
Clonal Micropropagation of Cymbidium erythrostylum Rolfe


Abstract
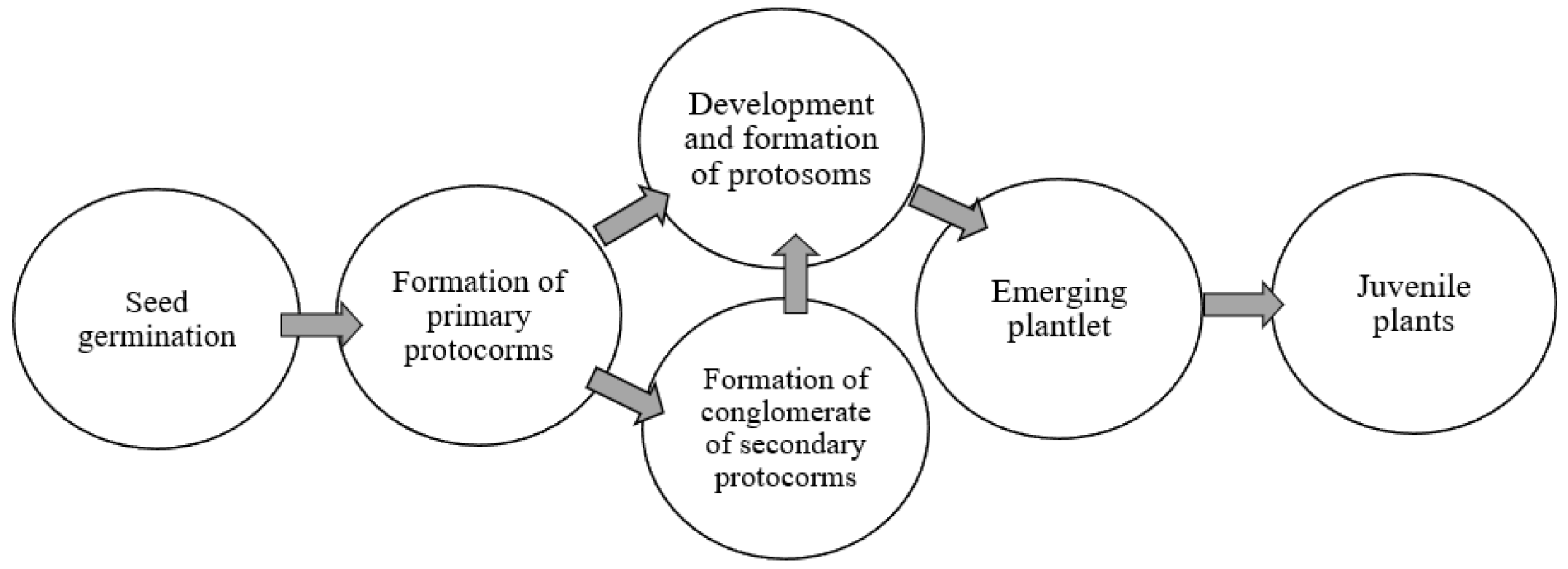
:1. Introduction
2. Materials and Methods
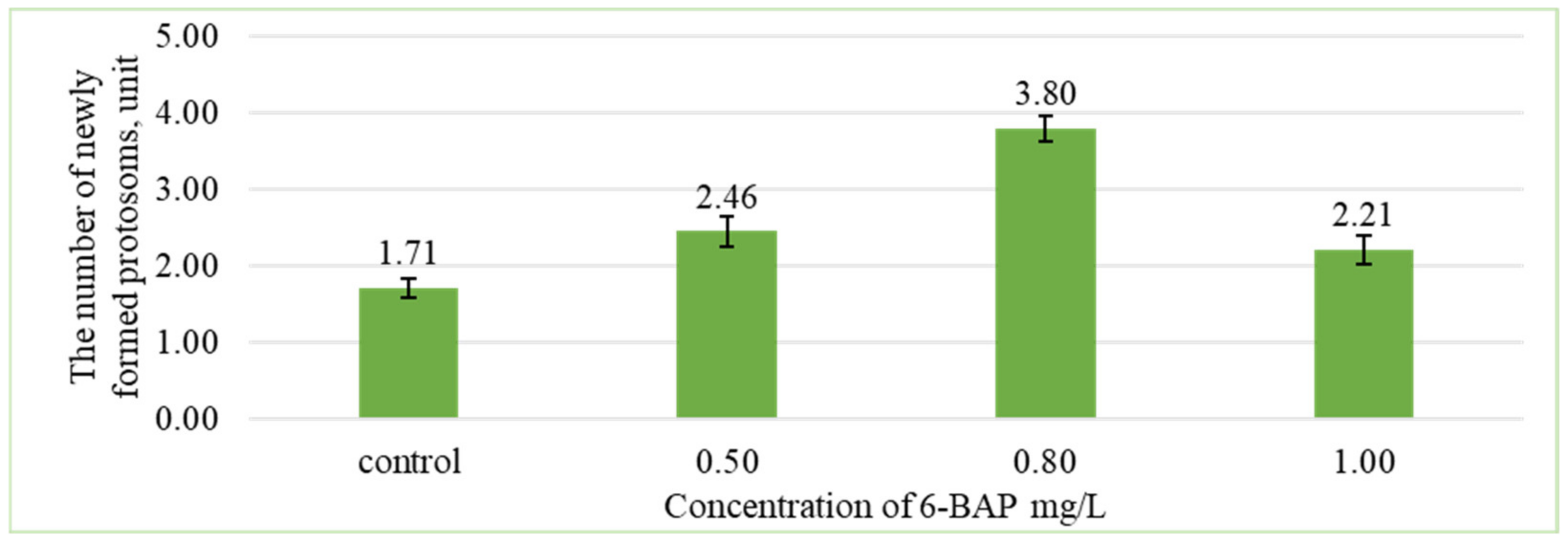
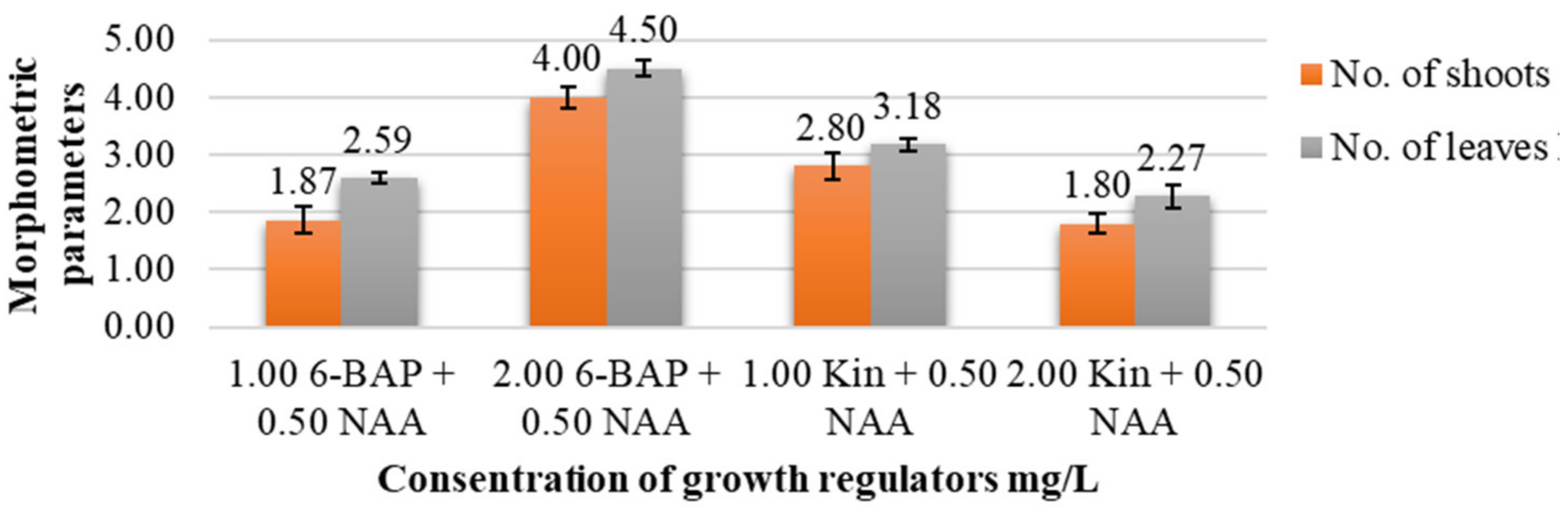
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De, L.C.; Singh, R. Organic Production of Cymbidium Orchids. Acta Sci. Agric. 2018, 2, 30–35. [Google Scholar]
- Yang, J.B.; Tang, M.; Li, H.T.; Zhang, Z.R.; Li, D.Z. Complete chloroplast genome of the genus Cymbidium: Lights into the species identification, phylogenetic implications and population genetic analyses. BMC Evol. Biol. 2013, 13, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regmi, T.; Pradhan, S.; Pant, B. In vitro Mass Propagation of an Epiphytic Orchid, Cymbidium aloifolium (L.) Sw., through Protocorm Culture. Biotechnol. J. Int. 2017, 19, 1–6. [Google Scholar] [CrossRef]
- De, L.C. Good agricultural practices of commercial orchids. Vigyan Varta 2020, 1, 53–64. [Google Scholar]
- Radhika, B.; Murthy, J.V.V.S.N.; Grace, D.N. Preliminary phytochemical analysis & antibacterial activity against clinical pathogens of medicinally important orchid Cymbidium aloifolium (L.) SW. Int. J. Pharm. Sci. Res. 2013, 4, 3925. [Google Scholar]
- White, J. Cymbidium Chatter; Cymbidium Society of Victoria: Melbourne, Australia, 2021; Volume 2, pp. 1–25. [Google Scholar]
- Kersey, M. Table Award; Atlanta Orchid Society: Atlanta, GA, USA, 2007; Volume 48, pp. 1–12. [Google Scholar]
- Averianov, L.V. Rare species of orchids (Orchidaceae) in the flora of Vietnam. Turczaninowia 2006, 9, 48–89. [Google Scholar]
- Aoyama, M. Karyomorphological studies in Cymbidium and its allied genera. Bull. Hiroshima Bot. Gard. 1989, 11, 55–57. [Google Scholar]
- De, L.C.; Rao, A.N.; Rajeevan, P.K.; Srivastava, M.; Chhetri, G. Morphological characterization in cymbidium species. J. Glob. Biosci. 2014, 3, 1060–1075. [Google Scholar]
