Re-Evaluation of the Impacts of Dietary Preferences on Macroinvertebrate Trophic Sources: An Analysis of Seaweed Bed Habitats Using the Integration of Stable Isotope and Observational Data

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sampling
2.2.1. Potential Food Sources
2.2.2. Consumers
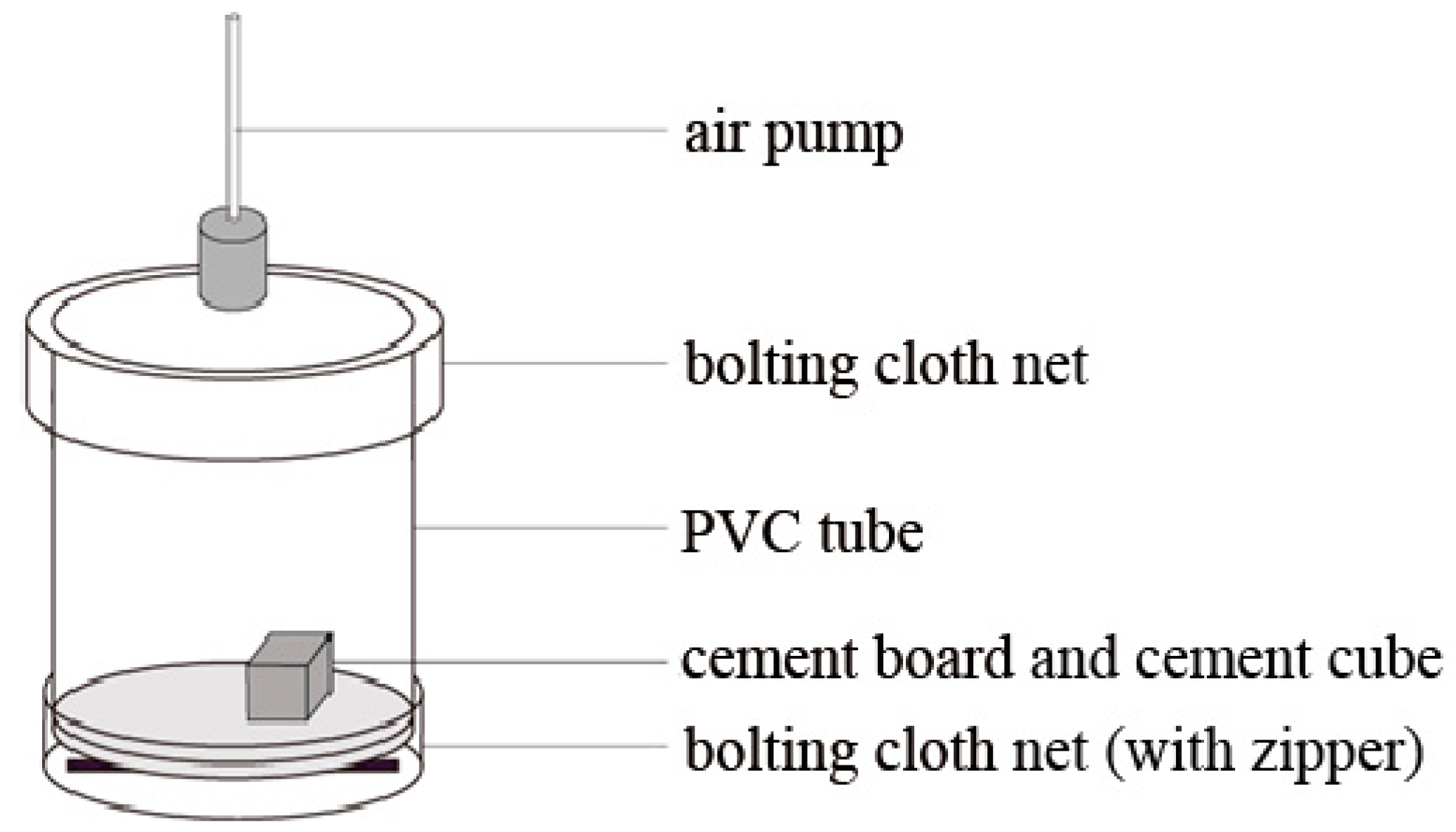
2.3. Species-Specific Diet Selection Observation
2.4. Estimation of Discrimination between Diets and Macroinvertebrates
2.5. Stable Isotope Analysis
2.6. Data Analysis
3. Results
3.1. Dietary Observations: Mobility, Mode and Preference
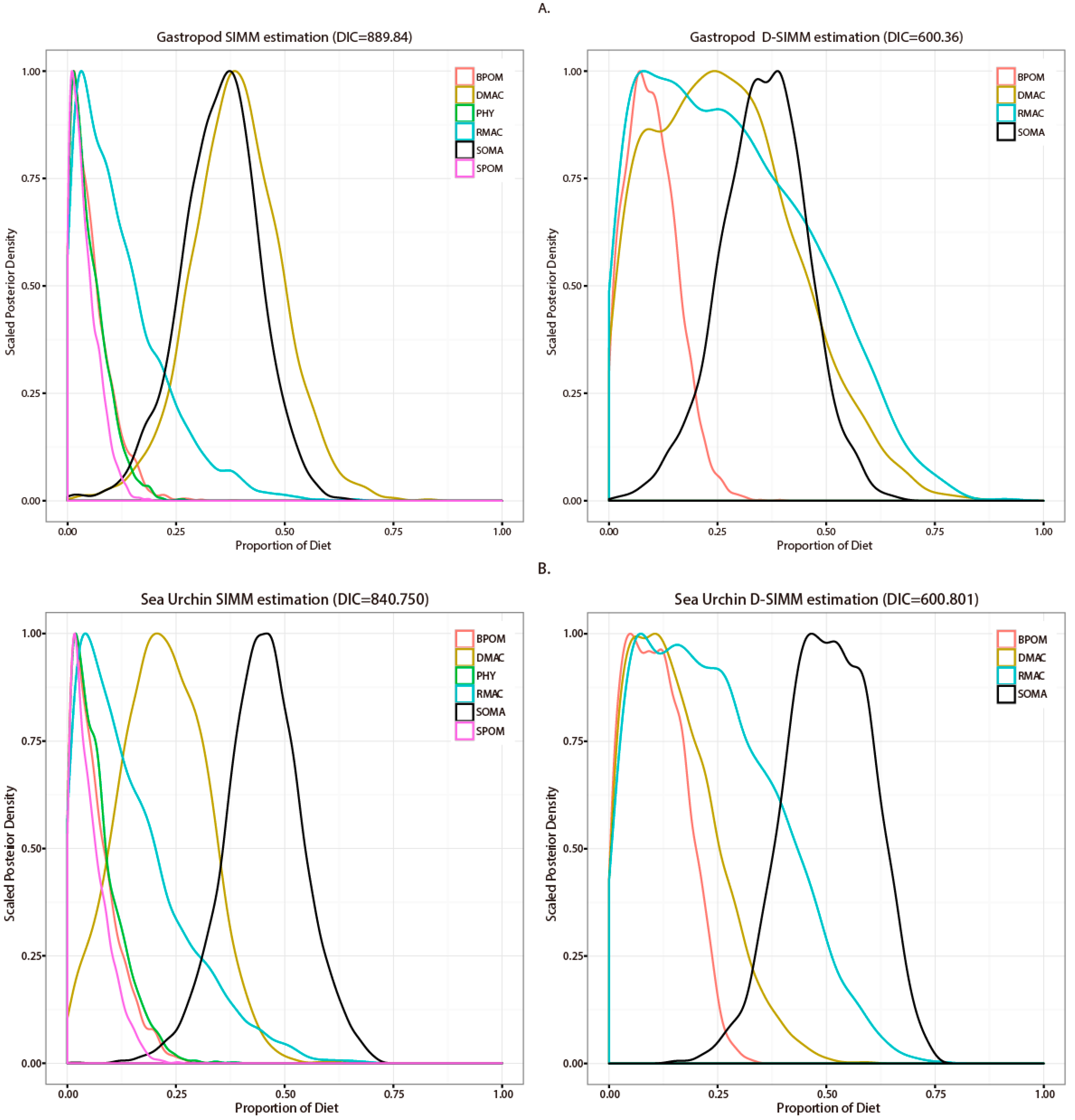
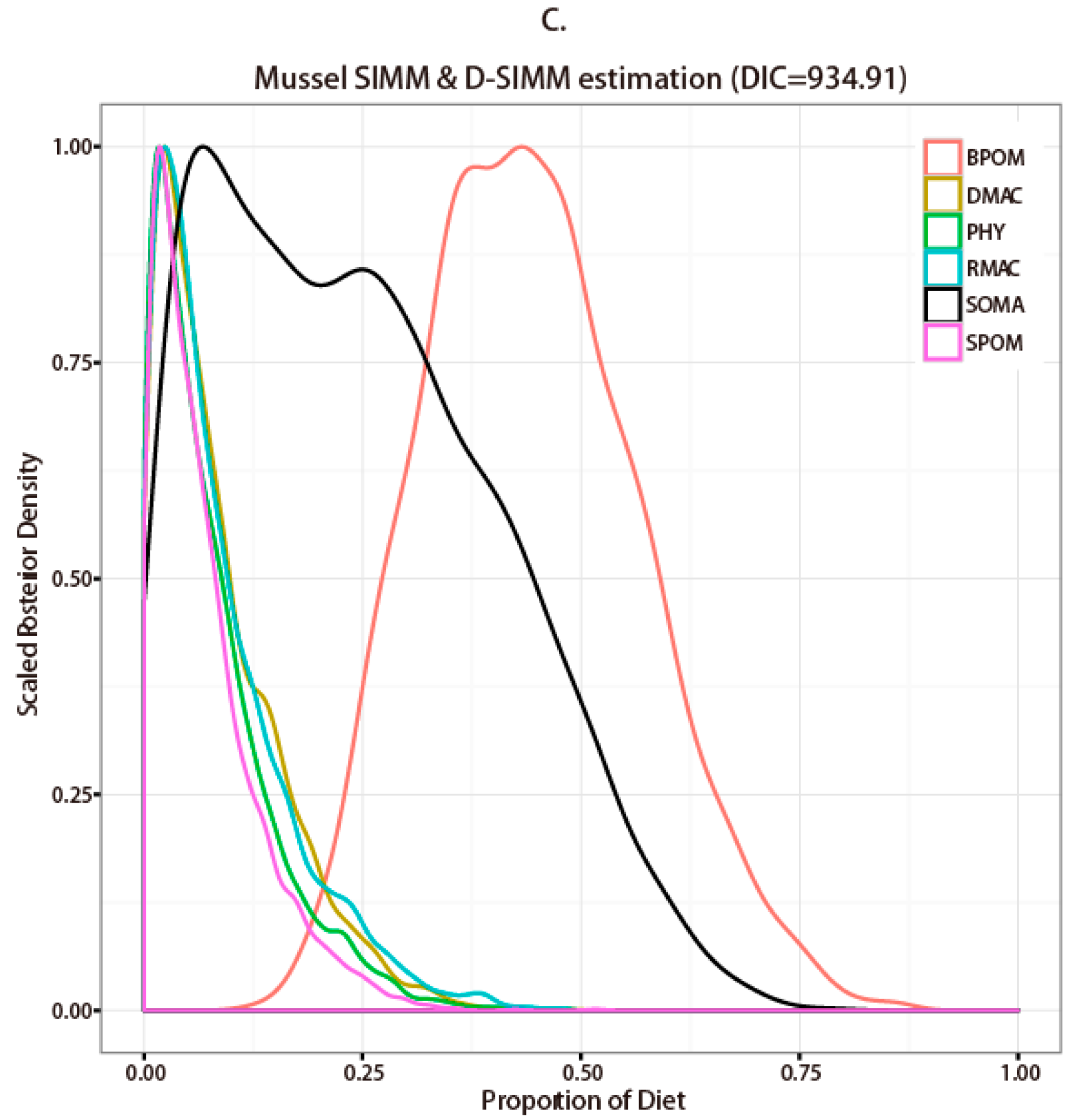
3.2. Stable Isotope Analysis and Source Contribution Evaluation
4. Discussion
4.1. Comparison between SIMM and D-SIMM
4.2. Diet Estimation of Representative Macroinvertebrate Species in Seaweed Beds
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Paine, R.T. Road Maps of Interactions or Grist for Theoretical Development? Ecology 1988, 69, 1648–1654. [Google Scholar] [CrossRef]
- Bertness, M.D.; Leonard, G.H. The role of positive interactions in communities: Lessons from intertidal habitats. Ecology 1997, 78, 1976–1989. [Google Scholar] [CrossRef]
- Bertness, M.D.; Leonard, G.H.; Levine, J.M.; Schmidt, P.R.; Ingraham, A.O. Testing the relative contribution of positive and negative interactions in rocky intertidal communities. Ecology 1999, 80, 2711–2726. [Google Scholar] [CrossRef]
- Whittaker, R.H.; Niering, W.A. Vegetation of the Santa Catalina Mountains, Arizona. V. Biomass, Production, and Diversity along the Elevation Gradient. Ecology 1975, 56, 771–790. [Google Scholar] [CrossRef]
- Reviews, B.; Iv, V.; Assemblages, O.; Ecology, M.; Geography, E.A.; Mann, K.H. Ecology of Coastal Waters—A Systems Approach. Studies in Ecology; Blackwell Scientific Publications: Oxford, UK, 1982; Volume 8, pp. 340–341. [Google Scholar]
- Zabala, S.; Bigatti, G.; Botto, F.; Iribarne, O.O.; Galván, D.E. Trophic relationships between a Patagonian gastropod and its epibiotic anemone revealed by using stable isotopes and direct observations. Mar. Biol. 2013, 160, 909–919. [Google Scholar] [CrossRef]
