | 252 | 19 (7.5) | 4.7–11.7 | |
| Female | 169 | 14 (8.3) | 4.8–13.8 | |
| p-value | 0.091 | |||
| Age (years) | ≤20 | 421 | 2 (0.5) | 0.1–1.9 |
| 21–30 | 421 | 6 (1.4) | 0.6–3.2 | |
| 31–40 | 421 | 12 (2.9) | 1.6–5.1 | |
| 41–50 | 421 | 9 (2.1) | 1.1–4.2 | |
| 41–60 | 421 | 2 (0.5) | 0.1–1.9 | |
| ≥61 | 421 | 2 (0.5) | 0.1–1.9 | |
| p-value | 0.899 | |||
| Age specific | ||||
| ≤20 | 22 | 2 (9.1) | 1.6–30.6 | |
| 21–30 | 107 | 6 (5.6) | 2.3–12.3 | |
| 31–40 | 112 | 12 (10.7) | 5.9–18.3 | |
| 41–50 | 96 | 9 (9.4) | 4.6–17.5 | |
| 41–60 | 42 | 2 (4.8) | 8.3–17.4 | |
| ≥61 | 42 | 2 (4.8) | 8.3–17.4 | |
| p-value | 0.677 | |||
| Province | Southern | 421 | 19 (4.5) | 2.8–7.1 |
| Eastern | 421 | 14 (3.3) | 1.9–5.7 | |
| p-value | 0.899 | |||
| Province specific | ||||
| Southern | 231 | 19 (8.2) | 5.5–12.7 | |
| Eastern | 190 | 14 (7.4) | 4.2–12.3 | |
| p-value | 0.091 | |||
| Districts | Namwala | 421 | 19 (4.5) | 2.8–7.1 |
| Chipata | 421 | 12 (2.9) | 1.6–5.1 | |
| Lundazi | 421 | 2 (0.5) | 0.1–1.9 | |
| p-value | 0.019 | |||
| District specific | ||||
| Namwala | 231 | 19 (8.2) | 5.5–12.7 | |
| Chipata | 109 | 12 (11.0) | 6.1–18.8 | |
| Lundazi | 81 | 2 (2.5) | 4.3–9.5 | |
| p-value | 0.022 | |||
| Overall | 33 | 7.8 | 5.5–10.9 | |
| Factors | Category | Frequency | Prevalence (%) n = 15 | 95% CI |
|---|---|---|---|---|
| Age of livestock in years | ≤5 | 142 | 0 (0.0) | 0.0–3.3 |
| 6–10 | 142 | 12 (8.5) | 4.6–14.6 | |
| ≥11 | 142 | 3 (2.1) | 0.6–6.5 | |
| p-value | 0.047 | |||
| Age specific | ||||
| ≤5 | 3 | 0 (0.0) | 0.0–69.0 | |
| 6–10 | 99 | 12 (12.1) | 6.7–20.6 | |
| ≥11 | 40 | 3 (7.5) | 2.0–21.5 | |
| p-value | 0.021 | |||
| District | Lundazi | 142 | 0 (0.0) | 0.0–3.3 |
| Chipata | 142 | 0 (0.0) | 0.0–3.3 | |
| Namwala | 142 | 15 (10.6) | 6.2–17.1 | |
| p-value | 0.007 | |||
| District specific | ||||
| Lundazi | 4 | 0 (0.0) | 0.0–60.4 | |
| Chipata | 9 | 0 (0.0) | 0.0–37.1 | |
| Namwala | 129 | 15 (11.6) | 6.9–18.8 | |
| p-value | 0.014 | |||
| Sex | Male | 142 | 10 (7.0) | 3.6–12.9 |
| Female | 142 | 5 (3.5) | 1.3–8.5 | |
| p-value | 0.001 | |||
| Sex specific | ||||
| Male | 80 | 10 (12.5) | 6.5–22.2 | |
| Female | 62 | 5 (8.1) | 3.0–18.5 | |
| p-value | 0.031 | |||
| Herd size | ≤50 | 142 | 3 (2.1) | 0.6–6.5 |
| 51–150 | 142 | 8 (5.6) | 2.6–11.2 | |
| ≥151 | 142 | 4 (2.8) | 0.9–7.5 | |
| p-value | 0.472 | |||
| Herd specific | ||||
| ≤50 | 13 | 3 (3.1) | 6.1–54.0 | |
| 51–150 | 65 | 8 (12.3) | 5.8–23.4 | |
| ≥151 | 64 | 4 (6.3) | 2.0–16.0 | |
| p-value | 0.001 | |||
| Housed | Yes | 142 | 15 (10.6) | 6.2–17.1 |