- DeYoung, G.; Rowe, B.; Runkle, E. Propagating Orchids; American Orchid Society: Atlanta, GA, USA, 2011; pp. 486–489. [Google Scholar]
- Koh, K.W.; Lu, H.-C.; Chan, M.-T. Virus resistance in orchids. Plant Sci. 2014, 228, 26–38. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Z.; Huang, L.; Su, J. In Vitro Mass Scale Propagation of Wild Cymbidium lowianum with a Rare and Endangered Plant. Am. J. Plant Sci. 2013, 4, 34755. [Google Scholar] [CrossRef]
- Tao, J.; Yu, L.; Kong, F.; Zhao, D. Effects of plant growth regulators on in vitro propagation of Cymbidium faberi Rolfe. Afr. J. Biotechnol. 2011, 10, 15639–15646. [Google Scholar] [CrossRef]
- Mohanty, P.; Paul, S.; Das, M.C.; Kumaria, S.; Tandon, P. A simple and efficient protocol for the mass propagation of Cymbidium mastersii: An ornamental orchid of Northeast India. AoB Plants 2012, 2012, pls023. [Google Scholar] [CrossRef] [Green Version]
- Salazar-Mercado, S.A.; Vega-Contreras, N.A. Asymbiotic seed germination and in vitro propagation of Cattleya trianae Linden & Reichb.f. (Orchidaceae). Acta Agron. 2017, 66, 544–548. [Google Scholar] [CrossRef]
- Reddy, J. Nutrient media used for micropropagation of orchids: A research review. World J. Pharm. Res. 2016, 5, 1719–1732. [Google Scholar] [CrossRef]
- Cardoso, J.C.; Zanello, C.A.; Chen, J.-T. An Overview of Orchid Protocorm-Like Bodies: Mass Propagation, Biotechnology, Molecular Aspects, and Breeding. Int. J. Mol. Sci. 2020, 21, 985. [Google Scholar] [CrossRef] [Green Version]
- Kryukov, L.A.; Shirokov, A.I.; Syrova, V.V. Analysis of the potential of vegetative reproduction of tuberoid orchids at the early stages of development in vitro. J. Lobachevsky State Univ. Nizhny Novgorod 2010, 2, 413–417. [Google Scholar]
