Amelioration of Composts for Greenhouse Vegetable Plants Using Pasteurised Agaricus Mushroom Substrate



Abstract
:1. Introduction
2. Materials and Methods
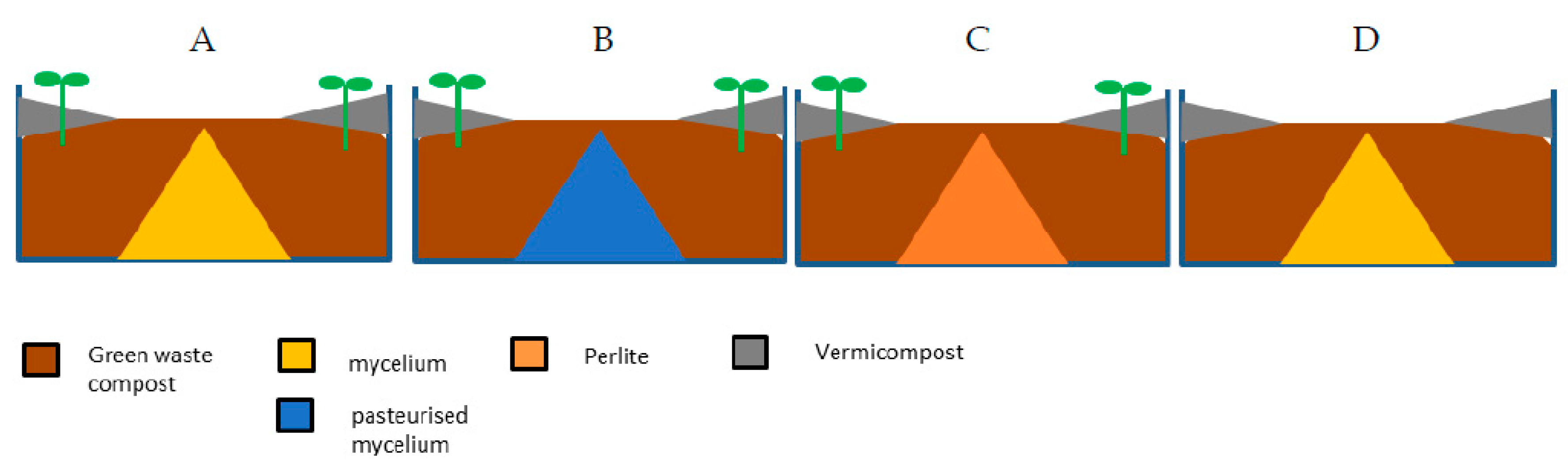
2.1. Substrates
2.2. Lettuce Cultivation
2.3. Lettuce Experiment 1
2.4. Lettuce Experiment 2
2.5. Lettuce Experiment 3
2.6. Lettuce Experiment 4
2.7. Lettuce Experiment 5
2.8. Tomato Experiment
2.9. Combined Experiment
2.10. Sampling and Analytical Methods
2.11. Statistical Analysis
3. Results
3.1. Chemical Composition of Materials
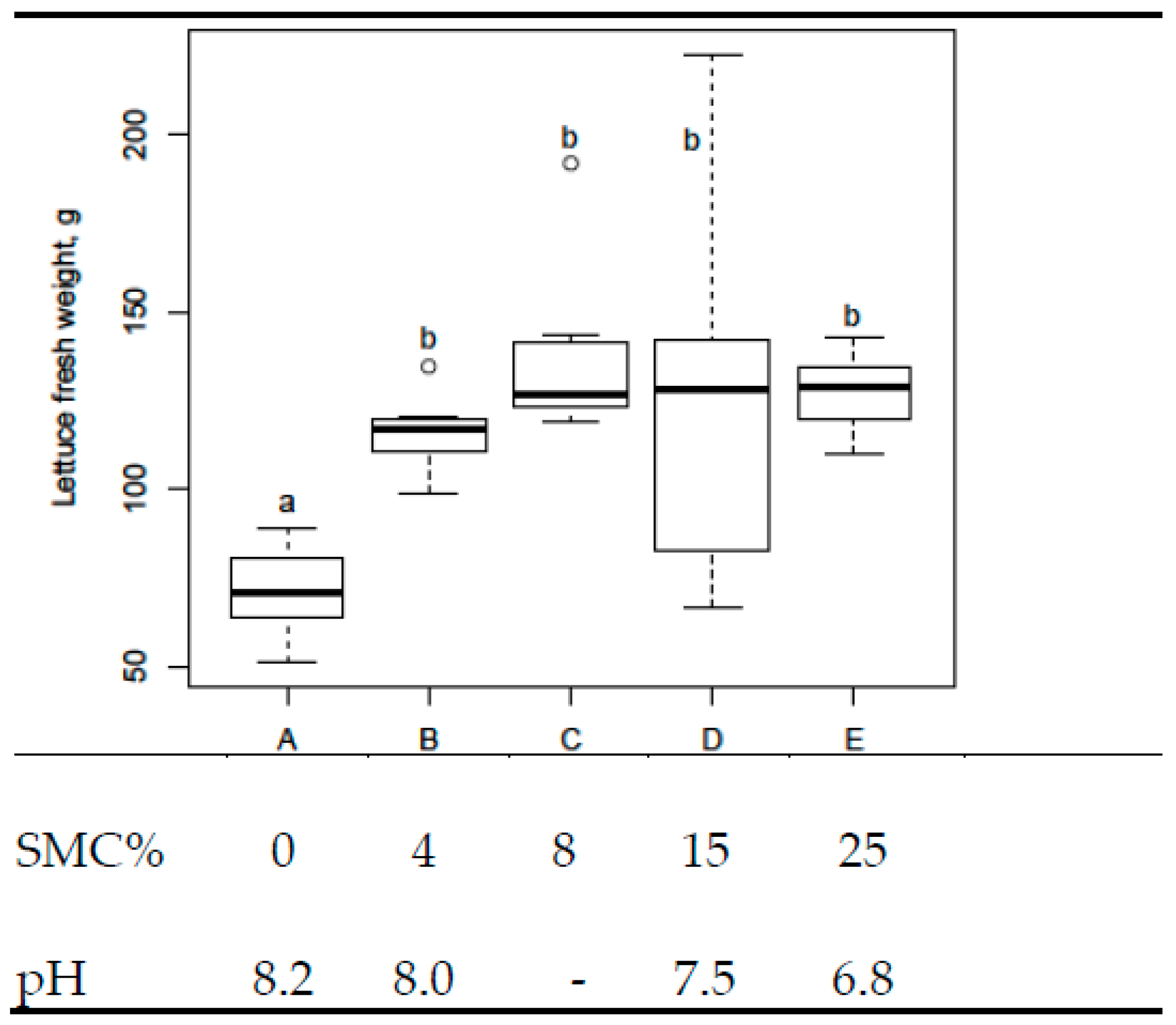
3.2. Lettuce Experiment 1
3.3. Lettuce Experiment 2
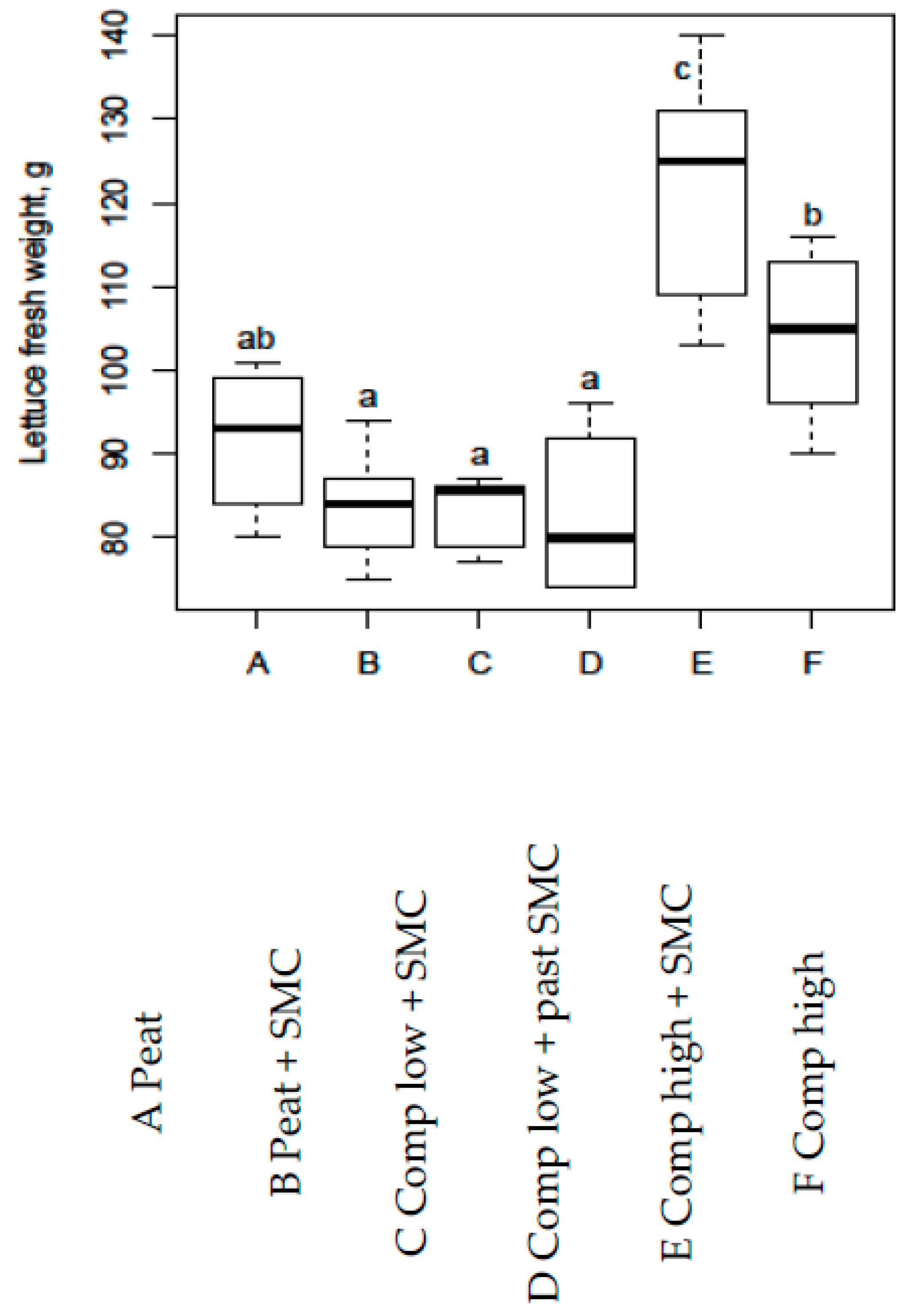
3.4. Lettuce Experiment 3
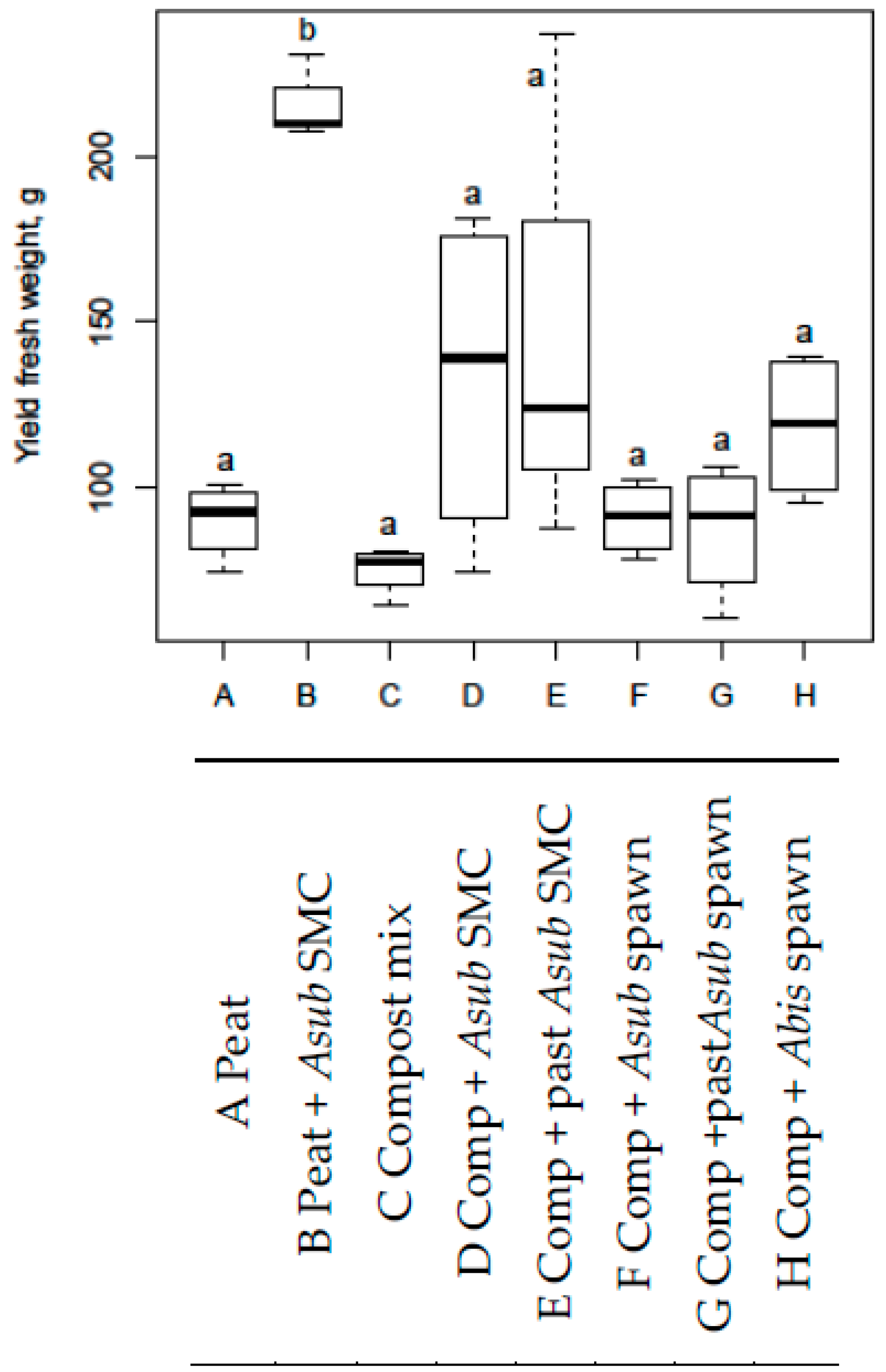
3.5. Lettuce Experiment 4
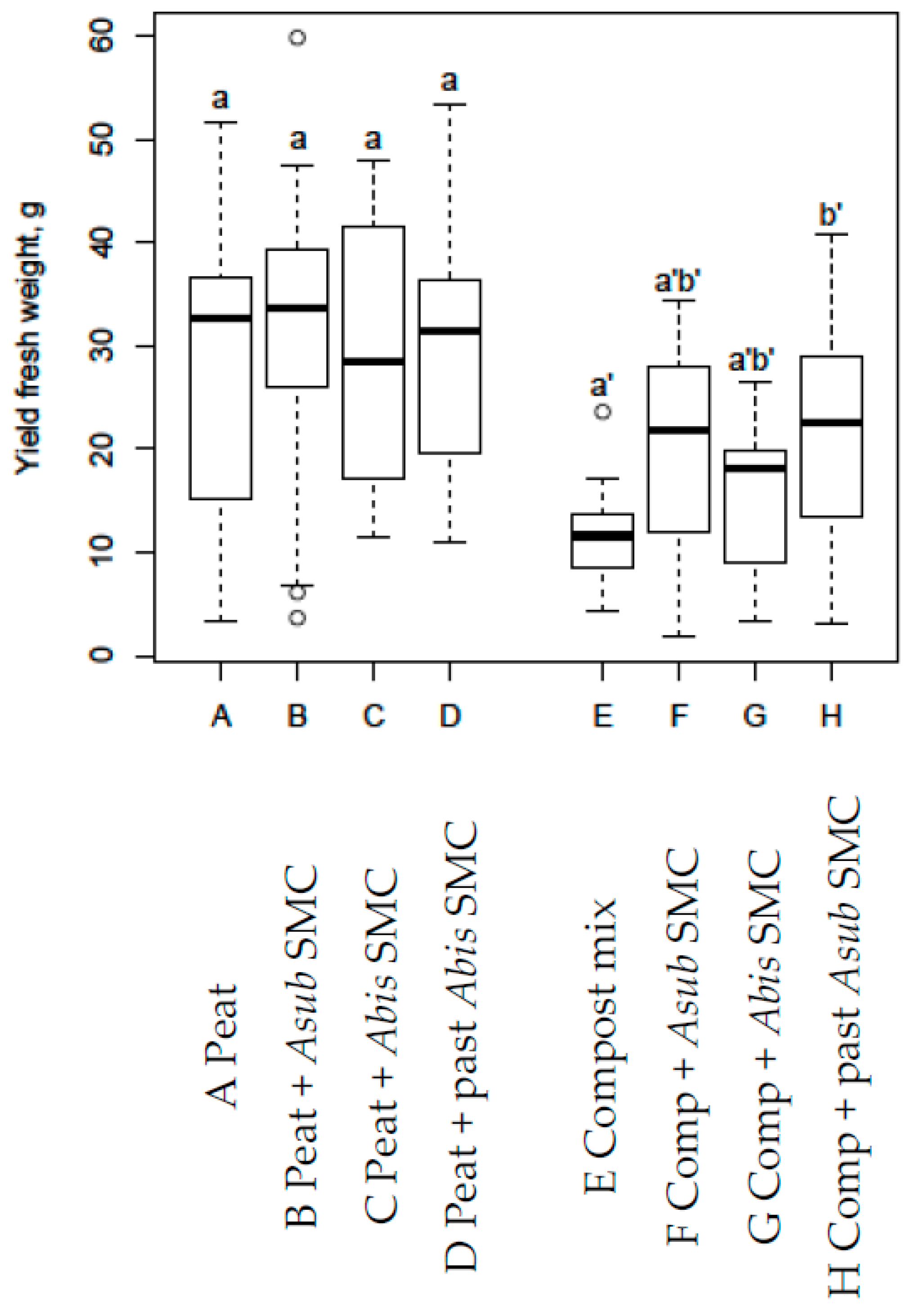
3.6. Lettuce Experiment 5
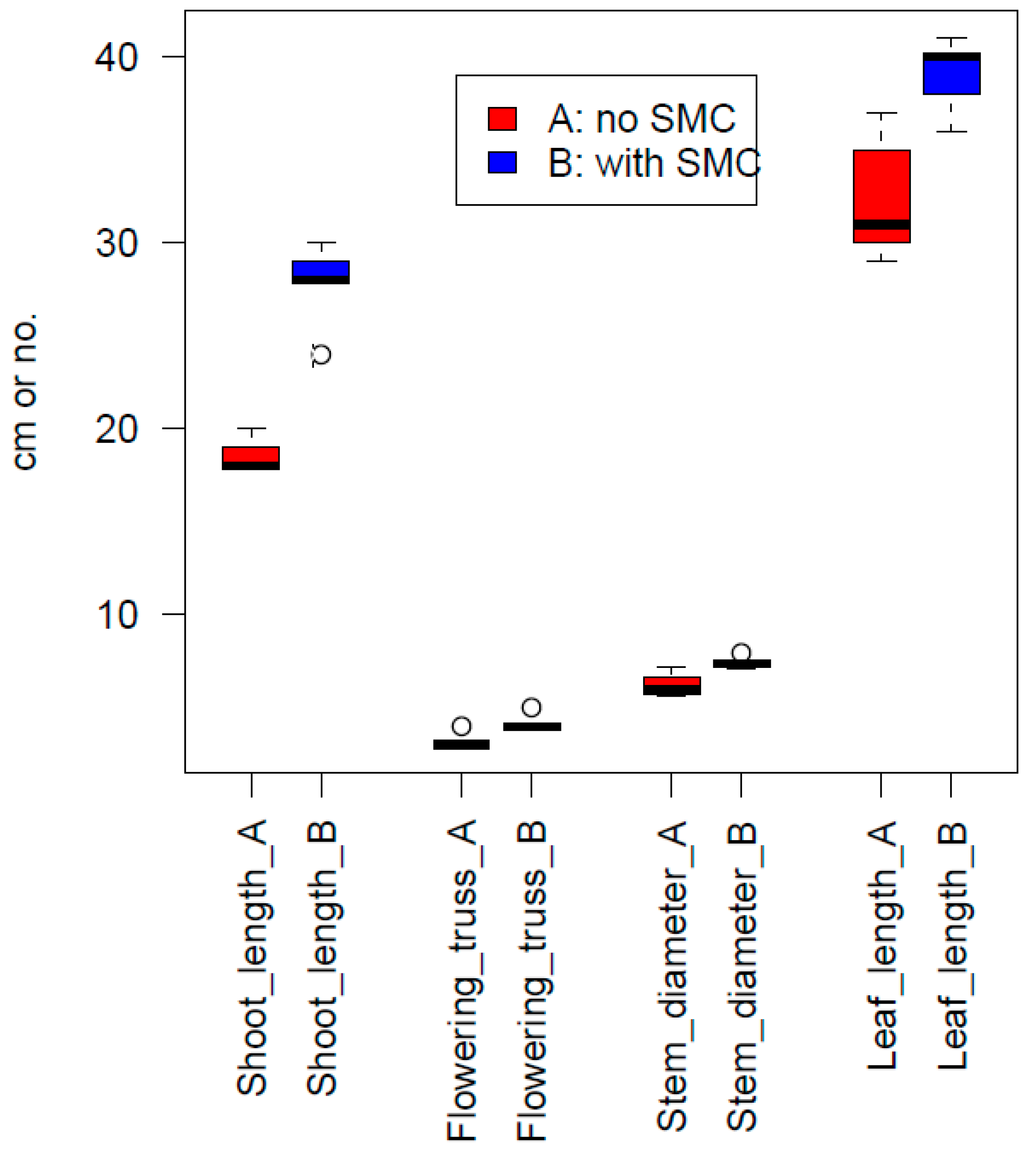
3.7. Tomato Experiment
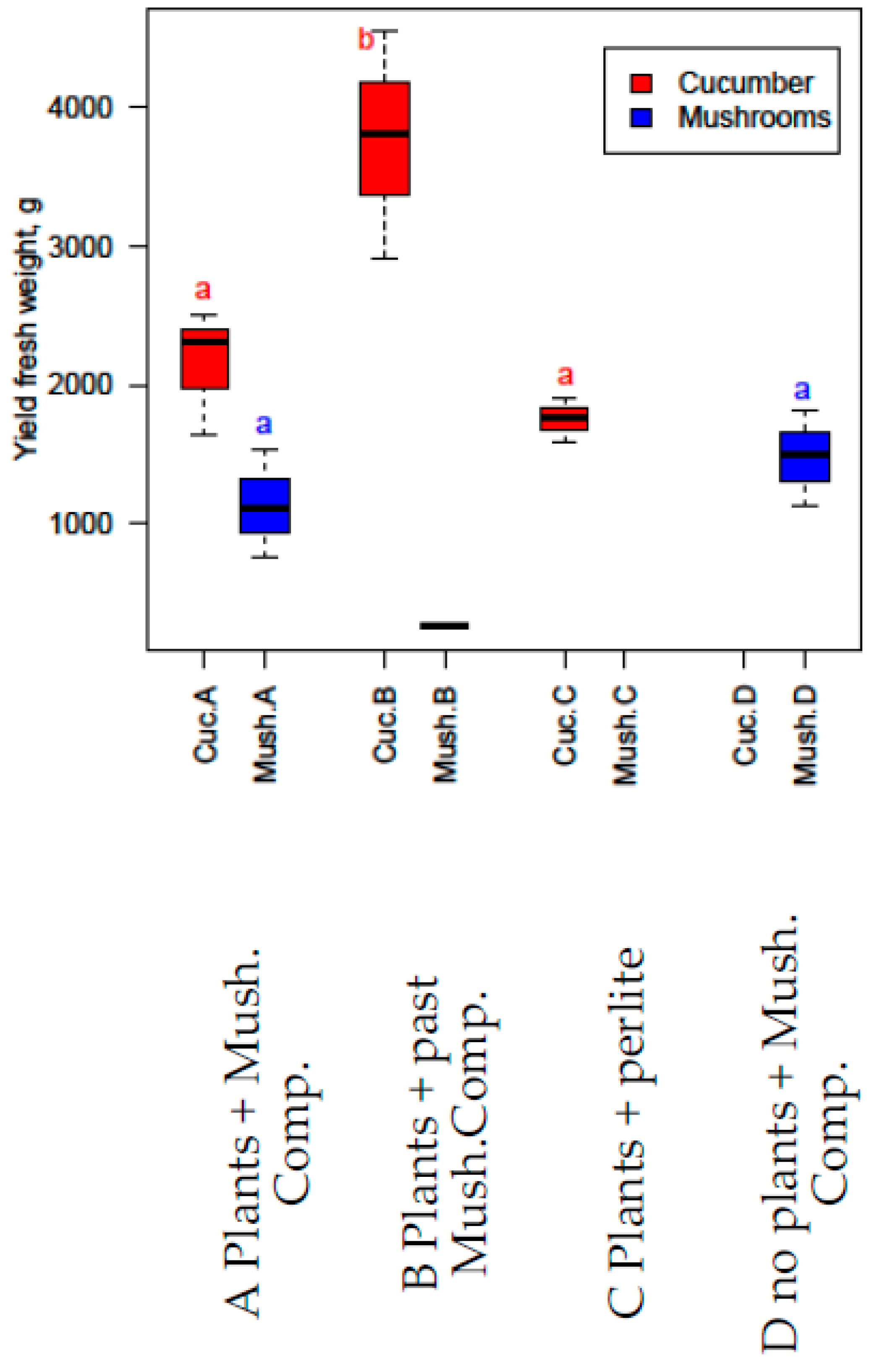
3.8. Combined Experiment
4. Discussion
5. Conclusions
- Lettuce experiment 1 (digestate fertigation used, EC = 1.5 mS cm−1).
- Lettuce experiment 3, in nutrient rich compost mix when 10% of the non-fertilised peat and perlite was substituted with SMC (no fertigation used).
- Lettuce experiment 4, in the commercial peat substrate when 20% of the volume was substituted with SMC and no fertigation was used.
- Lettuce experiment 5, in compost mix with added pasteurised SMC.
- Tomato experiment, in compost mix with added SMC (15% by volume, no fertigation used).
- Cucumber combined experiment, with pasteurised mushroom compost (no fertigation used).
- From increasing the amount of SMC further in Lettuce experiment 1.
- From adding SMC of any state to fertilised peat in Lettuce experiment 2 (mineral fertigation used, EC = 2.3 mS cm−1).
- From adding SMC of any state in Lettuce experiment 4 to the non-fertigated composts, or to any fertigated substrates.
- When pots were fertigated with digestate at EC = 2.5 mS cm−1 in Lettuce experiment 4.
- From adding spawn in Lettuce experiment 4.
- From adding SMC of any state to peat substrates or from adding non-pasteurised SMC to compost mix in Lettuce experiment 5 (no fertigation used).
Author Contributions
Funding
Conflicts of Interest
References