- Harrold, C.; Lisin, S. Radio-tracking fafts of giant-kelp—Local production and regional transport. J. Exp. Mar. Biol. Ecol. 1989, 130, 237–251. [Google Scholar] [CrossRef]
- Pollock, F.J.; Lamb, J.B.; Field, S.N.; Heron, S.F.; Schaffelke, B.; Shedrawi, G.; Bourne, D.G.; Willis, B.L. Sediment and turbidity associated with offshore dredging increase coral disease prevalence on nearby reefs. PLoS ONE 2014, 9, e102498. [Google Scholar] [CrossRef] [PubMed]
- Colombo, F.; Costa, V.; Dubois, S.F.; Gianguzza, P.; Mazzola, A.; Vizzini, S. Trophic structure of vermetid reef community: High trophic diversity at small spatial scales. J. Sea Res. 2013, 77, 93–99. [Google Scholar] [CrossRef]
- Lunt, J.; Smee, D.L. Turbidity influences trophic interactions in estuaries. Limnol. Oceanogr. 2014, 59, 2002–2012. [Google Scholar] [CrossRef] [Green Version]
- Duggins, D.O.; Eckman, J.E. The role of kelp detritus in the growth of benthic suspension feeders in an understory kelp forest. J. Exp. Mar. Biol. Ecol. 1994, 176, 53–68. [Google Scholar] [CrossRef]
- Abrantes, K.G.; Sheaves, M. Importance of freshwater flow in terrestrial-aquatic energetic connectivity in intermittently connected estuaries of tropical Australia. Mar. Biol. 2010, 157, 2071–2086. [Google Scholar] [CrossRef]
- Cardoso, P.G.; Raffaelli, D.; Lillebø, A.I.; Verdelhos, T.; Pardal, M.A. The impact of extreme flooding events and anthropogenic stressors on the macrobenthic communities’ dynamics. Estuar. Coast. Shelf Sci. 2008, 76, 553–565. [Google Scholar] [CrossRef] [Green Version]
- Duffy, J.E.; Cardinale, B.J.; France, K.E.; McIntyre, P.B.; Thébault, E.; Loreau, M. The functional role of biodiversity in ecosystems: Incorporating trophic complexity. Ecol. Lett. 2007, 10, 522–538. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.F.; Shin, P.K.S.; Lin, G.H.; Chen, S.P.; Siu, G.C. Stable isotope and fatty acid evidence for uptake of organic waste by green-lipped mussels Perna viridis in a polyculture fish farm system. Mar. Ecol. Prog. Ser. 2006, 317, 273–283. [Google Scholar] [CrossRef]