- Sidorov, A.V.; Zaytseva, Y.V.; Marakaev, O.A. Effect of supernatant of associative bacteria of the genus Pseudomonas on germination, morphogenesis, and growth of Dactylorhiza incarnata (L.) Soó (Orchidaceae) in vitro. Tomsk. State Univ. J. Biol. 2020, 51, 6–24. [Google Scholar] [CrossRef]
- Kryukov, L.A.; Shirokov, A.I.; Syrova, V.V. Multiplicity of ontogenetic patterns of Dactylorhiza incarnata (L.) soo in connection with protocorm vegetative propagation in vitro. J. Lobachevsky State Univ. Nizhny Novgorod 2011, 6, 144–148. [Google Scholar]
- Potshangbam, N.; Nirmala, C. In vitro Rapid Propagation of Cymbidium aloifolium (L.) Sw.: A Medicinally Important Orchid via Seed Culture. J. Biol. Sci. 2011, 11, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Velmyaykin, I.N.; Mokshin, E.V.; Lukatkin, A.S. Effect of growth regulators on in vitro shoot growth and propagation of Cymbidium hybridum Hort. J. Lobachevsky State Univ. Nizhny Novgorod 2013, 3, 133–137. [Google Scholar]
- Philip Robinson, J.; Jyoti, P.K.; Sebastinraj, J.; Suriya, K. In vitro seed germination of Cymbidium aloifolium (L.) Sw., a potential medicinal Orchid from Eastern Ghats of Tamil Nadu, India. J. Plant Biotechnol. 2017, 44, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Pant, B.; Swar, S. Micropropagation of Cymbidium iridioides. Nepal J. Sci. Technol. 2011, 12, 91–96. [Google Scholar] [CrossRef] [Green Version]
- Trunjaruen, A.; Taratima, W. An effective micropropagation of Cymbidium aloifolium (L.) Sw. Thai J. Bot. 2018, 10, 77–91. [Google Scholar]
- Ghosh, A.; Hossain, M.M.; Sharma, M. Mass propagation of Cymbidium giganteum Wall. ex Lindl. using in vitro seedlings. Indian J. Exp. Biol. 2014, 52, 905–911. [Google Scholar] [PubMed]
- Rittirat, S.; Klaocheed, S.; Thammasiri, K.; Prasertsongskun, S. In vitro Propagation and Forest Reestablishment of Cymbidium finlaysonianum Lindl., an Endangered Medicinal Orchid. Walailak J. Sci. Technol. (WJST) 2018, 15, 711–724. [Google Scholar] [CrossRef]