- Putra, P.A.; Yuliando, H. Soilless culture system to support water use efficiency and product quality: a review. Agric. Agric. Sci. Proced. 2015, 3, 283–288. [Google Scholar] [CrossRef]
- Barrett, G.E.; Alexander, P.D.; Robinson, J.S.; Bragg, N.C. Achieving environmentally sustainable growing media for soilless plant cultivation systems—A review. Sci. Hortic. 2016, 212, 220–234. [Google Scholar] [CrossRef]
- Gao, J.; Holden, J.; Kirkby, M. The impact of land-cover change on flood peaks in peatland basins. Water Resour. Res. 2016, 52, 3477–3492. [Google Scholar] [CrossRef]
- Heikkinen, K.; Ihme, R.; Osma, A.M.; Hartikainen, H. Phosphate removal by peat from peat mining drainage water during overland flow wetland treatment. J. Environ. Qual. 1995, 24, 597–602. [Google Scholar] [CrossRef]
- Fraixedas, S.; Lindén, A.; Meller, K.; Lindström, Å.; Keišs, O.; Kålås, J.A.; Husby, M.; Leivits, A.; Leivits, M.; Lehikoinen, A. Substantial decline of Northern European peatland bird populations: Consequences of drainage. Biol. Conserv. 2017, 214, 223–232. [Google Scholar] [CrossRef]
- Zheng, Y.; Huber, J.; Ping, Z.; Dixon, M. Searching for recyclable or biodegradable growing media. Acta Hortic. 2009, 819, 435–442. [Google Scholar] [CrossRef]
- Carlile, W.R. The use of composted materials in growing media. Acta Hortic. 2008, 779, 321–328. [Google Scholar] [CrossRef]
- Raviv, M. Composts in growing media: What’s new and what’s next? Acta Hortic. 2013, 982, 39–52. [Google Scholar] [CrossRef]
- Gonani, Z.; Riahi, H.; Sharifi, K. Impact of using leached spent mushroom compost as a partial growing media for horticultural plants. J. Plant Nutr. 2011, 34, 337–344. [Google Scholar] [CrossRef]
- Sendi, H.; Mohamed, M.T.M.; Anwar, M.P.; Saud, H.M. Spent mushroom waste as a media replacement for peat moss in kai-lan (Brassica oleracea var. Alboglabra) production. Sci. World J. 2013, 2013, 8. [Google Scholar] [CrossRef]
- Topcuoglu, B. The usability of sewage sludge municipal solid waste compost and spent mushroom compost as growing media on the growth of Euphorbia pulcherrima. IPCBEE 2011, 24, 386–392. [Google Scholar]