- Pauly, D.; Watson, R. Background and interpretation of the “Marine Trophic Index” as a measure of biodiversity. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Leitão, R.; Martinho, F.; Neto, J.M.; Cabral, H.; Marques, J.C.; Pardal, M.A. Feeding ecology, population structure and distribution of Pomatoschistus microps (Krøyer, 1838) and Pomatoschistus minutus (Pallas, 1770) in a temperate estuary, Portugal. Estuar. Coast. Shelf Sci. 2006, 66, 231–239. [Google Scholar] [CrossRef]
- Hopkins, J.B.; Ferguson, J.M. Estimating the diets of animals using stable isotopes and a comprehensive Bayesian mixing model. PLoS ONE 2012, 7, e28478. [Google Scholar] [CrossRef]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER—Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Ogle, K.; Tucker, C.; Cable, J.M. Beyond simple linear mixing models: Process-based isotope partitioning of ecological processes. Ecol. Appl. 2014, 24, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Winemiller, K.O.; Akin, S.; Zeug, S.C. Production sources and food web structure of a temperate tidal estuary: Integration of dietary and stable isotope data. Mar. Ecol. Prog. Ser. 2007, 343, 63–76. [Google Scholar] [CrossRef]
- Savini, D.; Occhipinti-Ambrogi, A. Consumption rates and prey preference of the invasive gastropod Rapana venosa in the Northern Adriatic Sea. Helgol. Mar. Res. 2006, 60, 153–159. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, S.; Wang, X.; Rijin, J.; Zhao, J. The feeding behaviour and ecological function during summer of one herbivore on seaweed bed in Gouqi Island: The gastropod, Turbo cornutus Solander. J. Fish. China 2015, 39, 511–519. [Google Scholar]
- Pearson, T.H.; Rosenberg, R. Feast and Famine: Structuring Factors in Marine Benthic Communities; Symposium of the British Ecological Society: Sussex, UK, 1987. [Google Scholar]
- Fauchald, K.; Jumars, P.A. The diet of worms: A study of polychaete feeding guilds. Oceanogr. Mar. Biol. Annu. Rev. 1979, 17, 193–284. [Google Scholar]
- McCutchan, J.H.; Lewis, W.M.; Kendall, C.; McGrath, C.C. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos 2003, 102, 378–390. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Rasmussen, J.B. Primary consumer delta C-13 and delta N-15 and the trophic position of aquatic consumers. Ecology 1999, 80, 1395–1404. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: http://www.R-project.org/ (accessed on 16 March 2016).