| No | 142 | 0 (0.0) | 0.00–3.3 | |
| p-value | 0.001 | |||
| Housed specific | ||||
| Yes | 138 | 15 (11.0) | 6.4–17.6 | |
| No | 4 | 0 (0.0) | 0.00–60.4 | |
| p-value | 0.001 | |||
| Flood plain | Yes | 142 | 15 (10.6) | 6.2–17.1 |
| No | 142 | 0 (0.0) | 0.0–3.3 | |
| p-value | 0.001 | |||
| Flood plain specific | ||||
| Yes | 137 | 15 (11.0) | 6.5–17.7 | |
| No | 5 | 0 (0.0) | 0.0–53.7 | |
| p-value | 0.001 | |||
| National park | Yes | 142 | 8 (5.6) | 2.6–11.2 |
| No | 142 | 7 (4.9) | 2.2–10.3 | |
| p-value | 0.611 | |||
| National park specific | ||||
| Yes | 8 | 8 (100.0) | 59.8–100.0 | |
| No | 134 | 7 (5.2) | 2.3–10.9 | |
| p-value | 0.001 | |||
| Grazing with wild animals | Yes | 142 | 8 (5.6) | 2.6–11.2 |
| No | 142 | 7 (4.9) | 2.2–10.3 | |
| p-value | 0.611 | |||
| Grazing with wild animals specific | ||||
| Yes | 87 | 8 (9.2) | 4.3–17.8 | |
| No | 55 | 7 (12.7) | 5.7–25.1 | |
| p-value | 0.019 | |||
| Overall | 15 | 10.6 | 6.2–17.1 | |
| Factors | Category | Frequency | Prevalence (%) n = 24 | 95% CI |
|---|---|---|---|---|
| Water source | Borehole | 231 | 18 (7.8) | 4.8–12.2 |
| Dam/pond/river | 231 | 2 (0.9) | 0.2–3.4 | |
| Tap | 231 | 4 (1.7) | 0.6–4.7 | |
| p-value | 0.033 | |||
| Borehole | 181 | 18 (9.9) | 6.2–15.5 | |
| Dam/pond/river | 33 | 2 (6.1) | 1.1–21.6 | |
| Tap | 17 | 4 (23.5) | 7.8–50.2 | |
| p-value | 0.021 | |||
| Purpose of water | Domestic | 231 | 19 (8.2) | 5.2–12.7 |
| Livestock | 231 | 1 (0.4) | 0.0–2.8 | |
| Both | 231 | 4 (1.7) | 0.6–4.7 | |
| p-value | 0.038 | |||
| Purpose of water specific | ||||
| Domestic | 190 | 19 (10.0) | 6.3–15.4 | |
| Livestock | 17 | 1 (5.9) | 0.3–30.8 | |
| Both | 24 | 4 (16.7) | 5.5–38.2 | |
| p-value | 0.036 | |||
| Location | Urban | 231 | 3 (1.3) | 0.3–4.1 |
| Peri-urban | 231 | 5 (2.2) | 0.8–5.3 | |
| Rural | 231 | 16 (6.9) | 4.1–11.2 | |
| p-value | 0.333 | |||
| Location specific | ||||
| Urban | 36 | 3 (8.3) | 2.2–23.6 | |
| Peri-urban | 44 | 5 (11.4) | 4.3–25.4 | |
| Rural | 151 | 16 (10.6) | 6.4–16.9 | |
| 0.899 | ||||
| Province | Eastern | 231 | 22 (9.5) | 6.2–14.2 |
| Southern | 231 | 2 (0.9) | 0.2–3.4 | |
| p-value | 0.018 | |||
| Province specific | ||||
| Eastern | 213 | 22 (10.3) | 6.7–15.4 | |
| Southern | 18 | 2 (11.1) | 1.9–36.1 | |
| p-value | 0.899 | |||
| District | Lundazi | 231 | 3 (1.3) | 0.3–4.1 |
| Chipata | 231 | 19 (8.2) | 5.2–12.7 | |
| Namwala | 231 | 2 (0.9) | 0.2–3.4 | |
| p-value | 0.016 | |||
| District specific | ||||
| Lundazi | 64 | 3 (4.7) | 1.2–13.9 | |
| Chipata | 149 | 19 (12.8) | 8.0–19.4 | |