- Vyas, S.; Guha, S.; Kapoor, P.; Rao, I.U. Micropropagation of Cymbidium Sleeping Nymph through protocorm-like bodies production by thin cell layer culture. Sci. Hortic. 2009, 123, 551–557. [Google Scholar] [CrossRef]
- Peres, L.E.P.; Amar, S.; Kerbauy, G.B.; Salatino, A.; Zaffar, G.R.; Mercier, H. Effects of Auxin, Cytokinin and Ethylene Treatments on the Endogenous Ethylene and Auxin-to-cytokinins Ratio Related to direct Root Tip Conversion of Catasetum fimbriatum Lindl. (Orchidaceae) into Buds. J. Plant Physiol. 1999, 155, 551–555. [Google Scholar] [CrossRef]
- Pradhan, S.; Tiruwa, B.L.; Subedee, B.R.; Pant, B. Micropropagation of Cymbidium aloifolium (L.) SW., A Medicinal Orchid by Artificial Seeds Technology. J. Nat. Hist. Mus. 2015, 28, 42–48. [Google Scholar] [CrossRef]
- Mengarda, L.H.G.; Cola, G.P.A.; De Oliveira, S.C.; De Freitas, A.R. Multiplication, rooting in vitro, and acclimatization of Brassavola tuberculata Hook. (Orchidaceae), an orchid endemic to the Brazilian Atlantic rainforest. Biosci. J. 2017, 33, 730–738. [Google Scholar] [CrossRef] [Green Version]
- Rizky, W.H.; Nuraini, A.; Suminar, E.; Syahruddin, K. Growth and development of protocorm like bodies hybrid dendrobium orchids on MS medium with cytokinin and auxin combination. In Proceedings of the International Seminar on Horticulture to Support Food Security 2010, Bandar Lampung, Indonesia, 22–23 June 2010; pp. 210–216. [Google Scholar]
- Hartati, S.; Arniputri, R.B.; Soliah, L.A.; Cahyono, O.N.G.K.O. Effects of organic additives and naphthalene acetid acid (NAA) application on the in vitro growth of Black orchid hybrid (Coelogyne pandurata Lindley). Bulg. J. Agric. Sci. 2017, 23, 951–957. [Google Scholar]
- Rohmah, K.N.; Taratima, W. Effect of Chitosan, Coconut Water and Potato Extract on Protocorm Growth and Plantlet Regeneration of Cymbidium aloifolium (L.) Sw. Curr. Appl. Sci. Technol. 2021, 22, 1–10. [Google Scholar] [CrossRef]