- Semmens, B.X.; Stock, B.C.; Ward, E.; Moore, J.W.; Parnell, A.; Jackson, A.L.; Phillips, D.L.; Bearhop, S.; Inger, R. MixSIAR: A Bayesian Stable Isotope Mixing Model for Characterizing Intrapopulation Niche Variation; ESA Convention: Minneapolis, MN, USA, 2013. [Google Scholar]
- Jackson, M.C.; Donohue, I.; Jackson, A.L.; Britton, J.R.; Harper, D.M.; Grey, J. Population-level metrics of trophic structure based on stable isotopes and their application to invasion ecology. PLoS ONE 2012, 7, e31757. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.W.; Semmens, B.X. Incorporating uncertainty and prior information into stable isotope mixing models. Ecol. Lett. 2010, 11, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A. On the Interpretation of χ2 from Contingency Tables, and the Calculation of P. J. R. Stat. Soc. 1922, 85, 87–94. [Google Scholar] [CrossRef]
- Gelman, A.A.; Carlin, J.B.; Stern, H.S.; Donald, B. Bayesian Data Analysis; Chapman & Hall/CRC, cop.: Boca Raton, FL, USA, 1995. [Google Scholar]
- Saito, L.; Redd, C.; Chandra, S.; Atwell, L.; Fritsen, C.H.; Rosen, M.R. Quantifying foodweb interactions with simultaneous linear equations: Stable isotope models of the Truckee River, USA. J. N. Am. Benthol. Soc. 2007, 26, 642–662. [Google Scholar] [CrossRef]
- Yokoyama, H.; Ishihi, Y. Variation in δ 13C and δ 15N among different tissues of three estuarine bivalves: Implications for dietary. Plankton Benthos Res. 2006, 1, 178–182. [Google Scholar] [CrossRef]
- Kasai, A.; Ishizaki, D.; Isoda, T. Isotopic trophic-step fractionation of the freshwater clam Corbicula sandai. Fish. Sci. 2016, 82, 491–498. [Google Scholar] [CrossRef]
- Dubois, S.; Jean-Louis, B.; Bertrand, B.; Lefebvre, S. Isotope trophic-step fractionation of suspension-feeding species: Implications for food partitioning in coastal ecosystems. J. Exp. Mar. Biol. Ecol. 2007, 351, 121–128. [Google Scholar] [CrossRef]
- Fry, B. Food Web Structure on Georges Bank from Stable C, N, and S Isotopic Compositions. Limnol. Oceanogr. 1988, 33, 1182–1190. [Google Scholar] [CrossRef]
- Gates, E.N. Determining Carbon and Nitrogen Stable Isotope Discrimination for Marine Consumers. Ph.D. Thesis, Edith Cowan University, Joondalup, WA, Australia, 2006. [Google Scholar]