- Medina, E.; Paredes, C.; Pérez-Murcia, M.D.; Bustamante, M.A.; Moral, R. Spent mushroom substrates as component of growing media for germination and growth of horticultural plants. Bioresour. Technol. 2009, 100, 4227–4232. [Google Scholar] [CrossRef] [PubMed]
- Wever, G.; van der Burg, A.M.M.; Straatsma, G. Potential of adapted mushroom compost as a growing medium in horticulture. Acta Hortic. 2005, 697, 171–177. [Google Scholar] [CrossRef]
- Zhang, R.-H.; Duan, Z.-Q.; Li, Z.-G. Use of spent mushroom substrate as growing media for tomato and cucumber seedlings. Pedosphere 2012, 22, 333–342. [Google Scholar] [CrossRef]
- Oancea, F.; Răut, I.; Șesan, T.E.; Doni, M.; Popescu, M.; Jecu, M.L. Enhancement of biostimulant activity of spent Pleurotus substrate for seedling production. Acta Hortic. 2017, 1164, 55–62. [Google Scholar] [CrossRef]
- Chong, C.; Cline, R.A.; Rinker, D. Bark- and peat- amended spent mushroom compost for containerized culture of shrubs. HortScience 1994, 29, 781–784. [Google Scholar] [CrossRef]
- Roy, S.; Barman, S.; Chakraborty, U.; Chakraborty, B. Evaluation of Spent Mushroom Substrate as biofertilizer for growth improvement of Capsicum annuum L. J. Appl. Biol. Biotechnol. 2015, 3, 022–027. [Google Scholar] [CrossRef]
- Ribas, L.C.C.; de Mendonça, M.M.; Camelini, C.M.; Soares, C.H.L. Use of spent mushroom substrates from agaricus subrufescens (syn. a. blazei, a. brasiliensis) and lentinula edodes productions in the enrichment of a soil-based potting media for lettuce (lactuca sativa) cultivation: growth promotion and soil bioremediation. Bioresour. Technol. 2009, 100, 4750–4757. [Google Scholar] [CrossRef]
- Marques, E.L.S.; Martos, E.T.; Souza, R.J.; Silva, R.; Zied, D.C.; Dias, E.S. spent mushroom compost as a substrate for the production of lettuce seedlings. J. Agric. Sci. 2014, 6, 138–143. [Google Scholar] [CrossRef]
- Lopes, R.X.; Zied, D.C.; Martos, E.T.; de Souza, R.J.; da Silva, R.; Dias, E.S. Application of spent agaricus subrufescens compost in integrated production of seedlings and plants of tomato. International. J. Recycl. Org. Waste Agric. 2015, 4, 211–218. [Google Scholar] [CrossRef]
- Stoknes, K.; Beyer, D.M.; Norgaard, E. Anaerobically digested food waste in compost for Agaricus bisporus and Agaricus subrufescens and its effect on mushroom productivity. J. Sci. Food Agric. 2013, 93, 2188–2200. [Google Scholar] [CrossRef] [PubMed]