- Kaur, S.; Bhutani, K.K. Organic growth supplement stimulants for in vitro multiplication of Cymbidium pendulum (Roxb.) Sw. Hortic. Sci. 2012, 39, 47–52. [Google Scholar] [CrossRef]
- Fang, Z.; Huang, W.; Zeng, S.; Wu, K. In vitro propagation of Cymbidium nanutum YS Wu et SC Chen. Propag. Ornam. Plants 2011, 11, 149–155. [Google Scholar]
- Utami, E.S.W.; Hariyanto, S. Organic Compounds: Contents and Their Role in Improving Seed Germination and Protocorm Development in Orchids. Int. J. Agron. 2020, 2020, 2795108. [Google Scholar] [CrossRef]
- Da Silva, J.A.T.; Hossain, M.M.; Sharma, M.; Dobránszki, J.; Cardoso, J.C.; Zeng, S. Acclimatization of in Vitro -derived Dendrobium. Hortic. Plant J. 2017, 3, 110–124. [Google Scholar] [CrossRef]
- Díaz, L.P.; Namur, J.J.; Bollati, S.A.; Arce, O.E.A. Acclimatization of Phalaenopsis and Cattleya obtained by micropropagation. Rev. Colomb. Biotecnol. 2010, 12, 27–40. [Google Scholar]
- Pal, R.; Dayamma, M.; Medhi, R.P. Manual for Acclimatization of Micropropagated Cymbidiums; Technical Bulletin, N° 11; CAR-NRC for Orchids: Sikkim, India, 2013; p. 20. [Google Scholar]
- Islam, S.; Islam, T.; Bhattacharjee, B.; Mondal, T.; Subramaniam, S. In vitro pseudobulb based micropropagation for mass development of Cymbidium finlaysonianum Lindl. Emir. J. Food Agric. 2015, 27, 469. [Google Scholar] [CrossRef] [Green Version]
- De, L.C.; Pathak, P.; Rao, A.N.; Rajeevan, P.K. Production Technology of Commercial Epiphytic Orchids for Cut flowers. In Commercial Orchids; De Gruyter Open: Warsaw, Poland, 2015; pp. 149–199. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration of 6-BAP mg/L | Number of Leaves, Unit | Number of Roots, Unit | Root Length, cm |
|---|---|---|---|
| Control | 2.22 ± 0.10 * | 1.14 ± 0.10 | 0.48 ± 0.11 |
| 0.5 | 2.89 ± 0.21 | 1.14 ± 0.10 | 0.76 ± 0.13 |
| 0.8 | 2.61 ± 0.23 | 1.07 ± 0.07 | 0.60 ± 0.13 |
| 1.0 | 2.83 ± 0.22 | 1.14 ± 0.18 | 0.73 ± 0.14 |