- Barton, D.R.; Johnson, R.A.; Campbell, L.; Petruniak, J.; Patterson, M. Effects of round gobies (Neogobius melanostomus) on dreissenid mussels and other invertebrates in Eastern Lake Erie, 2002–2004. J. Great Lakes Res. 2005, 31, 252–261. [Google Scholar] [CrossRef]
- Layman, C.A.; Quattrochi, J.P.; Peyer, C.M.; Allgeier, J.E. Niche width collapse in a resilient top predator following ecosystem fragmentation. Ecol. Lett. 2007, 10, 937–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, V.; Walters, C.J. Ecopath with Ecosim: Methods, capabilities and limitations. Ecol. Model. 2004, 172, 109–139. [Google Scholar] [CrossRef]
- Fulton, E.A.; Smith, A.D.M.; Johnson, C.R. Biogeochemical marine ecosystem models I: IGBEM—A model of marine bay ecosystems. Ecol. Model. 2004, 174, 267–307. [Google Scholar] [CrossRef]
- Plagányi, É.E.; Punt, A.E.; Hillary, R.; Morello, E.B.; Thébaud, O.; Hutton, T.; Pillans, R.D.; Thorson, J.T.; Fulton, E.A.; Smith, A.D.M.; et al. Multispecies fisheries management and conservation: Tactical applications using models of intermediate complexity. Fish Fish. 2014, 15, 1–22. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Multimodel inference: Understanding AIC and BIC in model selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Bhat, H.; Kumar, N. On the Derivation of the Bayesian Information Criterion; School of Natural Sciences, University of California: Oakland, CA, USA, 2015. [Google Scholar]
- Spiegelhalter, D.J.; Best, N.G.; Carlin, B.P.; Van der Linde, A. The deviance information criterion: 12 years on. J. R. Stat. Soc. Ser. B Stat. Methodol. 2014, 76, 485–493. [Google Scholar] [CrossRef]
- Gurney, L.J.; Froneman, P.W.; Pakhomov, E.A.; McQuaid, C.D. Trophic positions of three euphausiid species from the Prince Edward Islands (Southern Ocean): Implications for the pelagic food web structure. Mar. Ecol. Prog. Ser. 2001, 217, 167–174. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, S.; Chen, Y.; Bi, Y. Analysis of functional feeding groups of macroinvertebrates communities in the macroalgae beds of Gouqi Island, Zhejiang Province. J. Fish. China 2015, 39, 381–391. [Google Scholar]