- Stoknes, K.; Wojciechowska, E.; Jasinska, A.; Gulliksen, A.; Tesfamichael, A.A. Growing vegetables in the circular economy; cultivation of tomatoes on green waste compost and food waste digestate. Acta Hortic. 2018, 1215, 389–396. [Google Scholar] [CrossRef]
- Liu, C.J.; Duan, Y.L.; Jin, R.Z.; Han, Y.Y.; Hao, J.H.; Fan, S.X. Spent mushroom substrates as component of growing media for lettuce seedlings. IOP Conf. Ser. Earth Environ. Sci. 2018, 185, 012–016. [Google Scholar] [CrossRef]
- Svjetlana, Z.; Parađiković, N.; Šušak, U.; Tkalec, M. Use of spent mushroom substrate for growing geranium (Pelargonium peltatum L.) and surfinia (Petunia hybrida Juss.) seedlings. In Proceedings of the Sixth International Scientific Agricultural Symposium Agrosym 2015, Jahorina, Sarajevo, Bosnia and Herzegovina, 15–18 October 2015. [Google Scholar]
- Chong, C.; Cline, R.A.; Rinker, D.; Allen, O.B. Growth and mineral nutrient status of containerized woody species in media amended with spent mushroom compost. J. Am. Soc. Hortic. Sci. 1991, 116, 242–247. [Google Scholar] [CrossRef]
- Young, J.; Holcomb, E.; Heuser, C. Greenhouse growth of marigolds in three leached sources of spent mushroom compost over a 3-year period. Hort Technol. 2002, 12, 701–705. [Google Scholar] [CrossRef]
- Jonathan, S.G.; Lawal, M.M.; Oyetunji, O.J. Effect of Spent Mushroom Compost of Pleurotus pulmonarius on Growth Performance of Four Nigerian Vegetables. Mycobiology 2011, 39, 164–169. [Google Scholar] [CrossRef]
- Wang, S.H.-L.; Lohr, V.I.; Coffey, D.L. Growth response of selected vegetable crops to spent mushroom compost application in a controlled environment. Plant Soil 1984, 82, 31–40. [Google Scholar] [CrossRef]
- Polat, E.; Uzu, H.; Topçuol, B.; Öna, K.; Onus, A.; Karac, M. Effects of spent mushroom compost on quality and productivity of cucumber (Cucumis sativus L.) grown in greenhouses. Afr. J. Biotechnol. 2009, 8, 176–180. [Google Scholar]
- Ahmad, M.; Khan, I.; Shah, B.; Naeem, A.; Khan, N.; Ullah, W.; Adnan, M.; Rizwan, S. Study on the management of Ralstonia solanacearum (Smith) with spent mushroom compost. J. Entomolog. Zool. Stud. 2016, 4, 114–121. [Google Scholar]