| Namwala | 18 | 2 (11.1) | 1.9–36.1 | |
| p-value | 0.721 | |||
| Overall | 24 | 10.4 | 6.9–15.2 | |
| Variable | n | Positives | Prevalence (%) | 95% CI |
|---|---|---|---|---|
| Human | 421 | 33 | 7.8 | 5.5–10.9 |
| Cattle | 142 | 15 | 10.6 | 6.2–17.1 |
| Water | 231 | 24 | 10.4 | 6.9–15.2 |
| Overall | 794 | 72 | 9.07 | 7.21–11.34 |
| p-value | 0.106 |
| NTM Species | Source | Frequency (%) | |||
|---|---|---|---|---|---|
| Human Sputum | Cattle Tissues | Water | |||
| 1 | M. abscessus | 2 | 2 | 8.2 | |
| 2 | M. pulveris | 1 | 2.0 | ||
| 3 | M. kumamotonense | 1 | 2.0 | ||
| 4 | M. rutilum | 1 | 2 | 6.1 | |
| 5 | M. smegmatis | 1 | 2.0 | ||
| 6 | M. avium complex | 9 | 18.4 | ||
| 7 | M. fortuitum | 3 | 1 | 5 | 18.4 |
| 9 | M. boenickei | 1 | 1 | 4.1 | |
| 10 | M. littorale | 3 | 6.1 | ||
| 11 | M. parascrofulaceum | 1 | 2.0 | ||
| 12 | M. gordonae | 1 | 2 | 1 | 8.2 |
| 13 | M. phocaicum | 4 | 8.2 | ||
| 14 | M. mucogenicum | 1 | 2.0 | ||
| 15 | M. cosmeticum | 1 | 2.0 | ||
| 16 | M. species | 2 | 4.1 | ||
| 17 | M. pulveris | 1 | 2.0 | ||
| 18 | Coinfection of M. parascrofulaceum and M. europaeum | 1 | 2.0 | ||
| 19 | M. Senegalence | 1 | 2.0 | ||
| TOTAL | 28 | 4 | 17 | 100.0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zulu, M.; Malama, S.; Monde, N.; Kainga, H.; Tembo, R.; Mwaba, F.; Saad, S.A.; Daka, V.; Mukubesa, A.N.; Ndebe, J.; et al. Emergence of Nontuberculous Mycobacteria at the Human–Livestock–Environment Interface in Zambia. Microbiol. Res. 2023, 14, 430-447. https://doi.org/10.3390/microbiolres14010032
Zulu M, Malama S, Monde N, Kainga H, Tembo R, Mwaba F, Saad SA, Daka V, Mukubesa AN, Ndebe J, et al. Emergence of Nontuberculous Mycobacteria at the Human–Livestock–Environment Interface in Zambia. Microbiology Research. 2023; 14(1):430-447. https://doi.org/10.3390/microbiolres14010032
Chicago/Turabian StyleZulu, Mildred, Sydney Malama, Ngula Monde, Henson Kainga, Rabecca Tembo, Florence Mwaba, Shereen Ahmed Saad, Victor Daka, Andrew N. Mukubesa, Joseph Ndebe, and et al. 2023. "Emergence of Nontuberculous Mycobacteria at the Human–Livestock–Environment Interface in Zambia" Microbiology Research 14, no. 1: 430-447. https://doi.org/10.3390/microbiolres14010032
APA StyleZulu, M., Malama, S., Monde, N., Kainga, H., Tembo, R., Mwaba, F., Saad, S. A., Daka, V., Mukubesa, A. N., Ndebe, J., Shambaba, O., & Munyeme, M. (2023). Emergence of Nontuberculous Mycobacteria at the Human–Livestock–Environment Interface in Zambia. Microbiology Research, 14(1), 430-447. https://doi.org/10.3390/microbiolres14010032