- Fields, L.; Nixon, S.W.; Oviatt, C.; Fulweiler, R.W. Benthic metabolism and nutrient regeneration in hydrographically different regions on the inner continental shelf of Southern New England. Estuar. Coast. Shelf Sci. 2014, 148, 14–26. [Google Scholar] [CrossRef]
- Jiang, R.; Zhang, S.; Bi, Y.; Wang, Z.; Zhou, X.; Zhao, X.; Chen, L. Food sources of small invertebrates in the macroalgal bed of Gouqi Island. J. Fish. China 2015, 39, 1487–1498. [Google Scholar]
- Jiang, R.; Zhang, S.; Wang, K.; Zhou, X.; Zhao, J. Stable isotope analysis of the offshore food web of Gouqi Island. Chin. J. Ecol. 2014, 33, 930–938. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Sample Size (n) | δ13C (‰) | δ15N (‰) | ||
|---|---|---|---|---|---|
| Mean | SD | Mean | SD | ||
| Potential trophic source | |||||
| PHY | 12 | −20.64 | 0.92 | 5.84 | 2.02 |
| SPOM | 10 | −20.92 | 0.86 | 6.69 | 0.62 |
| BPOM | 11 | −15.42 | 1.63 | 8.76 | 1.88 |
| SOMA | 12 | −18.38 | 0.73 | 2.75 | 0.24 |
| SOMB | 18 | −21.95 | 0.51 | 2.53 | 0.51 |
| MADS | 39 | −14.55 | 3.08 | 4.99 | 1.55 |
| Corallina officinalis | 3 | −8.48 | 0.73 | 6.6 | 0.55 |
| Jania decussato-dichotoma | 3 | −8.93 | 0.74 | 6.82 | 0.36 |
| Ulva pertuca | 11 | −14.3 | 0.41 | 5.76 | 1.26 |
| Dictyopteris dichotoma | 3 | −14.77 | 3.19 | 5.06 | 1.26 |
| Sargassum vachellianum | 5 | −15.72 | 2.81 | 4.53 | 1.03 |
| Sargassum horueri | 10 | −16.83 | 1.54 | 3.8 | 1.59 |
| Sargassum fusiforme | 4 | −16.7 | 0.7 | 3.84 | 0.54 |
| MARS | 22 | −17.04 | 1.6 | 5.29 | 1.2 |
| Garteloupia livida | 3 | −14.45 | 0.51 | 4.55 | 0.09 |
| Chondria crassicaulis | 3 | −16.83 | 1.28 | 5.31 | 2.94 |
| Undaria pinnatifida | 7 | −16.39 | 0.65 | 5.17 | 0.84 |
| Codium fragile | 3 | −18.54 | 0.29 | 5.26 | 1.08 |
| Ceramium japonicum | 3 | −19.16 | 0.25 | 6.23 | 0.22 |
| Hypnea cervicornis | 3 | −18.55 | 0.34 | 5.78 | 0.61 |
| Consumers | |||||
| Gastropod (T. cornutus) | 26 | −16.3 | 0.85 | 5.86 | 0.54 |
| Sea urchin (A. crassispina) | 17 | −15.44 | 0.65 | 5.08 | 0.5 |
| Mussel (S. virgatus) | 41 | −15.97 | 0.64 | 6.15 | 0.62 |
| Description | Species | Functional Group | Potential Trophic Source | |||
|---|---|---|---|---|---|---|
| MAC | PHY | SOM | POM | |||
| Sea urchin | A. crassispina | Semi-mobile jawed surface omnivore (herbivore and detritus feeder) | ●a | ×c | ○b | ○b |