- Tesfamichael, A.A.; Stoknes, K. Substitution of peat with vermicompost from food waste digestate and green waste compost. Acta Hortic. 2017, 1168, 399–406. [Google Scholar] [CrossRef]
- Stoknes, K.; Scholwin, F.; Jasinska, A.; Wojciechowska, E.; Mleczek, M.; Hanc, A.; Niedzielski, P. Cadmium mobility in a circular food-to-waste-to-food system and the use of a cultivated mushroom (Agaricus subrufescens) as a remediation agent. J. Environ. Manag. 2019, 245, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Visscher, H. Casing soil. In The Cultivation of Mushrooms; van Griensven, L.J.L.D., Ed.; Darlington Mushroom Laboratories: Sussex, UK, 1988; pp. 73–89. [Google Scholar]
- Caesar-Tonthat, T.C.; Espeland, E.; Caesar, A.J.; Sainju, U.M.; Lartey, R.T.; Gaskin, J.F. Effects of agaricus lilaceps fairy rings on soil aggregation and microbial community structure in relation to growth stimulation of western wheatgrass (Pascopyrum smithii) in Eastern Montana rangeland. Microb. Ecol. 2013, 66, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Edwards, P.J. Effects of the fairy ring fungus agaricus arvensis on nutrient availability in grassland. New Phytol. 1988, 110, 377–381. [Google Scholar] [CrossRef]
- Wolfe, B.E.; Kuo, M.; Pringle, A. Amanita thiersii is a saprotrophic fungus expanding its range in the United States. Mycologia 2012, 104, 22–33. [Google Scholar] [CrossRef] [Green Version]
- Guhr, A.; Borken, W.; Spohn, M.; Matzner, E. Redistribution of soil water by a saprotrophic fungus enhances carbon mineralization. Proc. Natl. Acad. Sci. USA 2015, 112, 14647–14651. [Google Scholar] [CrossRef] [Green Version]
- Gramss, G. The universe of basidiomycetous ground fungi. In Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology; Formatex Research Center: Badajoz, Spain, 2010; pp. 218–229. [Google Scholar]
- Hildén, K.; Mäkelä, M.R.; Lankinen, P.; Lundell, T. Agaricus bisporus and related Agaricus species on lignocellulose: Production of manganese peroxidase and multicopper oxidases. Fungal Genet. Biol. 2013, 55, 32–41. [Google Scholar] [CrossRef]
- Fritsche, G. Tests on breeding with Agaricus arvensis. Mushroom Sci. 1979, 10, 91–101. [Google Scholar]