| Concentration of Growth Regulators, mg/L | Plant Height, cm | Number of Shoots, Unit | Number of Leaves, Unit | Number of Roots, Unit | Root Length, cm |
|---|---|---|---|---|---|
| 1.0 6-BAP + 0.5 NAA | 2.22 ± 0.20 a * | 1.86 ± 0.23 a | 2.59 ± 0.10 a | 1.31 ± 0.15 a | 1.00 ± 0.08 ab |
| 2.0 6-BAP + 0.5 NAA | 3.19 ± 0.17 b | 4.00 ± 0.19 c | 4.50 ± 0.14 c | 1.93 ± 0.11 b | 1.59 ± 0.12 b |
| 1.0 Kin + 0.5 NAA | 3.00 ± 0.20 b | 2.80 ± 0.22 b | 3.18 ± 0.11 b | 1.87 ± 0.17 b | 1.31 ± 0.14 b |
| 2.0 Kin + 0.5 NAA | 2.09 ± 0.16 a | 1.80 ± 0.17 a | 2.27 ± 0.19 a | 1.37 ± 0.12 a | 0.83 ± 0.04 a |
| LSD05 | 0.59 | 0.99 | 0.45 | 0.43 | 0.37 |
| Organic Additives | Number of Roots, Unit | Root Length, cm |
|---|---|---|
| Control | 1.69 ± 0.11 a * | 1.21 ± 0.10 a |
| coconut water | 2.24 ± 0.14 b | 3.53 ± 0.15 c |
| banana puree | 3.93 ± 0.20 c | 1.72 ± 0.48 b |
| sweet potatoes | 2.86 ± 0.12 b | 2.14 ± 0.10 b |
| LSD05 | 0.54 | 0.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussien, M.; Kryuchkova, V.; Raeva-Bogoslovskaya, E.; Molkanova, O. Clonal Micropropagation of Cymbidium erythrostylum Rolfe. Int. J. Plant Biol. 2023, 14, 28-38. https://doi.org/10.3390/ijpb14010003
Hussien M, Kryuchkova V, Raeva-Bogoslovskaya E, Molkanova O. Clonal Micropropagation of Cymbidium erythrostylum Rolfe. International Journal of Plant Biology. 2023; 14(1):28-38. https://doi.org/10.3390/ijpb14010003
Chicago/Turabian StyleHussien, Muthab, Viktoriya Kryuchkova, Ekaterina Raeva-Bogoslovskaya, and Olga Molkanova. 2023. "Clonal Micropropagation of Cymbidium erythrostylum Rolfe" International Journal of Plant Biology 14, no. 1: 28-38. https://doi.org/10.3390/ijpb14010003
APA StyleHussien, M., Kryuchkova, V., Raeva-Bogoslovskaya, E., & Molkanova, O. (2023). Clonal Micropropagation of Cymbidium erythrostylum Rolfe. International Journal of Plant Biology, 14(1), 28-38. https://doi.org/10.3390/ijpb14010003