| Gastropod | T. cornutus | Semi-mobile jawed surface omnivore (herbivore and detritus feeder) | ●a | ×c | ●a | ○b |
| Mussel | S. virgatus | Sessile filter-feeder (MAC and PHY and POM and SOM) | ●a | ●a | ●a | ●a |
| Potential Trophic Source | Replicate Number of Experiments (n) | Macroinvertebrate—MAC Dietary Preference | |
|---|---|---|---|
| Sea Urchin | Gastropod | ||
| Chlorophyta | |||
| Ulva pertuca | 6 | ●a | ●a |
| Codium fragile | 6 | ○b | ×c |
| Rhodophyta | |||
| Ceramium japonicum | 6 | ○b | ○b |
| Chondria crassicaulis | 6 | ○b | ×c |
| Corallina officinalis | 6 | ×c | ×c |
| Jania decussato dichotoma | 6 | ×c | ×c |
| Hypnea cervicornis J.Ag. | 6 | ○b | ×c |
| Garteloupia kurogii | 6 | ●a | ●a |
| Phaeophyta | |||
| Sargassum fusiforme | 6 | ●a | ●a |
| Sargassum horueri | 6 | ●a | ●a |
| Dictyopteris dichotoma | 6 | ●a | ○b |
| Ishige okamurai | 6 | ●a | ○b |
| Hizikia fusifarme | 6 | ●a | ●a |
| Group | Tissue | Diet | Lab. or Field | Δ13C (‰) | Δ15N (‰) | SDΔ13C (‰) | SDΔ15N (‰) | Ref. |
|---|---|---|---|---|---|---|---|---|
| Bivalves (M. veneriformis) | Muscle | POM | Lab. | 0.9 | 3.6 | [35] | ||
| Bivalves (R. philippinarum) | Muscle | POM | Lab. | 0.6 | 3.4 | [35] | ||
| Bivalves (C. sandai) | Soft tissue | PHY | Lab. | 0.7 | 2.1 | [36] | ||
| Bivalves (M. edulis) | Muscle | PHY | Lab. | 2.17 | 3.78 | 0.324 | 0.292 | [37] |
| Bivalves (C. gigas) | Muscle | PHY | Lab. | 1.85 | 3.79 | 0.194 | 0.194 | [37] |
| Bivalves (C. sandai) | Soft tissue | MAC | Lab. | 0.6 | 3.6 | [36] | ||
| Bivalves (C. sandai) | Soft tissue | MAC | Lab. | 0.1 | 3.3 | [36] | ||
| Bivalves (Scallops) | Soft tissue | POM | Field | 3.8 | 0.9 | [38] | ||
| Bivalves (Mussel) | Soft tissue | POM | Field | 3.4 | 1.8 | [38] | ||
| Sea urchin (H. erythrogramma) | Muscle | MAC (Phaeophytes) | Lab. | 1.85 | 2.44 | [39] | ||
| Sea urchin (H. erythrogramma) | Muscle | MAC (Fleshy Rhodophytes) | Lab. | 3.23 | 3.96 | [39] | ||
| Sea urchin (H. erythrogramma) | Muscle | MAC (Calcareous Rhodophytes) | Lab. | 4.39 | 3.02 | [39] | ||
| Sea urchin (H. erythrogramma) | Muscle | MAC (Chlorophytes) | Lab. | 1.2 | 3.15 | [39] | ||
| Gastropod (T. torquatus) | Foot tissue | MAC (Fleshy Rhodophytes) | Lab. | −0.17 | 1.56 | [39] | ||