- Noble, R. Alternative Agaricus: The Culture of Agaricus W4 and Agaricus Arvensis; HDC Project, M4b; Horticulture Research International: Wellesbourne, Warwick, UK, 1995; 14p. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Treatments | Number of Replicates | Composition by Volume (%) | Fertigation | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Green Waste Compost | Vermi-compost | Commercial Fertilised Peat | Peat | Perlite | A.sub. SMC | Pasteur. A.sub. SMC | A.bis. SMC | ||||
| Lettuce 1 | A Compost mix | 10 | 91 | 9 | Digestate EC = 1.5 mS cm−1 | ||||||
| B Comp + 4% SMC | 87 | 9 | 4 | ||||||||
| C Comp + 8% SMC | 84 | 8 | 8 | ||||||||
| D Comp + 15% SMC | 77 | 8 | 15 | ||||||||
| E Comp + 25% SMC | 63 | 12 | 25 | ||||||||
| Lettuce 2 | A Peat | 22 | 100 | Mineral EC = 2.3 mS cm−1 | |||||||
| B Peat + AsubSMC | 90 | 10 | |||||||||
| C Peat + past AsubSMC | 90 | 10 | |||||||||
| D Peat + AbitSMC | 90 | 10 I | |||||||||
| E Peat + min AsubSMC | 98 | 2 II | |||||||||
| Lettuce 3 | A Peat | 6 | 100 | no | |||||||
| B Peat + SMC | 90 | 10 | |||||||||
| C Comp low + SMC | 50 | 20 | 20 | 10 | |||||||
| D Comp low + past SMC | 50 | 20 | 20 | 10 | |||||||
| E Comp high + SMC | 30 | 40 | 15 | 5 | 10 | ||||||
| F Comp high | 30 | 40 | 20 | 10 | |||||||
| Lettuce 4 | A Peat | 8 | 100 | Digestate EC = 2.5 mS cm−1 on half of the replicate pots | |||||||
| B Peat + Asub SMC | 80 | 20 | |||||||||
| C Compost mix | 60 | 40 | |||||||||
| D Comp + Asub SMC | 40 | 40 | 20 | ||||||||
| E Comp + past Asub SMC | 40 | 40 | 20 | ||||||||
| F Comp + Asub spawn | 58 | 40 | 2 III | ||||||||
| G Comp +pastAsub spawn | 58 | 40 | 2 III | ||||||||
| H Comp + Abis spawn | 58 | 40 | 2 III | ||||||||
| Lettuce 5 | A Peat | 20 | 100 | no | |||||||
| B Peat + Asub SMC | 80 | 20 | |||||||||
| C Peat + Abis SMC | 80 | 20 | |||||||||
| D Peat + past Abis SMC | 80 | 20 IV | |||||||||
| E Compost mix | 60 | 40 | |||||||||
| F Comp + Asub SMC | 48 | 32 | 20 | ||||||||
| G Comp + Abis SMC | 48 | 32 | 20 | ||||||||
| H Comp + past Asub SMC | 48 | 32 | 20 | ||||||||
| Tomato | A Compost mix | 5 | 60 | 40 | no | ||||||
| B Comp + Asub SMC | 51 | 34 | 15 | ||||||||
| CombinedV (cucumber and mushroom) | A Plants + Mush. Comp. | 3 | 50 | 20 | 30VI | no | |||||
| B Plants + past M.Comp. | 50 | 20 | 30VI | ||||||||
| C Plants + perlite | 50 | 20 | 30 | ||||||||
| D no Plants + M. Comp. | 50 | 20 | 30VI | ||||||||
| A. sub. SMC Exp 2 | A. sub. SMC Exp 5 | A. bisp. SMC Exp 5 | A. sub. | A. bisp. | Vermi-Compost | Green Waste Compost | Commercial Peat Substrate | |
|---|---|---|---|---|---|---|---|---|
| Mushroom Compost in Cultivation | ||||||||
| pH | 6.1 | 5.1 | 8.7 | 5.7 | 6.6 | 7.7 | 7 | 5.8 |
| EC (mS cm−1) | 3.8 | 2.6 | 23 | 3.35 | 4.1 | 9 | 2 | 1.3 |
| Tot-N, mg kg−1 FW | 4530 | 5330 | 6000 | 2000 | 3000 | |||
| NO3-N, mg L−1 | 84 | 5 | 290 | 650 | 30 | 74 | ||
| NH4-N, mg L−1 | 5 | 3 | 1 | 5 | 50 | |||
| P, mg L−1 | 75 | 110 | 100 | 150 | 120 | 40 | ||
| K, mg L−1 | 377 | 4900 | 5500 | 1000 | 600 | 182 | ||
| Mg, mg L−1 | 340 | 360 | 330 | 350 | 125 | 125 | ||
| S, mg L−1 | 1640 | 1200 | 1600 | 200 | 50 | 52 | ||
| Ca, mg L−1 | 2000 | 2500 | 2000 | 900 | 971 | |||
| Mn, mg L−1 | 6.6 | 2.7 | 2.5 | 2 | 2.4 | |||
| B, mg L−1 | 0.88 | 0.69 | 1.6 | 1.5 | 0.2 | |||
| Fe, mg L−1 | 29 | 6.6 | 0.4 | 1 | 1.5 | |||
| Na, mg L−1 | 1200 | 1300 | 600 | 40 | 32 | |||
| Cl, mg L−1 | 2400 | 2400 | 700 | 30 | 11 | |||
| Dry Matter | 35% | 25% | 30% | 45% | 42% | |||
| Ash content | 51% | 64% | 62% | 40% | 60% | |||
| A | B | C | D | E | F | G | H | |
|---|---|---|---|---|---|---|---|---|
| At sowing: | ||||||||
| pH | 7.0 | 6.8 | 8.2 | 8.2 | 8.0 | 8.5 | 8.4 | 8.2 |
| EC, mS cm−1 | 2.4 | 9.1 | 4.3 | 12.5 | 13 | 4.3 | 5.4 | 5.5 |
| Tot-N, mg kg−1 FW | 2800 | 5520 | 7620 | 7630 | 8580 | 6680 | 7460 | 6950 |
| min. N i, mg L−1 | 135 | 205 | 119 | 310 | 260 | 43,5 | 59 | 93 |
| P, mg L−1 | 70 | 80 | 175 | 155 | 160 | 165 | 145 | 150 |
| K, mg L−1 | 250 | 955 | 730 | 1800 | 1550 | 785 | 850 | 795 |
| S, mg L−1 | 77 | 650 | 120 | 730 | 610 | 150 | 330 | 280 |
| Fe, mg L−1 | 1.1 | 1.7 | 0.53 | 1.4 | 1.1 | 0.62 | 0.55 | 0.56 |
| Na, mg L−1 | 38 | 230 | 270 | 570 | 540 | 290 | 320 | 300 |
| Cl, mg L−1 | 10 | 410 | 250 | 900 | 840 | 280 | 310 | 250 |
| At end of experiment: | ||||||||
| min. N i, mg L−1 (−/+ fert) | 2/52 | 7/61 | 10/30 | 16/89 | 50/97 | 16/44 | 2/35 | 5/55 |
| P, mg L−1 (−/+ fert) | 31/35 | 78/81 | 160/150 | 150/170 | 160/180 | 150/170 | 160/150 | 160/170 |