| Gastropod (T. torquatus) | Foot tissue | MAC (Calcareous Rhodophytes) | Lab. | 0.14 | 1.06 | [39] | ||
| Gastropod (T. cornutus) | Foot tissue | MAC and SOM | Lab. | 0.43 | 1.43 | 0.13 | 0.8 | Present study |
| Sea urchin (A. crassispina) | Gonad | MAC and SOM | Lab. | 1.93 | 0.8 | 1.1 | 0.2 | Present study |
| Source | TA | SEA | SEAC |
|---|---|---|---|
| SIMM (gastropod and sea urchin) | |||
| DMAC | 51.41 | 13.66 | 14.03 |
| RMAC | 16.79 | 6.09 | 6.39 |
| SOMA | 1.21 | 0.54 | 0.6 |
| BPOM | 20.35 | 9.48 | 10.54 |
| SPOM | 2.21 | 1.37 | 1.54 |
| PHY | 10.36 | 5.12 | 5.64 |
| Gastropod D-SIMM | |||
| DMAC | 35.82 | 9.13 | 9.43 |
| RMAC | 10.9 | 4.98 | 5.44 |
| SOMA | 1.12 | 0.54 | 0.6 |
| BPOM | 20.35 | 9.48 | 10.54 |
| Sea Urchin D-SIMM | |||
| DMAC | 35.82 | 9.13 | 9.43 |
| RMAC | 16.79 | 6.34 | 6.71 |
| SOMA | 1.12 | 0.54 | 0.6 |
| BPOM | 20.35 | 9.48 | 10.54 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Liu, Y.; Wang, K.; Zhao, J.; Zhao, X.; Zhang, S. Re-Evaluation of the Impacts of Dietary Preferences on Macroinvertebrate Trophic Sources: An Analysis of Seaweed Bed Habitats Using the Integration of Stable Isotope and Observational Data. Sustainability 2018, 10, 2010. https://doi.org/10.3390/su10062010
Zhou X, Liu Y, Wang K, Zhao J, Zhao X, Zhang S. Re-Evaluation of the Impacts of Dietary Preferences on Macroinvertebrate Trophic Sources: An Analysis of Seaweed Bed Habitats Using the Integration of Stable Isotope and Observational Data. Sustainability. 2018; 10(6):2010. https://doi.org/10.3390/su10062010
Chicago/Turabian StyleZhou, Xijie, Yumeng Liu, Kai Wang, Jing Zhao, Xu Zhao, and Shouyu Zhang. 2018. "Re-Evaluation of the Impacts of Dietary Preferences on Macroinvertebrate Trophic Sources: An Analysis of Seaweed Bed Habitats Using the Integration of Stable Isotope and Observational Data" Sustainability 10, no. 6: 2010. https://doi.org/10.3390/su10062010
APA StyleZhou, X., Liu, Y., Wang, K., Zhao, J., Zhao, X., & Zhang, S. (2018). Re-Evaluation of the Impacts of Dietary Preferences on Macroinvertebrate Trophic Sources: An Analysis of Seaweed Bed Habitats Using the Integration of Stable Isotope and Observational Data. Sustainability, 10(6), 2010. https://doi.org/10.3390/su10062010