| K, mg L−1 (−/+ fert) | 24/43 | 39/50 | 310/75 | 390/330 | 710/300 | 340/110 | 190/68 | 120/66 |
| A | B | C | D | E | F | G | H | |
|---|---|---|---|---|---|---|---|---|
| Germination% | 85 | 60 | 30 | 90 | 70 | 60 | 60 | 75 |
| pH | 5.4 | 6 | 5.8 | 6 | 7.8 | 8.5 | 7.9 | 8 |
| EC, mS cm−1 | 3.2 | 6.7 | 6.8 | 5.2 | 3.9 | 6.5 | 9.2 | 11 |
| Tot-N, mg kg−1 FW | 3210 | 3910 | 4360 | 2720 | 6590 | 5440 | 6810 | 5950 |
| min. N i, mg L−1 | 250 | 160 | 230 | 180 | 55 | 35 | 150 | 140 |
| P, mg L−1 | 110 | 71 | 72 | 79 | 140 | 120 | 150 | 140 |
| K, mg L−1 | 300 | 940 | 980 | 820 | 950 | 1800 | 1800 | 2100 |
| S, mg L−1 | 140 | 320 | 320 | 320 | 180 | 430 | 420 | 520 |
| Na, mg L−1 | 40 | 210 | 220 | 180 | 170 | 370 | 370 | 450 |
| Cl, mg L−1 | 2 | 480 | 470 | 370 | 180 | 770 | 770 | 1000 |
| A | B | |
|---|---|---|
| Substrate (Plant Available Nutrients) | ||
| pH | 8.4 | 8.4 |
| EC, mS cm−1 | 2.2 | 5.3 |
| NO3-N + NH4-N, mg L−1 | 19 | 40 |
| P, mg L−1 | 130 | 86 |
| K, mg L−1 | 700 | 1000 |
| S, mg L−1 | 71 | 290 |
| Fe, mg L−1 | 1.0 | 5.4 |
| Plants (leaf sap analysis) | ||
| Tot-N, mg−kg DM | 17800 | 23900 |
| P, mg−kg DM | 4230 | 3330 |
| K, mg−kg DM | 33000 | 36000 |
| S, mg−kg DM | 10000 | 12000 |
| Fe, mg−kg DM | 55 | 64 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoknes, K.; Wojciechowska, E.; Jasinska, A.; Noble, R. Amelioration of Composts for Greenhouse Vegetable Plants Using Pasteurised Agaricus Mushroom Substrate. Sustainability 2019, 11, 6779. https://doi.org/10.3390/su11236779
Stoknes K, Wojciechowska E, Jasinska A, Noble R. Amelioration of Composts for Greenhouse Vegetable Plants Using Pasteurised Agaricus Mushroom Substrate. Sustainability. 2019; 11(23):6779. https://doi.org/10.3390/su11236779
Chicago/Turabian StyleStoknes, Ketil, Ewelina Wojciechowska, Agnieszka Jasinska, and Ralph Noble. 2019. "Amelioration of Composts for Greenhouse Vegetable Plants Using Pasteurised Agaricus Mushroom Substrate" Sustainability 11, no. 23: 6779. https://doi.org/10.3390/su11236779
APA StyleStoknes, K., Wojciechowska, E., Jasinska, A., & Noble, R. (2019). Amelioration of Composts for Greenhouse Vegetable Plants Using Pasteurised Agaricus Mushroom Substrate. Sustainability, 11(23), 6779. https://doi.org/10.3390/su11236779